 2021, Vol. 42
2021, Vol. 42



浮游植物是指在水中营浮游生活的微小植物, 其利用光能进行光合作用合成有机物[1].浮游植物是水生生态系统中的主要初级生产者, 是形成水生动物食物链的基础环节[2].浮游植物的组成和多样性变化将直接影响水生生态系统的结构与功能[3], 对湖泊[4]、江河[5]及湿地[6]等区域的生物多样性与生态稳定性起到至关重要的作用.由于浮游植物群落结构对水体环境敏感, 研究其群落结构组成及其多样性等指标能准确地反映水体状况, 可以弥补水体理化指标在水质评价上的不足[7], 是目前国际上对于水质评价和水体营养状况监测的重要研究方向[8~10].
哈尔滨城市河网处于松花江流域内, 是我国北部重要水运干线.每年6~8月为其丰水期, 降雨量为全年最大, 受地表径流影响, 在丰水期内河流水体受污染程度显著增高.随着经济的快速发展以及城市化进程的推进, 哈尔滨城市河网受人为活动与工农业污水废水排入的影响, 其水环境压力逐年升高.根据文献[11], 松花江哈尔滨段水质Ⅲ类和Ⅳ类居多.城市河网水生生态系统的稳定与沿岸城市生物多样性的保护、生物信息流动、能量循环及城市生态安全关系密切[12].城市河网对于城市的交通运输、环境美化、排污防洪及生物多样性保护等方面有着至关重要的作用, 它不仅拥有生态景观功能, 还维持着城市生态系统的平衡与稳定[13].哈尔滨城市河网由松花江哈尔滨段作为主要干流, 其中包含了景观水体(太阳岛)以及湿地(滨江、阿勒锦等), 3条河沟(何家沟、马家沟和阿什河)为其主要支流, 是承担哈尔滨市环境建设、排污与防洪等重要功能的水资源[14].目前关于哈尔滨城市河网的研究主要集中于生态规划[14]、流域水资源研究[15]和景观设计[16]等方面, 研究区域主要集中于松花江哈尔滨段干流上[17, 18], 对于阿什河流域[19, 20]及何家沟[21]等支流污染状况及生物多样性的相关研究较少涉及浮游植物, 尤其关于浮游植物多样性格局及环境相关性的报道尚缺乏系统性的生态学研究.
本研究以哈尔滨城市河网为主体, 分析沿着河网及人为活动干扰梯度下的浮游植物多样性分布格局; 基于多元统计分析, 详细阐述驱动浮游植物空间分布的主要环境因子; 并初步分析水体营养状态, 以期为城市河网生态修复及生物多样性保护提供生物学及生态学基础资料和理论依据.
1 材料与方法 1.1 采样点及采集周期本文以哈尔滨市区三环以内的城市河网系统为研究对象, 在松花江干流及市内汇入松花江的3条支流(何家沟、马家沟、阿什河)共设置了23个采样点.河网交织呈三叉戟状, 采样点几乎覆盖了整个哈尔滨城市的河网水域, 基本囊括了哈尔滨城市河网水域生态系统的典型特征, 具体设置如图 1所示, 共采集23个样点, 其中松花江(S)设置8个采样点位, 阿什河(A)5个点位, 何家沟(H)与马家沟(M)各设置5个采样点位.采样点使用GPS全球卫星定位系统进行定位并记录.样品的采集周期为2019年丰水期(6~8月), 每月采集一次.

|
图 1 哈尔滨城市河网采样点示意 Fig. 1 Sample sites in Harbin urban river network |
浮游植物样品的采集参照文献[22], 其中定性样品的采集使用25#浮游生物网进行采集, 使用时在水面下0.5 m左右呈“∞”字状捞取藻类, 现场加入4%~5%的甲醛溶液进行固定; 定量样品的采集使用5L容量的不锈钢采水器, 在水面下0.5 m处采集, 采集后置于聚乙烯瓶中, 用15 mL鲁格氏溶液固定, 静置24 h后用吸管小心吸取上清溶液, 至样品浓缩为20~30 mL.浮游植物鉴定时取均匀样品1 mL注入Sedgewick-Rafte浮游植物计数框中, 在光学显微镜下进行观察, 浮游植物的鉴定主要参考文献[23].
1.3 数据的采集、处理与分析 1.3.1 理化指标的采集现场使用YSI多参数水质监测仪对水体的水温(WT)、pH、溶解氧(DO)、电导率(Spc)和浊度(Tur.)进行测定并记录; 样品于24 h内在实验室内参照文献[24]测得总氮(TN)、总磷(TP)以及高锰酸盐指数.
1.3.2 营养状态指数(TSI)利用TSI营养状态指数对哈尔滨城市河网的营养状态进行评价.

|
TSI(高锰酸盐指数)3评价标准为:0 < TSI≤30为贫营养状态, 30 < TSI≤50为中营养状态, TSI>50为富营养状态.
1.3.3 优势种的判定优势种根据不同采样点的浮游植物优势度(Y)来判定.
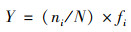
|
式中, ni为i种的个体数, N为所有种类总个体数, fi为该种出现的频率, Y值大于0.02的种类为优势种[25].
1.3.4 数据分析本实验采用Origin 2018进行绘图, 采样点地及浮游植物丰度时空分布特征使用AcrMap 10.2软件绘制.去趋势分析(DCA)使用软件CANOCO 4.5进行, 在排序前对解释变量和响应变量数据进行log(x+1)转换, 使样本总体趋于正态分布.通过对响应变量进行去趋势分析(detrended correspondence analysis, DCA)筛选排序模型, 若DCA分析中长度梯度大于4, 则单峰模型(canonical correspondence analysis, CCA)进行排序分析, 若长度梯度小于3, 则线性模型(redundancy analysis, RDA)可更有效地解释浮游生物生态分布特点, 长度梯度在3~4之间, 两种模型均可.使用SPSS 22.0进行独立样本t检验, 观察水体差异性.浮游植物群落结构分析使用Primer 5.0进行ANOSIM分析、SIMPER分析以及非度量多维尺度分析.
