 2021, Vol. 42
2021, Vol. 42



2. 苏州科技大学天平学院, 苏州 215009;
3. 城市生活污水资源化利用技术国家地方联合工程实验室, 苏州 215009;
4. 江苏高校水处理技术与材料协同创新中心, 苏州 215009
2. College of Tianping, Suzhou University of Science and Technology, Suzhou 215009, China;
3. National and Local Joint Engineering Laboratory of Municipal Sewage Resource Utilization Technology, Suzhou 215009, China;
4. Jiangsu High Education Collaborative Innovation Center of Water Treatment Technology and Material, Suzhou 215009, China
部分亚硝化-厌氧氨氧化(partial nitritation/ANAMMOX, PN/A)是一种全自养生物脱氮工艺, 其首先通过好氧氨氧化菌(AOB)将一部分氨氮氧化为亚硝态氮, 再由厌氧氨氧化菌(AMX)将剩余的氨氮和亚硝态氮转化为氮气, 并产生少量硝态氮, 理论脱氮率在85%以上[1].一般认为, 生物脱氮反应对环境温度较为敏感.在中高温条件(30~35℃)下, AOB和AMX的比生长速率可以分别达到0.81~0.85 d-1和0.002 7 d-1, 污泥脱氮活性处于最佳状态[2].当环境温度低于15℃时, PN/A污泥的增殖速率和反应活性均会大幅降低, 并容易出现亚硝盐氧化菌(NOB)过度生长, 导致厌氧氨氧化反应路径受阻等现象, 这在很大程度上限制了PN/A工艺在大规模市政污水处理中的推广应用[3, 4].
有研究表明, 经过长期驯化后, 以颗粒污泥为核心的单级PN/A反应器能够在低温环境中实现稳定的脱氮过程.Gonzalez-Martinez等[5]在260 d内将PN/A系统的反应温度由35℃逐级降低至15℃, 并使总氮去除负荷稳定在0.28~0.32 kg·(m3·d)-1.Lotti等[6]的研究表明, 经低温驯化后, PN/A颗粒污泥反应器在15℃下的处理效能仍远高于传统活性污泥法, 对总氮去除负荷可达到0.4 kg·(m3·d)-1的水平.Gilbert等[7]的研究发现, 颗粒污泥在低温适应性方面较悬浮污泥有明显优势, 前者能够在13℃下保持较高的脱氮效率, 出水中没有出现亚硝态氮的大量累积.以往的报道大多关注PN/A颗粒污泥在低温驯化过程中的性能演变, 但对污泥驯化后反应热力学特征、颗粒外观形态和微生物多样性的变化仍缺乏系统阐述.
为此, 本文将中高温培养和低温驯化后的PN/A颗粒污泥作为研究对象, 通过对比分析, 深入探究两者在环境温度敏感性、颗粒密实程度和微生物群落结构等方面的差异, 并对导致差异的可能原因进行论述, 以期为全自养生物脱氮技术在低温条件下的运行优化提供理论指导.
1 材料与方法 1.1 PN/A颗粒污泥在本研究中, 中高温培养的PN/A颗粒污泥(GH)取自1个实验室规模的连续流反应器, 曝气区有效容积为1.7 L.进水采用无机人工配水, 以氨氮(浓度约70 mg·L-1)作为唯一氮源.运行初期, 反应器内水温控制在(30±1)℃, 氨氮容积负荷为2.2 kg·(m3·d)-1.稳定运行100 d后, 对GH进行取样, 颗粒呈现深红棕色, 总氮比去除负荷为0.25 g·(g·d)-1[8].随后, 逐级降低反应器水温至(15±1)℃, 并采取限制曝气和延长水力停留时间(HRT)等控制方法, 实现了反应器在低温条件下的稳定运行[9].低温驯化200 d后, 颗粒污泥(GL)呈棕黄色, 粒径明显减小, 总氮比去除负荷降至0.11 g·(g·d)-1.
