 2021, Vol. 42
2021, Vol. 42



2. 长江水利委员会河湖保护与建设运行安全中心, 武汉 430010
2. Safety Center for River and Lake Protection, Construction and Operation of Yangtze River Water Resources Commissions, Wuhan 430010, China
罗红霉素(roxithromycin, ROX)作为一种大环内酯类抗生素, 被广泛运用于治疗呼吸道、泌尿和软组织感染[1].在中国每年使用量超过了150 t, ROX结构稳定, 且污水处理厂工艺去除效果有限[2~4].在我国太湖流域地表水中ROX检出浓度达到20.1 ng·L-1[5], 沉积物中含量(以dw计)为42.8 ng·g-1 [5], 饮用水水源地中达到3.5 ng·L-1 [6].据抗生素的累积动力学模型预测, 中国大沽河沉积物中ROX的含量在2100年会达到13 000 ng·g-1[7].Wang等[8]调查了1999~2019年污水处理厂尾水中抗生素的含量, 发现ROX的平均值为134 ng·L-1.医院废水中ROX达到了2 189 ng·L-1[9].水环境中, ROX广泛存在, 能够刺激藻类的生长[10], 影响大型溞的存活率[11].ROX及其代谢物能在鱼体内累积[5], 导致鱼类生化功能紊乱[12], 对水生态系统造成风险, 进一步危害人类健康[13].目前关于ROX对浮游动物的生长发育、生殖及生态行为方面的影响研究相对较少, 需进一步开展相关研究提高ROX对水生生物毒理效应认知.
本研究以标准模式生物大型溞作为受试生物[14], 以ROX急性毒性实验结果中96h-LC50的1‰和0.1‰为暴露浓度, 分析ROX对大型溞生长发育(体长、心率、胸肢跳动频率等)、生殖(产卵率等)和游泳等生态行为学指标, 测试大型溞抗氧化和代谢系统酶活性变化及相关基因的表达, 以期为ROX的生态毒理效应提供一定的依据.
1 材料与方法 1.1 受试生物受试生物大型溞(Daphnia magna)由中国武汉水生生物研究所提供.本实验开始前, 大型溞在实验室驯化14 d, 驯化条件包括培养介质: 曝气24 h的去离子水, 温度为(21±1)℃, 光暗周期16 h∶8 h, 光照强度为1000 lx.每天定时喂食纯种斜生栅藻(Scenedesmus obliquus).
1.2 实验试剂与仪器本实验试剂包括罗红霉素(ROX, CAS: 8024-83-1)和乙醇(色谱纯).本实验仪器包括培养箱、酶标仪、电视显微镜、液相色谱-质谱联用仪、荧光定量PCR仪、固相萃取、氮吹仪、色谱柱、定量浓缩仪、冰箱、高速冷冻离心机、高压灭菌锅、水浴锅、Mini-Q超纯水机、离心管和烧杯等.
1.3 急性毒性实验在预实验的基础上设置5、10、20、40、60、80和100 mg·L-1这7个浓度梯度, 同时设置清水空白对照组和溶剂对照组.在实验中, 往250 mL烧杯中加入100 mL目标浓度的暴露溶液, 每个烧杯加5只健康幼溞(6~24 h), 每个浓度设置4个平行.本实验周期为96 h, 在实验期间不进行喂食操作, 也不更换暴露溶液.在实验开始后24、48和96 h, 分别观察和记录每个烧杯中幼溞死亡的个数.
1.4 长期暴露实验根据急性毒性实验结果中96h-LC50的千分之一和十万分之一, 本实验设置了两个暴露浓度0.5 μg·L-1[实测浓度(0.412±0.052)μg·L-1]、50 μg·L-1[实测浓度(43.1±7.4)μg·L-1]以及清水空白对照组和溶剂对照组.在盛有50 mL暴露液的100 mL烧杯中随机放入1只健康幼溞(6~24 h), 每个暴露组设置20个平行, 用于生殖、生长及行为学实验.另在盛有800 mL暴露溶液的1 000 mL烧杯中随机放置150只健康幼溞(6~24 h), 每个暴露组设置3个平行, 用于抗氧化损伤实验和荧光定量PCR实验.本实验条件维持在(21±1)℃, 16 h∶8 h(光∶暗), 每天喂食斜生栅藻, 喂藻密度为5×104 cells·mL-1.每两天换水一次, 换水时清洗烧杯[11].母溞暴露周期为21 d, 暴露期间记录各项指标: ①母溞的产卵时间; ②产卵数量; ③ 21 d时母溞的体长; ④ 21 d时母溞1 min内胸枝、心脏跳动频率; ⑤ 21 d时游泳速度.将暴露结束后的溞放置在-80℃冰箱保存.测定SOD(超氧化物歧化酶)、CAT(过氧化氢酶)、GSH(谷胱甘肽)、GST(谷胱甘肽硫转移酶)、POD(过氧化物酶)、PPO(多酚氧化酶)、TPX(硫氧还蛋白过氧化物酶)和MDA(丙二醛).具体测定方法参照Wang等[15]和Aksakal等[16]的方法进行测定.用荧光定量PCR测定大型溞生殖、生长、胚胎发育、防御机制和氧化应激相关的基因表达情况.