2 结果与分析 2.1 哈尔滨城市河网水体理化环境特征哈尔滨城市河网丰水期(6~8月)水体理化指标详见表 1.本研究期间内, WT变化范围在18.7~31.9℃之间, 平均温度24.5℃, 其中7月平均温度最高, 由于哈尔滨市2019年8月频繁降雨, 河网总体温度下降明显; 水体pH变化范围在6.4~9.39之间, 均值为7.69, 除距离排污口较近的样点外, 水体整体上为偏碱性或弱碱性; SpCond相对比较平稳; DO在6月时普遍高于7、8月; 水体Tur.逐月上升, 在8月最大平均值为97.07 mg·L-1; TN、TP与高锰酸盐指数随时间呈现上升趋势, 调查期间平均值分别为0.88、0.55和7.88 mg·L-1(表 1).独立样本t检验显示, WT与Tur.在6、7以及8月这3个月中差异性显著(P < 0.05或P < 0.01). pH、TN、TP和高锰酸盐指数在3个月中差异不显著, 但其在不同采样河段上差异性显著, 如松花江采样点(S1~S8)平均pH最高, 但TN、TP与高锰酸盐指数比其它采样点较低, 且TP在大部分样点间差异性显著(P < 0.05或P < 0.01).
|
|
表 1 哈尔滨城市河网丰水期环境因子均值1) Table 1 Mean value of environmental factors in the Harbin urban river network during the wet season |
有研究表明营养状态指数TSI除了可以对湖泊的营养状态进行评价, 对一些特定河流的营养状态评价也有较高信度[26], 故本研究运用各采样区域加权营养状态指数平均值对哈尔滨城市河网的水体营养状态进行初步评价, 结果如表 2所示.本研究期间内, 哈尔滨城市河网, 水体营养状态总体处于富营养状态.在空间上, 马家沟、阿什河富营养化程度相对较高, 松花江段TSI指数相对较低, 何家沟段、马家沟段和阿什河段采样点TSI指数沿流向升高, 在接近松花江汇合处达到最大(H2、M2和A3).而在时间上, 6月TSI变化范围最大, 最高值也出现在6月和7月TSI显著下降, 由于8月丰水期降雨量的升高, 雨水将河岸的有机污染物冲刷入河流, 使得8月TSI总体呈升高趋势(表 2).
|
|
表 2 哈尔滨城市河网平均营养状态指数评价结果 Table 2 Evaluation results of average trophic state indices in the Harbin urban river network |
2.2 浮游植物群落结构分布特征
本研究期间, 鉴定浮游植物149个分类单位(含变种和变形), 隶属于8个门、9个纲、19个目、34个科和57个属.在时间上, 6月浮游植物共鉴定8个门117个分类单位, 种类组成主要以绿藻门与硅藻门为主, 其中绿藻门54个种, 占浮游植物总数的58.28%; 硅藻门28个种, 占浮游植物总数的28.01%; 其他门类藻类占19.71%.7月与8月共鉴定浮游植物7个门132个分类单位, 与6月浮游植物种类不同, 7月和8月浮游植物主要以硅藻门和蓝藻门为主.在空间上, 不同采样河段的浮游植物组成不同, 松花江段浮游植物种类更为丰富, 有7个门134个种, 主要以硅藻门、绿藻门以及蓝藻门为主, 而其它河段(8个门84个种)大多以硅藻门、蓝藻门为主.
哈尔滨城市河网浮游植物平均丰度为6.54×106个·L-1, 变化范围在3.13×106~10.37×106个·L-1之间.时间上, 6月与7月、8月的浮游植物丰度差异性显著(P < 0.05), 尤其是绿藻门与蓝藻门的丰度差异显著(P < 0.05).其中蓝藻门相对丰度最大为65.96%, 其次绿藻门、硅藻门相对丰度为16.77%和15.74%.空间上, 松花江段平均丰度最高(9.71×106个·L-1), 其它支流平均丰度相对较低(3.59×106个·L-1).非度量多维尺度分析结果显示, 松花江段浮游植物丰度与其它河段具有一定的差异性(图 2), 且独立样本t检验也显示出, 松花江浮游植物丰度与何家沟(P < 0.05)、马家沟(P < 0.05)、阿什河(P < 0.01)存在着显著差异, 并且除马家沟与阿什河外(P>0.05), 3条支流之间差异显著(P < 0.05).

|
应力值stress为0.17 < 0.20, 表示结果可信 图 2 哈尔滨城市河网丰水期浮游植物丰度非度量多维尺度分析 Fig. 2 Non-metric multi-dimensional scale analysis of phytoplankton abundance in the Harbin urban river network during the wet season |
哈尔滨城市河网浮游植物优势种如图 3所示, 主要为硅藻门的梅尼小环藻(Cyclotella meneghiniana Kützing)、谷皮菱形藻[Nitzschia palea (Kützing) W.Smith]、肘状针杆藻[Synedra ulna (Nitzsch) Ehrenberg]和颗粒直链藻极狭变种(Melosira granulata var. angustissima Müller)等; 蓝藻门的湖泊假鱼腥藻[Pseudanabaena limnetica (Lemmermann) Komárek]、固氮鱼腥藻(Anabaena azotica Ley.)等; 绿藻门的狭形纤维藻(Ankistrodesmus angustus Bernard)和湖生卵囊藻(Oocystis lacustris Chodat)等.