1.2 污泥活性测定和活化能计算在批次反应中, 控制水温分别为10、15、20、25、30和35℃.分别取10 mL颗粒污泥混合液, 装入多个配备有透气硅胶塞的150 mL锥形瓶中, 加入90 mL反应器进水(pH 7.6~7.8), 并置于机械摇床中振荡反应.控制转速为160 r·min-1, 确保溶液处于好氧状态.每隔10或20 min取样测定上清液中氨氮(NH4+-N)、硝态氮(NO3--N)和亚硝态氮(NO2--N)浓度, 并计算总无机氮(TIN)的比降解速率[q(TIN), mg·(g·h)-1], 见式(1):
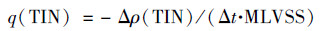
|
(1) |
式中, Δρ(TIN)为反应前后总无机氮的消耗量, mg·L-1; Δt为浓度线性变化对应的反应时间, h; MLVSS为混合液中挥发性悬浮固体浓度, mg·L-1.
依据阿伦尼乌斯(Arrhenius)方程, 对两种PN/A颗粒污泥在不同温度区间内的q(TIN)进行线性拟合, 计算相应的反应活化能Ea[8]:
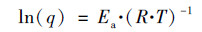
|
(2) |
为表征颗粒污泥对环境温度的敏感性, 使用下式计算PN/A反应的温度系数θ[10]:
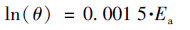
|
(3) |
式中, Ea为反应活化能, J·mol-1; R为气体常数, 8.314 J·(K·mol)-1; T为热力学温度, K.
1.3 分析方法水中NH4+-N、NO3--N和NO2--N浓度分别采用纳氏试剂光度法、紫外分光光度法和N-(1-萘基)-乙二胺光度法测定, 污泥MLVSS采用标准重量法测定[11].采用便携式pH计(METTLER TOLEDO)和溶解氧仪(HACH HQ30d型)测定溶液pH和溶解氧(DO)浓度.
使用OLYMPUS CX41型显微镜观察污泥形态变化.采用湿网筛分法测算污泥粒径分布, 具体为从反应器中取出一定量污泥样品, 经生理盐水反复清洗后, 依次经孔径2.0、1.6、1.4、1.0、0.8、0.6、0.5、0.3和0.2 mm筛网筛分, 并计算各粒径区间样品占总质量的质量分数.
通过静态沉降实验, 测定颗粒污泥沉降性能.随机选取20个颗粒污泥, 在1 L量筒(高45 cm)内测定污泥下降30 cm的沉降时间, 并计算平均沉降速率v(m·h-1).
采用甲醇-NaOH法提取颗粒污泥的胞外聚合物(EPS)[12], 其主要组分中蛋白质(PN)和多糖(PS)含量分别采用Lowry法和改进苯酚-硫酸法测定[13].
1.4 高通量测序对GH和GL污泥, 分别采集6个平行样品, 置于-80℃条件下保存.使用试剂盒(Fast DNA Spin kit for soil, M.P., U.S.)对所有污泥样品同时进行DNA的提取.选择引物338F (5′-ACTCCTACGGG AGGCAGCA-3′)和806R (5′-GGACTACHVGGGTWT CTAAT-3′)进行16S rRNAV3-V4区基因扩增.在完成PCR扩增后, 使用AxyPrepTM DNA凝胶提取试剂盒(Axygen Biosciences, Union City, U.S.)对产物进行纯化和定量.12个样品的高通量测序在Illumina MiSeq测序平台上完成.基于测序结果, 在OTU水平上, 对样品进行α多样性分析、主坐标分析(PCoA)和ANOSIM分析(bray_curtis算法).在门和纲水平上, 对两种污泥菌群结构进行Wilcox秩和检验, 并绘制属水平的系统发育树, 方法详见文献[14].
2 结果与讨论 2.1 污泥活性对温度的敏感性根据污泥在不同温度下的脱氮活性可知, GH的最大q(TIN)值出现在35℃条件下, 其脱氮活性随着水温的下降而显著降低, 10℃对应的q(TIN)值仅相当于35℃的4%左右, 并出现了明显的亚硝态氮累积现象.相比之下, 低温驯化使GL的最适脱氮温度降至25℃, 对应q(TIN)值分别比35℃和10℃的脱氮速率高出42.8%和97.0%.Hu等[15]和Park等[16]的研究也发现, AMX污泥经长期低温驯化(< 15℃)后, 最适反应温度会由30~35℃降至25℃左右.