1.4.1 种群增长指标测定内禀增长率的计算方法参照文献[17]中的计算公式:
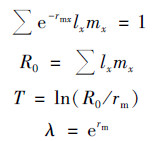
|
式中, rm为大型溞内禀增长率(ind·d-1, rm在粗略计算的基础上用逐步逼近法得出); x为大型溞的日龄(d); lx为第x天大型溞的存活率; mx为每只大型溞在第x天的产卵数(ind); R0为净增殖率(ind); T为世代时间(d); λ为周限增长率(ind·d-1).
1.4.2 生长、行为指标测定大型溞放在载玻片上, 加入适量的暴露溶液, 记录体长.然后用显微镜和录屏软件记录大型溞的胸枝和心脏跳动频率, 将大型溞放入24孔板中, 用ZebraLab软件测量出游泳轨迹, 计算出游泳速度.
1.4.3 生物标志物测定从空白对照、溶剂对照、0.5 μg·L-1和50 μg·L-1的暴露组的每个平行样品中取出50只大型溞进行测定.用生理盐水冲洗后, 将冲洗后的大型溞用滤纸吸干表面水分, 加入到已经称重的离心管中, 用电子天平准确称取大型溞的重量, 按重量(g)∶体积(mL)=1∶9的比例, 加入9倍体积的生理盐水, 在冰水浴条件下机械匀浆, 以2 500 r·min-1离心10 min, 取出上清液进行测定.测定的生物标志物包括SOD(超氧化物歧化酶)、CAT(过氧化氢酶)、GSH(谷胱甘肽)、GST(谷胱甘肽硫转移酶)、POD(过氧化物酶)、PPO(多酚氧化酶)、TPX(硫氧还蛋白过氧化物酶)和MDA(丙二醛).具体测定方法参照文献[15, 16]的方法进行.
1.4.4 荧光定量PCR从空白对照、溶剂对照、0.5 μg·L-1和50 μg·L-1的暴露组的每个平行样品中取出50只大型溞进行测定.将暴露21 d后的大型溞放入液氮中速冻3~5 min, 放入-80℃冰箱中保存.用大型溞基因组DNA提取试剂盒进行DNA提取, 用1.5%琼脂糖凝胶电泳进行DNA检测, 然后进行特异性PCR扩增.目的基因、引物设计和检测方法见文献[18, 19], 目的基因引物序列如表 1所示, 采用实时荧光定量PCR测定.使用QuantiTect Reverse Transcription Kit试剂盒进行逆转录.PCR扩增条件如下: 10 μL SYBR Green Mix、2 μL Primer Mix、3 μL Nuclease-free water和3 μL cDNA.反应参数如下: 在PCR中, 95℃ 3 min, 然后30个循环, 每次94℃ 30 s, 50℃ 30 s, 72℃ 30 s.熔解曲线验证了扩增特异性.每组重复3次, 计算3次重复的平均值.所有结果用两个内参基因(如actin和gapdh)的mRNA水平和Cq值进行归一化.
|
|
表 1 目的基因及引物序列 Table 1 Target gene and primer sequences |
2 结果与讨论 2.1 半致死效应
ROX对大型溞急性毒性的剂量-效应曲线如图 1所示(5 mg·L-1和10 mg·L-1暴露时未出现死亡, 图 1中不包含这两个浓度).在实验过程中, 空白对照组和溶剂对照组均未出现明显死亡(死亡率<5%).大型溞暴露于20~100 mg·L-1 5个浓度组时, 均出现大型溞死亡现象, 且随着暴露浓度的增加, 死亡率不断提高, 通过线性回归计算, ROX对大型溞的48h-LC50为60.26 mg·L-1, 96h-LC50为39.81 mg·L-1.