|
(a)梅尼小环藻; (b)颗粒直链藻极狭变种; (c)谷皮菱形藻; (d)肘状针杆藻; (e)狭形纤维藻; (f)湖生卵囊藻; (g)卵形隐藻; (h)细小平裂藻; (i)湖泊假鱼腥藻; (j)固氮鱼腥藻 图 3 哈尔滨城市河网丰水期浮游植物相对丰度 Fig. 3 Relative abundance of phytoplankton in the Harbin urban river network during the wet season |
哈尔滨城市河网浮游植物群落的单因子交叉相似性分析(One-way crossed ANOSIM)显示, 采样区域(R=0.082, P<0.05)与月份(R=0.122, P < 0.05)均对浮游植物群落结构存在着显著影响, 并且松花江与其他支流之间浮游植物群落的结构与组成有显著差异(P < 0.05), 3条支流间差异性不显著(P>0.05, 表 3).SIMPER分析结果显示, 湖泊假鱼腥藻[Pseudanabaena limnetica (Lemmermann) Komárek]、卵形隐藻(Cryptomonas ovata Ehrenberg)、梅尼小环藻(Cyclotella meneghiniana Kützing)、谷皮菱形藻[Nitzschia palea (Kützing) W.Smith]和肘状针杆藻[Synedra ulna (Nitzsch) Ehrenberg]是采样区域浮游植物群落产生差异的主要贡献物种, 导致时间上浮游植物群落差异的主要贡献物种为梅尼小环藻(Cyclotella meneghiniana Kützing)、湖泊假鱼腥藻[Pseudanabaena limnetica (Lemmermann) Komárek]、卵形隐藻(Cryptomonas ovata Ehrenberg)以及谷皮菱形藻[Nitzschia palea (Kützing) W.Smith]和针状菱形藻[Nitzschia longissima (Brébisson) Ralfs].
|
|
表 3 哈尔滨城市河网浮游植物群落结构单因子交叉相似性分析及比较1) Table 3 Analysis and comparison of single-factor cross similarity of phytoplankton community structure in the Harbin urban river network |
2.3 排序分析
利用浮游植物优势种与WT、pH、SpCond、TP、TN、Tur.和高锰酸盐指数进行排序分析, 寻找影响哈尔滨城市河网丰水期浮游植物群落变化的主要环境因子.对丰水期浮游植物优势种丰度与理化因子数据进行去趋势分析, DCA结果显示在4个排序轴中最大长度梯度2.738, 因此使用线性模型冗余分析(RDA)进行排序能更准确地解释驱动哈尔滨城市河网丰水期主要驱动的环境因子.
RDA排序显示, 轴1与轴2的特征值分别为0.086和0.051, 优势种浮游生物与环境因子相关系数达到0.743和0.684, 前4个排序轴共解释了79.2%的累计种类环境因子百分比, 基本反映了在丰水期哈尔滨城市河网浮游植物群落与环境因子的生态关系.RDA排序分析可以看出[图 4(a)], 松花江采样点主要位于坐标轴的右侧, DO、pH与WT对其影响最大, 何家沟主要位于上半轴, DO是主要影响因子, 马家沟与阿什河位于左半轴, TP与Tur.是主要影响其浮游植物群落的环境因子.通过蒙特卡洛置换检验, 发现TP(F=4.449, P < 0.01)、pH(F=3.461, P < 0.01)是驱动哈尔滨城市河网浮游植物群落分布的主要环境因子, 这两条理化因子的线性长度均较长、平均长度接近, 说明这两种理化因子与优势种的总体相关性明显且差异较小.6月样点主要在轴上侧[图 4(b)], 证明6月对浮游植物群落最主要的环境贡献因子为DO、pH, 7月与8月大都分布在轴下半侧且分布不均匀, 表明7月和8月水体环境复杂, 其主要的环境贡献因子为高锰酸盐指数、Tur.以及TP.

|
图 4 哈尔滨城市河网丰水期浮游植物与环境因子的冗余分析排序 Fig. 4 Redundant analysis and ranking diagram of phytoplankton and environmental factors in Harbin urban river network during wet season |
浮游植物的空间分布格局与河网内水文条件、营养特征及人为活动干扰密切相关, 其分布特征是城市河网生态功能完整性与生物多样性的有效生物指示信号[4].本研究对哈尔滨城市河网内不同营养状态河流浮游植物的群落特征进行生态分析.研究期间观察到的哈尔滨城市河网优势种多为广布种, 其中, 硅藻门、蓝藻门以及绿藻门的种类为浮游植物群落组成的主要贡献者, 符合国内外河流丰水期浮游植物群落结构经典模式[27, 28].城市河网内浮游植物的群落构建受到流域内环境过滤和支流干流关系的共同影响, 其中河网内水体流向等空间过程被认为是驱动浮游植物群落生态分布的主要生态因素[2].本研究中群落结构相似性分析(ANOSIM)显示, 哈尔滨城市河网内主要干流松花江与支流的3条河流藻在浮游植物群落结构上存在显著差异性(表 3, P < 0.05).这不仅体现在群落结构、丰度及优势种组成上, 也体现在不同类群的时空分布特征上.这与国内外城市河网的浮游植物群落组成模式相类似, 进一步证明浮游植物群落的空间分布格局是温带城市河网水体理化特征变化及营养状态改变的有效生态指示信号.