如图 1所示, 在低温(10~20℃)和中高温(20~35℃)区间内, 两种PN/A颗粒的ln(q)与1/T呈现线性相关.依据Arrhenius方程, 计算出不同温度区间内自养脱氮反应的表观活化能Ea, 并与已有数据进行比较, 结果如表 1所示.在本研究中, GH在20~35℃区间的Ea值约为10~20℃对应值的6.2倍, 明显高于已有报道的3.9倍均值[4, 15~17].这表明低温环境会显著抑制未经驯化污泥的自养脱氮活性.相比之下, GL在10~20℃区间的Ea值较GH降低了约28.4%, 20~35℃区间的Ea值则升高了1.1倍.尽管GL在低温区间的脱氮效能仍然较差, 但低温驯化明显降低了PN/A反应对温度变化的敏感性.

|
GH数据来自本研究, GH-1~GH-3数据来自文献[4, 17, 15]; GL数据来自本研究, GL-1~GL-3数据来自文献[15, 16, 17] 图 1 使用Arrhenius方程对中高温培养和低温驯化污泥的脱氮速率进行拟合 Fig. 1 Arrhenius plot for nitrogen removal rates of sludges cultivated at medium-high temperatures and acclimated to low temperatures |
|
|
表 1 不同温度区间内污泥自养脱氮反应的表观活化能 Table 1 Apparent activation energy of autotrophic nitrogen removal by sludges in different temperature intervals |
从工程应用的角度, 温度系数θ更适合用于表征污泥性能随反应温度的变化特征.有研究表明, 传统活性污泥和厌氧氨氧化污泥的θ值通常随温度的降低而增大[4, 18, 19].在本研究中, GH和GL在中高温区间的θ值分别为1.046和1.097, 低温区间的数值则相应升至1.317和1.218, 驯化过程使得污泥θ值的相对增幅明显减小.上述变化可以作为评判PN/A污泥是否成功适应低温环境的重要依据.
2.2 颗粒污泥形态的差异如表 2所示, 低温驯化使得PN/A颗粒污泥的粒径明显减小, 其中, 主导粒径区间由0.9~1.0 mm(占比61.3%)减小至0.2~0.5 mm(占比45.2%), 平均粒径由1.20 mm减小至0.89 mm.同时, 污泥EPS含量增加, PN/PS比值上升, 污泥结构更加松散, 边缘变得不规则(图 2).由沉降实验结果可知, GL的平均沉降速率v较驯化前(GH)降低了53.4%, 颗粒的密实程度变差.He等[20]和Guo等[21]在研究低温驯化(2.5~13℃)对厌氧氨氧化颗粒污泥性能影响时也有类似结论.
|
|
表 2 低温驯化前后PN/A颗粒污泥的形态结构特性 Table 2 Morphological characteristics of PN/A granular sludges before and after cold acclimation |

|
图 2 低温驯化前后, PN/A颗粒污泥的外观形态对比 Fig. 2 Morphological comparison of PN/A granular sludges before and after cold acclimation |
实际上, 低温环境不仅导致了自养菌生长速率和反应活性的下降, 还会使流体的黏度增大, 水力剪切作用增强, 大颗粒很难稳定存在[22].通常认为, 产生大量EPS有利于削弱低温环境对功能菌活性的胁迫作用, 是微生物聚集体自适应策略的重要体现[23, 24].作为亲水性较强的EPS组分, PN含量的持续增加将使得更多的结合水进入到微生物聚集体中, 形成密度较低的多孔絮状结构, 削弱污泥的沉降性能[25, 26].