|
图 1 暴露48 h和96 h后ROX对大型溞的致死效应 Fig. 1 Toxic effect of ROX on Daphnia magna after 48 h and 96 h |
根据化学品分类和标签规范第28部分: 对水生环境的危害(GB 30000.28-2013)的毒性等级划分[20], ROX对大型溞的急性毒性等级为Ⅲ级(10~100 mg·L-1).ROX对鳜鱼的96 h半致死浓度(96h-LC50)为51.64 mg·L-1[21], 对大型溞的48 h半致死浓度(48h-LC50)在74.3 mg·L-1[22].但ROX对绿藻产生较高的生态毒性, 96h-LC50为4.663 mg·L-1[21], 水环境中ROX对初级生产者的毒理效应明显高于消费者.
2.2 对大型溞生殖的影响ROX对大型溞生殖指标的影响如表 2所示.空白对照组和溶剂对照组的数据无明显差异, 以下数据分析以溶剂暴露为参照组, 高低两个ROX暴露组对大型溞的生殖能力产生一定的影响, 低浓度组轻微延长了大型溞性成熟时间, 提升了首胎产卵个数, 而高浓度组与此相反, 高浓度组明显缩短了大型溞性成熟时间, 首胎产卵个数明显减少.从21 d暴露来看, 高低两个浓度组ROX都增加了大型溞的产卵胎次、产卵总量和平均每胎产卵量, 高浓度组表现更为突出.从暴露过程来看, 暴露初期低浓度ROX能够刺激大型溞的生殖能力, 而高浓度ROX对大型溞生殖能力产生一定的抑制作用, 但随着暴露时间的延长, 大型溞在环境胁迫下产生生态补偿, 导致产卵量显著提升.研究表明, 大型溞在低浓度污染下, 为了维持自身种群正常的繁衍能力, 能够通过调节性成熟时间和单胎产卵数量, 弥补环境压力造成的环境胁迫[23].
|
|
表 2 ROX对大型溞生殖指标的影响1) Table 2 Influence of ROX on the reproductive index of Daphnia magna |
2.3 对大型溞种群增长的影响
从ROX对大型溞种群增长指标的影响来看(表 3), 高浓度组的净生殖率、周限增长率和内禀增长率均显著升高.内禀增长率是表征污染物对大型溞种群生育力、寿命和发育速率毒性影响的综合指标[24].在高低两个ROX暴露浓度下, 大型溞种群生殖能力都得到提升, 高浓度组提升了57%.Tessier等[25]的研究发现大型溞种群增长率较大时, 生存所需的最低食物水平相对较高, 而自然环境中食物有限, 过度增殖反而容易引起大型溞因饥饿而死亡[25].根据衰老的进化理论, 维持身体机能所需的代谢成本和生殖所需的代谢成本保持着平衡, 过度生殖会消耗促进身体代谢的资源, 最终缩短寿命[26].大型溞生殖率的上升, 意味着寿命可能会缩减, 生存所需的最低食物水平也会提升, 最终导致种群生殖和寿命受到影响[25, 26].
|
|
表 3 ROX对大型溞种群增长指标的影响1) Table 3 Influence of ROX on the population growth index of Daphnia magna |
2.4 对大型溞生长发育和生态行为的影响
大型溞心率、胸肢跳动频率和体长与大型溞摄食、呼吸、代谢和内分泌系统等健康状况密切相关.从图 2可以看出, 两个暴露组均显著抑制了大型溞的体长生长, 提高了大型溞心脏跳动频次, 高浓度组诱导率超过了45%.从图 3可以看出, 两个暴露组均对游速产生抑制作用, 高浓度组抑制率更高.对大型溞胸肢跳动频率略有诱导作用, 但诱导不显著.