松花江哈尔滨段浮游植物丰水期种类与丰度明显高于其他支流河段(图 3), 其优势种多为蓝藻门与绿藻门种类.硅藻门与蓝藻门种类则构成了何家沟、马家沟及阿什河的主要浮游植物类群, 其中, 梅尼小环藻与湖泊假鱼腥藻在3条支流大部分样点中均有出现.梅尼小环藻与湖泊假鱼腥藻被认为有较高的生态位宽度, 适应环境能力强, 它们也是河流浮游植物组成的常见种类[29, 30].浮游植物在水域生态系统中的生态位反映了个体在种群水平对环境过滤作用的响应与适应, 生态位宽度反映了水环境的稳定与健康状态.有研究表明不同生境间的水体理化差异会导致丝状藻类群落组成的变化[31], 本研究中干流松花江哈尔滨段主要优势物种为蓝藻门的湖泊假鱼腥藻和固氮鱼腥藻, ANOSIM分析表明, 湖泊假鱼腥藻为松花江与其它采样区域间的差异物种, 这是由于松花江哈尔滨段段采样点生境多为湿地、人工景观湖泊, 这些生境更有利于藻类尤其蓝藻的生长[32, 33], 本研究中样点S1、S2和S9为静止水体, 这类水体与流动水体的浮游植物群落结构不同, 进一步证明水体的流速是影响浮游植物的生长与定殖的环境过滤因子之一[34]. Reynold[35]的研究表明, 具有较强运动能力的硅藻与其它藻类相比具有资源竞争优势及污染耐受性, 并能在富营养条件下生长繁殖[36]. 在连通水系内, 水域仅受农业或城市化活动的影响, 则空间异质性较低[37].ANOSIM分析显示, 何家沟与马家沟、阿什河段浮游植物群落组成差异性不显著(P>0.05), 这3条支流的地理位置虽相似, 均汇入松花江, 但是其间并未有连通处, 因此3条支流生态环境相互独立.何家沟、马家沟位于城市中心区域, 具有主要的排污功能, 受到的人为因素与工业污染也更为严重, 而阿什河主要受到了农业与工业污染, 且阿什河段为高泥沙量河流, 在研究期间内, 由于降雨量增加导致3条支流水位上升, 为防止洪涝灾害而减少污水排放, 河流流速与流量加大并汇入松花江, 再加上松花江段采样点的地理性质, 也受到人为因素、漕运和渔业的影响, 使得松花江段营养盐含量与3条支流的差距不大.文献[6]表明水体自净能力的差异会导致水体浮游植物类群的差异性, 自净能力越强的水体的浮游植物群落所受理化因素的影响越小.由于城市河网水体的自净能力差异, 松花江段自净能力优于3条支流, 故对松花江段浮游植物群落的影响有限, 且主要对3条支流的浮游植物群落产生影响.本研究中, 何家沟、马家沟及阿什河样点均以运动能力较强的耐污种类谷皮菱形藻、肘状针杆藻和梅尼小环藻为主, 这与采集区域的水体营养状态及水文特征相吻合.硅藻门种类可耐受于高浑浊度水体, Zhao等[38]在高泥沙量的渭河及其支流中的研究中发现浮游群落组成以硅藻为主可能与其水体浑浊度有关, 且水体浑浊度是影响高含沙量河流浮游植物种群结构的主要因素, 这可能也是研究期间阿什河段硅藻门占据优势地位的主要原因之一.
3条支流的TSI指数相对较高, 物种多样性低, 优势种支配地位明显, 在整个研究期间都处于富营养状态(TSI>50, 表 2).与硅藻的密度可能会由于溪流中营养物的增加而升高的研究结果不同[39], 富营养化通常会引起蓝藻的增加, 随着时间推移, 蓝藻门的湖泊假鱼腥藻、固氮鱼腥藻等在3条支流中占据主要优势地位.由于蓝藻是一种喜温藻类[40], WT对蓝藻的生长产生的积极影响更为显著[41], 这也是蓝藻在研究期间占优势的主要原因.值得注意的是, 在本研究中, SIMPER分析表明不同采样区域与时间的浮游植物群落结构存在着显著差异, 造成差异的主要贡献物种为梅尼小环藻、湖泊假鱼腥藻、卵形隐藻与谷皮菱形藻.梅尼小环藻表面积体积之比大, 能快速吸收水体的营养物质[42], 且在流动水体中分布广泛.谷皮菱形藻作为一种污染指示种, 其相对丰度的增加与水体的高沉积物和总磷含量相关[43].卵形隐藻是一种具鞭毛藻类, 分布范围广, 在研究期间其丰度随着河流营养程度的增加而增加, 这种鞭毛藻类, 借助鞭毛寻找适宜环境吸收营养盐可以使其快速增长[42].在未来城市河网水生态研究中, 梅尼小环藻、谷皮菱形藻与卵形隐藻的生态分布格局是水体环境管理及生物多样性保护工作开展的有效生态指示信号.
3.2 河网水环境因子对浮游植物群落的影响哈尔滨城市河网生态环境存在一定空间异质性, 不同生境与环境因子的作用, 会产生具有差异性的浮游植物群落结构[44].有研究表明WT是丰水期影响浮游植物群落分布的关键环境因子, WT与浮游植物的生长关系密切[45], 可以控制浮游植物的基本特性, 引起浮游植物的碳固定与生物合成[46].WT与藻类丰度关系密切, 在本研究中, 通过独立样本t检验(表 1), 6月与7月、7月与8月之间WT的差异性显著(P < 0.01).6~8月丰水期WT上升, 蓝藻门丰度显著升高, 较以往在纬度相近地区的研究中以硅藻门种类为主不同[47], 蓝藻门所占比例加重[48], 证明WT的升高是不同月份之间浮游植物群落差异性的关键因素, 也是促进河流富营养化的潜在因子之一.WT升高虽然可以增强浮游植物的光合作用、呼吸作用以及其生长与繁殖, 但不同采样河段的浮游植物群落构建取决于水温和养分等环境因子的共同作用[49].本研究中RDA分析指示TP与pH是哈尔滨城市河网浮游植物群落演替的关键因子.pH是影响水体浮游植物活动的一项重要生态因素, 是水体化学性状和生命活动的综合反映[50].水体中浮游植物的生长发育会随着pH值降低而受到影响[51], 过高的pH值则会抑制浮游植物的光合作用, 而在pH值为7.75~8.75之间浮游植物的生物量会达到最高峰[52].RDA结果显示, pH对不同河流的影响程度不同[图 4(a)], 松花江段浮游植物群落组成受pH(6.24~9.39)值的影响最重, 这是由于在松花江段有很多的沉水植物, 沉水植物的光合作用会使水体pH上升[53], 而高pH的水环境也可能与蓝、绿藻的暴发有关[50], 这与在本研究期间松花江段浮游植物群落以蓝、绿藻占据主要优势地位相符, 这也表明pH是影响松花江段与3条支流之间浮游植物群落差异的主要环境因素之一.