2.3 污泥菌群结构的差异使用MiSeq高通量测序技术对低温驯化前后PN/A颗粒污泥的菌群结构进行分析.如表 3所示, 6个GL样品中的运算分类单位(OTU)数量较GH组多出了255个, 另有384个OTU在两组中均能找到.有研究认为, 低温环境容易导致污泥中菌群多样性和丰度的降低[27, 28], 但本研究中GL组的多样性指数(高Shannon指数、低Simpson指数)和菌群丰度(Ace、Chao指数更大)均明显高于GH组.这说明改变菌群结构也是PN/A颗粒污泥适应低温条件的重要策略之一.从PCoA分析的结果上看[图 3(a)], GH组和GL组在主坐标PCoA1(贡献率57.71%)方向上呈现区域聚集性分布, 同时, OTU水平上的ANOSIM分析进一步说明了组间的显著差异[图 3(b)].
|
|
表 3 低温驯化前后, PN/A颗粒污泥的微生物多样性指数 Table 3 The α diversity indices of PN/A granular sludges before and after cold acclimation |

|
图 3 两组污泥样品间的菌群结构差异性分析(OTU水平) Fig. 3 Difference analysis of the bacterial community structure in two sample groups(OTU level) |
图 4分别给出了两组污泥样品在门和纲水平上相对丰度前15位的细菌分布情况.其中, GH组的优势菌门依次为Proteobacteria(34.8%, 丰度)、Planctomycetes(22.2%, 丰度)、Chloroflexi(16.8%, 丰度)、Bacteroidetes(17.8%, 丰度)和Acidobacteria(4.7%, 丰度).在GL组中, Planctomycetes、Chloroflexi和Acidobacteria菌门的相对丰度有所增长(1.6%~7.7%), Proteobacteria和Bacteroidetes菌门的占比则分别减少了10.2%和6.1%.在纲水平上, 低温驯化后PN/A颗粒污泥中AOB所属的γ-Proteobacteria丰度较驯化前下降了12.8%, AMX所属的Brocadiae和异养菌Anaerolineae丰度分别增长了3.0%和4.2%, NOB占比始终低于1%.

|
图 4 两组污泥样品中丰度前15的细菌组成差异分析 Fig. 4 Statistical analyses of top 15 taxa in two sample groups |
图 5为PN/A颗粒污泥在属水平上的系统发育树.尽管Candidatus_Brocadia属AMX被认为具有很强的低温适应能力, 其在运行温度10~18℃的A/O颗粒污泥反应器[29]、气提式反应器[30]和上流式厌氧污泥床(UAnSB)[31, 32]中都有检出, 但本研究中Candidatus_Kuenenia始终是占绝对主导的AMX菌属.根据Dosta等[33]的测算, Candidatus_Kuenenia脱氮反应的活化能Ea约为63 kJ·mol-1, 这与GL在高温区间的Ea值61.62 kJ·mol-1(表 1)非常接近.另外, GL组中AOB(Nitrosomonas)与AMX(Candidatus_Kuenenia)的丰度比值为0.03, 明显小于GH组的0.34.在低温环境中, 占据颗粒表面的AOB生长速率放缓, 经水力剪切和碰撞摩擦后, AOB菌簇容易从污泥上脱落并随出水淘洗, 造成相对丰度下降.相比之下, 位于颗粒内层的AMX获得了较好的保护, 有利于发生缓慢富集过程.有研究表明, 在13~16℃条件下, Candidatus_Kuenenia在厌氧氨氧化颗粒污泥中仍可持续增殖, 相对丰度可达42.4%~55.2%, 使系统脱氮效率稳定在60%~80%[20, 23].此外, GH和GL中反硝化菌Denitratisoma的占比相差不大, 但低温驯化使Haliangium和norank_Caldilineaceae出现了明显的富集现象.有研究认为, 后两种菌属可利用AMX细胞残体作为生长基质, 发生部分反硝化反应, 参与生物脱氮过程[34, 35].

|
图 5 PN/A颗粒污泥的微生物系统发育树(属水平) Fig. 5 Microbial phylogenetic tree (genus level) of PN/A granules |
与Gilbert等[7]的报道相似, 本研究中低温驯化并未改变PN/A污泥中的AMX菌属类型, 同时, AMX丰度的增长并不能直接解释脱氮反应自由能Ea的显著变化.为此, Rattray等[35]的研究认为, 低温驯化改变了AMX中重要细胞器——厌氧氨氧化体(ANAMMOXOSOME)的膜结构.当水温由20℃降至12℃时, AMX可通过调高C20-梯烷脂肪酸产量, 抑制C18-梯烷脂肪酸形成, 改变生物膜的双层结构, 以适应温度变化.Lin等[36]使用代谢蛋白组学技术, 从蛋白质表达的层面, 对不同AMX菌属在低温环境中的自适应策略进行了表征.结果表明, 当温度由35℃降至20℃时, Candidatus_Kuenenia细胞中与脱氮相关的蛋白酶表达(NirS、HZS和HDH)没有发生明显改变, 但继续降至15℃时, 类脱氢酶(kustc0824和kustc0827)丰度降低, 并导致电子流失衡, 氨氧化和亚硝酸盐还原速率均受到抑制, 负责能量供给的功能蛋白数量也相应减少.在经过低温驯化后, 上述电子流失衡现象有可能得到缓解, 污泥通过优化AMX菌群的功能性结构, 可以有效提高系统的脱氮效能.类似地, 在低温环境中, Candidatus_Jettenia属AMX可以通过调节自身基因转录水平, 由翻译启动因子(IF-3)形成大量负责能量供给的功能蛋白, 以驱动自养脱氮反应.目前, 关于低温驯化对自养脱氮反应调控机制的影响仍有待进一步研究.
3 结论(1) 经低温驯化后, PN/A颗粒污泥进行脱氮反应的最适温度由原先的35℃下降至25℃.在10~20℃条件下, GL的反应自由能Ea较GH降低了28.4%, 前者的θ值也更小.这意味着低温驯化显著降低了PN/A颗粒污泥对环境温度的敏感性.
(2) 低温驯化使得GL的平均粒径较GH减小了25.8%, EPS含量的增长有利于维持颗粒污泥结构的稳定性, 但也使颗粒的沉降性能显著降低, 平均沉降速率v减小了53.4%.
(3) 低温驯化并未改变PN/A颗粒污泥中的AOB和AMX菌属类型, 但GL中菌群多样性有所提高.Candidatus_Kuenenia属AMX在颗粒污泥中的有效富集是实现菌群功能结构优化, 适应低温环境的重要基础.
| [1] | Third K A, Paxman J, Schmid M, et al. Treatment of nitrogen-rich wastewater using partial nitrification and Anammox in the CANON process[J]. Water Science & Technology, 2005, 52(4): 47-54. |
| [2] | Strous M, Heijnen J J, Kuenen J G, et al. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms[J]. Applied Microbiology and Biotechnology, 1998, 50(5): 589-596. DOI:10.1007/s002530051340 |
| [3] | Lotti T, Kleerebezem R, van Erp Taalman Kip C, et al. Anammox growth on pretreated municipal wastewater[J]. Environmental Science & Technology, 2014, 48(14): 7874-7880. |
| [4] | Lotti T, Kleerebezem R, Van Loosdrecht M C M. Effect of temperature change on anammox activity[J]. Biotechnology and Bioengineering, 2015, 112(1): 98-103. DOI:10.1002/bit.25333 |
| [5] | Gonzalez-Martinez A, Rodriguez-Sanchez A, Garcia-Ruiz M J, et al. Performance and bacterial community dynamics of a CANON bioreactor acclimated from high to low operational temperatures[J]. Chemical Engineering Journal, 2016, 287: 557-567. DOI:10.1016/j.cej.2015.11.081 |
| [6] | Lotti T, Kleerebezem R, Hu Z, et al. Simultaneous partial nitritation and anammox at low temperature with granular sludge[J]. Water Research, 2014, 66: 111-121. DOI:10.1016/j.watres.2014.07.047 |
| [7] | Gilbert E M, Agrawal S, Schwartz T, et al. Comparing different reactor configurations for Partial Nitritation/Anammox at low temperatures[J]. Water Research, 2015, 81: 92-100. DOI:10.1016/j.watres.2015.05.022 |
| [8] |
陈希, 钱飞跃, 王建芳, 等. 环境因子对全自养脱氮颗粒污泥功能菌协同效应的影响[J]. 环境科学, 2018, 39(4): 1756-1762. Chen X, Qian F Y, Wang J F, et al. Effects of environmental factors on the synergy of functional bacteria in completely autotrophic granular sludge[J]. Environmental Science, 2018, 39(4): 1756-1762. |
| [9] |
刘雨馨, 王建芳, 钱飞跃, 等. 低温下全自养脱氮颗粒污泥适应低基质效能[J]. 环境科学, 2020, 41(9): 4161-4168. Liu Y X, Wang J F, Qian F Y, et al. Adaptability of completely autotrophic nitrogen removal over granular sludge to low-strength at low temperature[J]. Environmental Science, 2020, 41(9): 4161-4168. |