磷是水体化学循环的重要元素, 淡水生态系统受到磷的限制[54], 在研究期间TP浓度在0.1~1.4mg·L-1范围变化, 丰水期一系列的强降雨导致径流的增加, 随着雨水的冲刷使得河岸泥土与有机污染物流入河流, 腐殖质与养分(氮和磷)从水生高等植物中释放出来, 使水体TP与TN随时间变化一直呈增高趋势, 于8月达到最大值.TN与TP升高的同时, 氮磷比下降, 有研究表明氮磷比影响着藻类的丰度, 尤其是蓝藻的暴发与氮磷比呈明显的负相关[55], 这与研究期间7月和8月蓝藻门丰度上升相符合.本研究的优势种湖泊假鱼腥藻、固氮鱼腥藻等蓝藻暴发的形成, TN起着至关重要的作用[56], 且高TN浓度也会导致绿藻更高效地生长且占据优势地位[35].梅尼小环藻与谷皮菱形藻等硅藻的生长与TP关系密切, TP的升高使得水体二氧化硅减少, 硅藻的生长与繁殖基于二氧化硅的供给[57].RDA结果显示了TP主要影响了3条支流的浮游群落组成, 且与松花江段不同, 3条支流的浮游植物群落中硅藻门占据很大比例, 这进一步印证了TP为3条支流浮游植物群落构建的关键环境因素, 这与以往研究[58]城市河网多受磷的影响相符合, 因此TP也是松花江干流与3条支流在城市河网内引起差异的主要原因之一.
从整体上来看, 在丰水期多雨情况下, 除了WT可以引起城市河网浮游植物在时间上的差异性, 河网内不同河段的水体环境因素的空间分布与浮游植物群落耦合也关系紧密.TP与pH共同作用, 对哈尔滨城市河网丰水期浮游植物群落的组成产生影响.梅尼小环藻、谷皮菱形藻等硅藻在3条支流中的广泛分布及湖泊假鱼腥藻、固氮鱼腥藻在松花江段的优势地位均与研究区域内pH和TP的分布格局存在一定耦合关系.
4 结论(1) 2019年哈尔滨城市河网浮游植物共鉴定出浮游植物149个分类单位, 浮游植物群落结构主要由硅藻门、绿藻门与蓝藻门种类构成, 优势种为梅尼小环藻、谷皮菱形藻和湖泊假鱼腥藻.
(2) 浮游植物群落时空变化显著.时间上, 6~8月蓝藻门逐渐占据优势, 浮游植物丰度在丰水期呈现随时间增高的趋势.在空间上差异性显著, 松花江河段以绿藻门、蓝藻门为主, 其它河段以硅藻门、蓝藻门为主, 松花江段浮游植物丰度显著高于其它河段, 但在丰水期松花江段浮游植物多样性降低, 环境稳定性有所下降.
(3) WT、TP和pH是丰水期影响哈尔滨城市河网浮游植物群落组成的主要环境因子, 其中松花江段主要受pH影响, 马家沟、何家沟及阿什河段受TP影响最大.
| [1] | 刘建康. 高级水生生物学[M]. 北京: 科学出版社, 1999. |
| [2] |
贾慧娟, 赖子尼, 王超. 珠三角河网浮游植物物种丰富度时空特征[J]. 生态学报, 2019, 39(11): 3816-3827. Jia H J, Lai Z N, Wang C. Temporal and spatial patterns of phytoplankton species richness in the Pearl River Delta[J]. Acta Ecologica Sinica, 2019, 39(11): 3816-3827. |
| [3] |
李然然, 章光新, 张蕾. 查干湖湿地浮游植物与环境因子关系的多元分析[J]. 生态学报, 2014, 34(10): 2663-2673. Li R R, Zhang G X, Zhang L. Multivariate analysis of the relations between phytoplankton assemblages and environmental factors in Chagan Lake Wetland[J]. Acta Ecologica Sinica, 2014, 34(10): 2663-2673. |
| [4] |
蔡阳, 陆欣鑫, 巴秋爽, 等. 镜泊湖春、夏两季浮游生物群落结构及其与环境因子的关系[J]. 海洋与湖沼, 2019, 50(1): 116-128. Cai Y, Lu X X, Ba Q S, et al. Plankton community structure in Jingpo Lake and the relationship with environmental factors[J]. Oceanologia et Limnologia Sinica, 2019, 50(1): 116-128. |
| [5] |
马煜, 陆欣鑫, 范亚文. 松花江哈尔滨段浮游植物群落格局及其与环境因子的相关性[J]. 生态学报, 2021, 41(1): 224-234. Ma Y, Lu X X, Fan Y W. Correlation between phytoplankton community patterns and environmental factors in Harbin section of the Songhua River[J]. Acta Ecologica Sinica, 2021, 41(1): 224-234. |
| [6] |
贺筱蓉, 李共国. 杭州西溪湿地首期工程区浮游植物群落结构及与水质关系[J]. 湖泊科学, 2009, 21(6): 795-800. He X R, Li G G. Community structure of phytoplankon and its relation with water quality in Xixi Wetland for the frist project aera, Hangzhou[J]. Journal of Lake Sciences, 2009, 21(6): 795-800. |
| [7] |
陆欣鑫, 刘妍, 范亚文. 呼兰河湿地夏、秋两季浮游植物功能分组演替及其驱动因子[J]. 生态学报, 2014, 34(5): 1264-1273. Lu X X, Liu Y, Fan Y W. Relationships between environmental variables and seasonal succession inphytoplankton functional groups in the Hulan River Wetland[J]. Acta Ecologica Sinica, 2014, 34(5): 1264-1273. |
| [8] |
任辉, 田恬, 杨宇峰, 等. 珠江口南沙河涌浮游植物群落结构时空变化及其与环境因子的关系[J]. 生态学报, 2017, 37(22): 7729-7740. Ren H, Tian T, Yang Y F, et al. Spatial and temporal distribution of phytoplankton community and its relationship with environment factors in Nansha's Rivers, Pearl River estuary[J]. Acta Ecologica Sinica, 2017, 37(22): 7729-7740. |
| [9] |
杨振雄, 董燕红, 娄全胜, 等. 一种用于浮游植物检测能力评估的比测方法[J]. 海洋环境科学, 2019, 38(5): 790-795. Yang Z X, Dong Y H, Lou Q S, et al. A case study on phytoplankton identifying proficiency test[J]. Marine Environmental Science, 2019, 38(5): 790-795. |
| [10] | David V, Tortajada S, Philippine O, et al. Ecological succession and resilience of plankton recovering from an acute disturbance in freshwater marshes[J]. Science of the Total Environment, 2020, 709. DOI:10.1016/j.scitotenv.2019.135997 |
| [11] | 中华人民共和国生态环境部. 2019中国生态环境状况公报[EB/OL]. http://www.mee.gov.cn/hjzl/sthjzk/zghjzkgb/, 2020-05-18. |
| [12] |