| [10] | Grady C P L Jr, Lim H C. Biological wastewater treatment: theory and applications[M]. New York: Marcel Dekker, Inc, 1980. |
| [11] | 国家环境保护局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [12] |
李金璞, 张雯雯, 杨新萍. 活性污泥污水处理系统中胞外多聚物的作用及提取方法[J]. 生态学杂志, 2018, 37(9): 2825-2833. Li J P, Zhang W W, Yang X P. The roles and extraction methods of extracellular polymeric substances in activated sludge wastewater treatment system[J]. Chinese Journal of Ecology, 2018, 37(9): 2825-2833. |
| [13] | Adav S S, Lee D J. Extraction of extracellular polymeric substances from aerobic granule with compact interior structure[J]. Journal of Hazardous Materials, 2008, 154(1-3): 1120-1126. DOI:10.1016/j.jhazmat.2007.11.058 |
| [14] | Wang J F, Qian F Y, Liu X P, et al. Cultivation and characteristics of partial nitrification granular sludge in a sequencing batch reactor inoculated with heterotrophic granules[J]. Applied Microbiology and Biotechnology, 2016, 100(21): 9381-9391. DOI:10.1007/s00253-016-7797-9 |
| [15] | Hu Z Y, Lotti T, De Kreuk M, et al. Nitrogen removal by a nitritation-anammox bioreactor at low temperature[J]. Applied and Environmental Microbiology, 2013, 79(8): 2807-2812. DOI:10.1128/AEM.03987-12 |
| [16] | Park G, Takekawa M, Soda S, et al. Temperature dependence of nitrogen removal activity by anammox bacteria enriched at low temperatures[J]. Journal of Bioscience and Bioengineering, 2017, 123(4): 505-511. DOI:10.1016/j.jbiosc.2016.11.009 |
| [17] | Isaka K, Date Y, Kimura Y, et al. Nitrogen removal performance using anaerobic ammonium oxidation at low temperatures[J]. FEMS Microbiology Letters, 2008, 282(1): 32-38. DOI:10.1111/j.1574-6968.2008.01095.x |
| [18] | Zhu W Q, Li J, Dong H Y, et al. Nitrogen removal performance and operation strategy of anammox process under temperature shock[J]. Biodegradation, 2017, 28(4): 261-774. DOI:10.1007/s10532-017-9794-9 |
| [19] |
彭永臻, 张自杰. 活性污泥法处理系统中温度对动力学常数的影响[J]. 环境科学学报, 1990, 10(2): 226-232. Peng Y Z, Zhang Z J. Effects of temperature on the reaction kinetics of an activated sludge process[J]. Acta Scientiae Circumstantiae, 1990, 10(2): 226-232. DOI:10.3321/j.issn:0253-2468.1990.02.011 |
| [20] | He S L, Chen Y, Qin M, et al. Effects of temperature on anammox performance and community structure[J]. Bioresource Technology, 2018, 260: 186-195. DOI:10.1016/j.biortech.2018.03.090 |
| [21] | Guo Q, Xing B S, Li P, et al. Anaerobic ammonium oxidation (anammox) under realistic seasonal temperature variations: characteristics of biogranules and process performance[J]. Bioresource Technology, 2015, 192: 765-773. DOI:10.1016/j.biortech.2015.06.049 |
| [22] | Laureni M, Weissbrodt D G, Szivák I, et al. Activity and growth of anammox biomass on aerobically pre-treated municipal wastewater[J]. Water Research, 2015, 80: 325-336. DOI:10.1016/j.watres.2015.04.026 |
| [23] | Wang Y Y, Xie H C, Wang D L, et al. Insight into the response of anammox granule rheological intensity and size evolution to decreasing temperature and influent substrate concentration[J]. Water Research, 2019, 162: 258-268. DOI:10.1016/j.watres.2019.06.060 |