曹新向, 翟秋敏, 郭志永. 城市湿地生态系统服务功能及其保护[J]. 水土保持研究, 2005, 12(1): 145-148. Cao X X, Zhai Q M, Guo Z Y. Ecosystem services of urban wetland and its conservation[J]. Research of Soil and Water Conservation, 2005, 12(1): 145-148. DOI:10.3969/j.issn.1005-3409.2005.01.043 |
| [13] | 车越, 杨凯. 发挥河网调蓄功能消减城市雨洪灾害——基于传统生态智慧的思考[J]. 生态学报, 2016, 36(16): 4946-4948. |
| [14] |
原煜涵. 哈尔滨主城区生态网络规划策略研究[D]. 哈尔滨: 哈尔滨工业大学, 2012. Yuan Y H. The strategy research of ecological network planning in the main urban area of Harbin[D]. Harbin: Harbin Institute of Technology, 2012. |
| [15] |
娄德君, 刘玉莲, 王冀, 等. 松花江流域盛夏降水分布型的环流差异及影响机制[J]. 高原气象, 2020, 39(2): 239-242. Lou D J, Liu Y L, Wang J, et al. Circulation differences and influence mechanisms of distribution modes of midsummer precipitation in the Songhua River basin[J]. Plateau Meteorology, 2020, 39(2): 239-242. |
| [16] |
张玉娟, 何鲲, 李城林, 等. 松花江流域(哈尔滨段)景观敏感度时空变化分析[J]. 测绘与空间地理信息, 2019, 42(11): 5-7, 11, 16. Zhang Y J, He K, Li C L, et al. Temporal-spatial variation of landscape sensitivity in Songhua River basin (Harbin section)[J]. Geomatics & Spatial Information Technology, 2019, 42(11): 5-7, 11, 16. DOI:10.3969/j.issn.1672-5867.2019.11.002 |
| [17] |
魏南. 松花江哈尔滨段丰水期浮游植物多样性格局及环境相关性研究[J]. 黑龙江环境通报, 2018, 42(3): 86-91. Wei N. Relationship between phytoplankton diversity and environment in the Harbin section of Songhua River[J]. Heilongjiang Environmental Journal, 2018, 42(3): 86-91. DOI:10.3969/j.issn.1674-263X.2018.03.024 |
| [18] |
阎齐, 于洪贤. 松花江哈尔滨段夏季浮游植物群落结构与环境影响评价[J]. 水产学杂志, 2011, 24(2): 46-52. Yan Q, Yu H X. Study on the community structure of phytoplankton and environmental impact assessment in summer in Harbin section of Songhua River[J]. Chinese Journal of Fisheries, 2011, 24(2): 46-52. DOI:10.3969/j.issn.1005-3832.2011.02.011 |
| [19] |
马广文, 王业耀, 香宝, 等. 阿什河丰水期氮污染特征及其来源分析[J]. 环境科学与技术, 2014, 37(11): 116-120, 115. Ma G W, Wang Y Y, Xiang B, et al. Nitrogen pollution characteristics and source analysis in Ashi River during wet season[J]. Environmental Science & Technology, 2014, 37(11): 116-120, 115. |
| [20] |
马广文, 王业耀, 香宝, 等. 高纬区阿什河面源氮和磷污染输出特征[J]. 中国环境监测, 2017, 33(2): 47-54. Ma G W, Wang Y Y, Xiang B, et al. Non-point source pollution output characteristics of nitrogen and phosphorus in the Ashi River at high latitudes[J]. Environmental Monitoring in China, 2017, 33(2): 47-54. |
| [21] |
王力. 哈尔滨市何家沟污染源调查与综合整治研究[D]. 哈尔滨: 哈尔滨工业大学, 2013. Wang L. The investigation on Hejia Trench's pollution survey and comprehensive improvement[D]. Harbin: Harbin Institute of Technology, 2013. |
| [22] | SC/T 9402-2010, 淡水浮游生物调查技术规范[S]. |
| [23] | 胡鸿钧, 魏印心. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [24] |
王明翠, 刘雪芹, 张建辉. 湖泊富营养化评价方法及分级标准[J]. 中国环境监测, 2002, 18(5): 47-49. Wang M C, Liu X Q, Zhang J H. Evaluate method and classification standard on lake eutrophication[J]. Environmental Monitoring in China, 2002, 18(5): 47-49. DOI:10.3969/j.issn.1002-6002.2002.05.018 |
| [25] | Lampitt R S, Wishner K F, Turley C M, et al. Marine snow studies in the Northeast Atlantic Ocean: distribution, composition and role as a food source for migrating plankton[J]. Marine Biology, 1993, 116(4): 689-702. DOI:10.1007/BF00355486 |
| [26] |
马煜. 松花江哈尔滨段浮游植物群落结构动态变化与环境因子相关性研究[D]. 哈尔滨: 哈尔滨师范大学, 2020. Ma Y. Correlation between dynamic changes of phytoplankton community structure and environmental factors in Harbin section of the Songhua River[D]. Harbin: Harbin Normal University, 2020. |
| [27] | Belous Y P, Klochenko P D. Phytoplankton taxonomical structure in the middle part of Southern Bug River (Ukraine)[J]. International Journal on Algae, 2015, 17(3): 253-262. DOI:10.1615/InterJAlgae.v17.i3.60 |
| [28] | Zhong F, Wu J, Dai Y R, et al. Responses of water quality and phytoplankton assemblages to remediation projects in two hypereutrophic tributaries of Chaohu Lake[J]. Journal of Environmental Management, 2019, 248. DOI:10.1016/j.jenvman.2019.109276 |