| [24] | Wang S Y, Peng Y Z, Ma B, et al. Anaerobic ammonium oxidation in traditional municipal wastewater treatment plants with low-strength ammonium loading: Widespread but overlooked[J]. Water Research, 2015, 84: 66-75. DOI:10.1016/j.watres.2015.07.005 |
| [25] | Jin R C, Ma C, Yu J J. Performance of an Anammox UASB reactor at high load and low ambient temperature[J]. Chemical Engineering Journal, 2013, 232: 17-25. DOI:10.1016/j.cej.2013.07.059 |
| [26] | Yang S F, Li X Y. Influences of extracellular polymeric substances (EPS) on the characteristics of activated sludge under non-steady-state conditions[J]. Process Biochemistry, 2009, 44(1): 91-96. DOI:10.1016/j.procbio.2008.09.010 |
| [27] | Parijs I, Steenackers H P. Competitive inter-species interactions underlie the increased antimicrobial tolerance in multispecies brewery biofilms[J]. The ISME Journal, 2018, 12(8): 2061-2075. DOI:10.1038/s41396-018-0146-5 |
| [28] | Palmer M W, Earls P G, Hoagland B W, et al. Quantitative tools for perfecting species lists[J]. Environmetrics, 2002, 13(2): 121-137. DOI:10.1002/env.516 |
| [29] | Winkler M K H, Kleerebezem R, Van Loosdrecht M C M V. Integration of anammox into the aerobic granular sludge process for main stream wastewater treatment at ambient temperatures[J]. Water Research, 2012, 46(1): 136-144. DOI:10.1016/j.watres.2011.10.034 |
| [30] | Hendrickx T L G, Kampman C, Zeeman G, et al. High specific activity for anammox bacteria enriched from activated sludge at 10℃[J]. Bioresource Technology, 2014, 163: 214-221. DOI:10.1016/j.biortech.2014.04.025 |
| [31] | Gong X F, Wang B, Qiao X, et al. Performance of the anammox process treating low-strength municipal wastewater under low temperatures: Effect of undulating seasonal temperature variation[J]. Bioresource Technology, 2020, 312. DOI:10.1016/j.biortech.2020.123590 |
| [32] | Reino C, Suárez-Ojeda M E, Pérez J, et al. Stable long-term operation of an upflow anammox sludge bed reactor at mainstream conditions[J]. Water Research, 2018, 128: 331-340. DOI:10.1016/j.watres.2017.10.058 |
| [33] | Dosta J, Fernández I, Vázquez-Padín J R, et al. Short-and long-term effects of temperature on the Anammox process[J]. Journal of Hazardous Materials, 2008, 154(1-3): 688-693. DOI:10.1016/j.jhazmat.2007.10.082 |
| [34] | Chen Y S, Zhao Z, Peng Y K, et al. Performance of a full-scale modified anaerobic/anoxic/oxic process: high-throughput sequence analysis of its microbial structures and their community functions[J]. Bioresource Technology, 2016, 220: 225-232. DOI:10.1016/j.biortech.2016.07.095 |
| [35] | Rattray J E, Van de Vossenberg J, Jaeschke A, et al. Impact of temperature on ladderane lipid distribution in anammox bacteria[J]. Applied and Environmental Microbiology, 2010, 76(5): 1596-1603. DOI:10.1128/AEM.01796-09 |
| [36] | Lin X M, Wang Y Y, Ma X, et al. Evidence of differential adaptation to decreased temperature by anammox bacteria[J]. Environmental Microbiology, 2018, 20(10): 3514-3528. DOI:10.1111/1462-2920.14306 |