| [29] |
王岳, 夏爽, 裴国凤. 长江干流湖北段浮游藻类群落结构特征[J]. 生态与农村环境学报, 2015, 31(6): 916-922. Wang Y, Xia S, Pei G F. Community structure of phytoplankton in the mainstream of the Yangtze River in Hubei[J]. Journal of Ecology and Rural Environment, 2015, 31(6): 916-922. |
| [30] |
君珊, 王东波, 周健华, 等. 拉萨河流域浮游植物群落结构特征及与环境因子的关系[J]. 生态学报, 2019, 39(3): 787-798. Jun S, Wang D B, Zhou J H, et al. Community structures of phytoplankton and its relationship with environmental factors in the Lhasa River[J]. Acta Ecologica Sinica, 2019, 39(3): 787-798. |
| [31] | Çelekli A, Arslanargun H, Soysal C, et al. Biochemical responses of filamentous algae in different aquatic ecosystems in South East Turkey and associated water quality parameters[J]. Ecotoxicology and Environmental Safety, 2016, 133: 403-412. DOI:10.1016/j.ecoenv.2016.08.002 |
| [32] |
周卢茜, 裘钱玲琳, 唐剑锋, 等. 城市湖泊春季绿藻水华特征及其影响因素——以宁波月湖为例[J]. 湖泊科学, 2019, 31(4): 1023-1034. Zhou L X, Qiu Q L L, Tang J F, et al. Characteristics of spring green algae blooms and their influencing factors in an urban lake, Moon Lake in Ningbo City, China[J]. Journal of Lake Sciences, 2019, 31(4): 1023-1034. |
| [33] |
范小晨, 代存芳, 陆欣鑫, 等. 金河湾城市湿地浮游植物功能类群演替及驱动因子[J]. 生态学报, 2018, 38(16): 5726-5738. Fan X C, Dai C F, Lu X X, et al. Study on phytoplankton functional group succession and driving parameters in the Jinhewan Urban Wetland[J]. Acta Ecologica Sinica, 2018, 38(16): 5726-5738. |
| [34] |
周静, 苟婷, 张洛红, 等. 流速对不同浮游藻类的生长影响研究[J]. 生态科学, 2018, 37(6): 75-82. Zhou J, Gou T, Zhang L H, et al. The effect of flow velocity on the growth of different phytoplankton[J]. Ecological Science, 2018, 37(6): 75-82. |
| [35] | Reynolds C S. The ecology of phytoplankton[M]. Cambridge: Cambridge University Press, 2006. |
| [36] | Barnett A, Méléder V, Blommaert L, et al. Growth form defines physiological photoprotective capacity in intertidal benthic diatoms[J]. The ISME Journal, 2015, 9(1): 32-45. DOI:10.1038/ismej.2014.105 |
| [37] | Özer T, Erkaya DI·A, Koçer M A T, et al. Spatial and temporal variations in composition of algae assemblages with environmental variables in an urban stream (Ankara, Turkey)[J]. Environmental Monitoring and Assessment, 2019, 191(6): 387. DOI:10.1007/s10661-019-7527-8 |
| [38] | Zhao G N, Pan B Z, Li Y P, et al. Phytoplankton in the heavy sediment-laden Weihe River and its tributaries from the northern foot of the Qinling Mountains: community structure and environmental drivers[J]. Environmental Science and Pollution Research, 2020, 27(9): 8359-8370. DOI:10.1007/s11356-019-07346-6 |
| [39] | Licursi M, Gómez N, Sabater S. Effects of nutrient enrichment on epipelic diatom assemblages in a nutrient-rich lowland stream, Pampa Region, Argentina[J]. Hydrobiologia, 2016, 766(1): 135-150. DOI:10.1007/s10750-015-2450-7 |
| [40] |
胡旻琪, 张玉超, 马荣华, 等. 巢湖2016年蓝藻水华时空分布及环境驱动力分析[J]. 环境科学, 2018, 39(11): 4925-4937. Hu M Q, Zhang Y C, Ma R H, et al. Spatial and temporal dynamics of floating algal blooms in Lake Chaohu in 2016 and their environmental drivers[J]. Environmental Science, 2018, 39(11): 4925-4937. |
| [41] | Elliott J A, Jones I D, Thackeray S J. Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake[J]. Hydrobiologia, 2006, 559(1): 401-411. DOI:10.1007/s10750-005-1233-y |
| [42] |
朱为菊, 庞婉婷, 尤庆敏, 等. 淮河流域春季浮游植物群落结构特征及其水质评价[J]. 湖泊科学, 2017, 29(3): 637-645. Zhu W J, Pang W T, You Q M, et al. Phytoplankton community structure and the evaluation of water quality in spring, Huaihe River basin[J]. Journal of Lake Sciences, 2017, 29(3): 637-645. |
| [43] | Licursi M, Gómez N. Effects of dredging on benthic diatom assemblages in a lowland stream[J]. Journal of Environmental Management, 2009, 90(2): 973-982. DOI:10.1016/j.jenvman.2008.03.004 |
| [44] |
王徐林, 张民, 殷进. 巢湖浮游藻类功能群的组成特性及其影响因素[J]. 湖泊科学, 2018, 30(2): 431-440. Wang X L, Zhang M, Yin J. Composition and influential factors of phytoplankton function groups in Lake Chaohu[J]. Journal of Lake Sciences, 2018, 30(2): 431-440. |
| [45] |
陆强, 陈慧丽, 邵晓阳, 等. 杭州西溪湿地大型底栖动物群落特征及与环境因子的关系[J]. 生态学报, 2013, 33(9): 2803-2815. Lu Q, Chen H L, Shao X Y, et al. Ecological characteristics of macrobenthic communities and its relationships with enviromental factors in Hangzhou Xixi Wetland[J]. Acta Ecologica Sinica, 2013, 33(9): 2803-2815. |
| [46] |
栾卓, 范亚文, 门晓宇. 松花江哈尔滨段水域硅藻植物群落及其水质的初步评价[J]. 湖泊科学, 2010, 22(1): 86-92. Luan Z, Fan Y W, Men X Y. Environmental quality evaluation of Songhua River in Harbin using diatom assemblages[J]. Journal of Lake Sciences, 2010, 22(1): 86-92. |
| [47] | Alam M G M, Jahan N, Thalib L, et al. Effects of environmental factors on the seasonally change of phytoplankton populations in a closed freshwater pond[J]. Environment International, 2002, 27(5): 363-371. |
| [48] | Heisler J, Glibert P M, Burkholder J M, et al. Eutrophication and harmful algal blooms: a scientific consensus[J]. Harmful Algae, 2008, 8(1): 3-13. DOI:10.1016/j.hal.2008.08.006 |
| [49] | Staehr P A, Sand-Jensen K. Seasonal changes in temperature and nutrient control of photosynthesis, respiration and growth of natural phytoplankton communities[J]. Freshwater Biology, 2006, 51(2): 249-262. DOI:10.1111/j.1365-2427.2005.01490.x |
| [50] |
苏发文. 大丰盐碱养殖虾塘浮游植物群落结构组成及其对水环境pH值的影响[D]. 上海: 上海海洋大学, 2016. Su F W. Community characteristics of microalgae and effect of the water environment pH in DaFeng Shrmip saline-alkalinity culture ponds[D]. Shanghai: Shanghai Ocean University, 2016. |
| [51] | Comeau S, Gorsky G, Jeffree R, et al. Key Arctic pelagic mollusc (Limacina helicina) threatened by ocean acidification[J]. Biogeosciences Discussions, 2009, 6: 2523-2537. |
| [52] |
马国红, 杜兴华, 段登选, 等. 盐碱地鱼池浮游植物与pH、总碱度、总硬度、含盐量的关系[J]. 齐鲁渔业, 2001, 18(5): 36-38. Ma G H, Du X H, Duan D X, et al. The relationship between phytoplandton and the pH, total alkalinity, total hardness and salinity in Salinizational pond[J]. Shandong Fisheries, 2001, 18(5): 36-38. |
| [53] |
赵联芳, 朱伟, 莫妙兴. 沉水植物对水体pH值的影响及其脱氮作用[J]. 水资源保护, 2008, 24(6): 64-67. Zhao L F, Zhu W, Mo M X. Effects of submerged macrophytes on pH values and nitrogen removal[J]. Water Resources Protection, 2008, 24(6): 64-67. DOI:10.3969/j.issn.1004-6933.2008.06.016 |
| [54] | Yang J, Wang F, Lu J P, et al. Interactive effects of temperature and nutrients on the phytoplankton community in an urban river in China[J]. Environmental Monitoring and Assessment, 2019, 191(11): 688. DOI:10.1007/s10661-019-7847-8 |
| [55] | Davis T W, Bullerjahn G S, Tuttle T, et al. Effects of increasing nitrogen and phosphorus concentrations on phytoplankton community growth and toxicity during Planktothrix blooms in Sandusky Bay, Lake Erie[J]. Environmental Science & Technology, 2015, 49(12): 7197-7207. |
| [56] | Ahn C Y, Oh H M, Park Y S. Evaluation of environmental factors on cyanobacterial bloom in eutrophic reservoir using artificial neural networks[J]. Journal of Phycology, 2011, 47(3): 495-504. DOI:10.1111/j.1529-8817.2011.00990.x |
| [57] | Schelske C L, Stoermer E F. Eutrophication, silica depletion, and predicted changes in algal quality in Lake Michigan[J]. Science, 1971, 173(3995): 423-424. DOI:10.1126/science.173.3995.423 |
| [58] | Hoellein T J, Arango C P, Zak Y. Spatial variability in nutrient concentration and biofilm nutrient limitation in an urban watershed[J]. Biogeochemistry, 2011, 106(2): 265-280. DOI:10.1007/s10533-011-9631-x |