|
*表示显著性检验, P < 0.05 图 2 21 d暴露后ROX对大型溞的体长、胸肢跳动频率、心率和游速的影响 Fig. 2 Effect of ROX on body length, thoracic limb activity, heart rate, and swimming speed of Daphnia magna after exposure for 21 d |

|
每列3个重复,正方形代表起点,圆形代表终点 图 3 1 d暴露后大型溞的游泳轨迹 Fig. 3 Swimming track map of Daphnia magna after exposure for 21 d |
ROX对大型溞心率的影响鲜有报道, 心率是反映污染物对大型溞血液循环系统影响程度的敏感生理指标[27], 心率紊乱说明器官可能受到损伤, 导致大型溞呼吸能力下降[28], 且健康状况易受水体病原体侵扰[29].胸肢是大型溞的摄食器官, 它完成摄食动作受心脏能量供给、神经系统调控的双重作用.心率过快一定程度上增加了胸肢跳动的能量供应, 能量的过度输出, 易引起生长抑制、发育迟缓.大型溞体长的变化也是环境胁迫下生物体的一种防御机制, 但污染物影响下执行的错误防御策略可能会诱导死亡, 即使体长的改变很小, 在实际生态环境中, 也可能会影响大型溞的种群稳定[30].有研究报道400 μg·L-1硫氰酸红霉素(ET)使得大型溞的体长减短[31], 1 mg·L-1四环素显著缩短了大型溞的体长[32].随着暴露浓度增加, 大型溞游泳能力不断受到抑制, 最高浓度组呈现显著抑制效应, 从大型溞的游泳轨迹可以看出高低浓度组ROX显著降低了大型溞的泳道密度, ROX对大型溞游泳活性产生的抑制作用可能是ROX和运动神经中控制第二触角肌肉的钠离子通道相互作用的结果[29].
2.5 对大型溞抗氧化系统的影响大型溞在应对非生物胁迫时有两类抗氧化防御体系, 一类是酶促防御系统, 包括POD、SOD、CAT和GST等; 另一类是非酶促防御系统, 包括GSH和抗坏血酸等.为进一步研究ROX对大型溞抗氧化系统的影响, 检测了大型溞抗氧化系统中TPX、PPO、SOD、POD、CAT、GST、GSH-Px活性和MDA含量.
从图 4可以看出, 高低两个暴露组ROX都显著增加了MDA的含量, 高浓度组对MDA的诱导率甚至超过了100%; 显著抑制了CAT、GST、POD和PPO的活性, 高浓度组对POD和CAT活性的抑制接近50%, 抑制作用随浓度增加显著增强.低浓度组ROX对SOD、TPX和GSH-Px的活性无明显改变, 高浓度组产生显著抑制作用, 对GSH-Px活性的抑制超过45%.

|
*表示显著性检验, P < 0.05, 下同 图 4 21 d暴露后ROX对大型溞TPX、PPO、SOD、CAT、POD、GST、GSH-Px活性和MDA含量的影响 Fig. 4 Effect of ROX on TPX, PPO, SOD, CAT, POD, GST, GSH-Px activity, and MDA content of Daphnia magna after expoure for 21 d |
正常生理状态下, 大型溞体内的活性氧产生和清除处于动态平衡, ROX的存在刺激大型溞体内的氧化还原状态趋向氧化[16].活性氧族分子(ROS)是影响生物体内氧化还原稳态的主要因子, 包括过氧化氢(H2O2)、羟自由基(·OH)、和超氧自由基(·O2-)等.TPX和PPO作为抗氧化酶, 能够在生物中发挥抗逆性功能, 阻止活性氧产生, 维持体内正常的氧化还原平衡[33], TPX可通过分解烷基过氧化物和过氧化氢减少羟自由基的产生从而实现抗氧化活性.实验结果表明ROX胁迫产生的ROS消耗了PPO和TPX, 且ROX浓度越高, 导致大型溞体内PPO和TPX活性水平越低, TPX和PPO在大型溞对ROS的防御系统中发挥着重要作用.TPX活性失常能够通过氧化还原反应、信号传导、细胞凋亡等途径影响生物体正常生理功能.
在抗氧化系统中, 细胞内的抗氧化酶SOD、CAT、GSH-Px、POD和GST相互协同, 共同完成抗氧化作用.SOD能够歧化超氧化物自由基形成H2O2和O2[34].CAT和POD是一种H2O2去除剂, 催化H2O2分解[16, 35].GST催化GSH与底物结合, 在氧化应激因子的解毒中发挥重要作用[16].本研究中检测的所有酶促系统酶活性显著降低, 而脂质过氧化产物MDA含量显著提升.MDA是表征生物体内ROS水平的重要指标, 代表了机体的膜脂质过氧化水平[16].氧自由基剧增和抗氧化体系的还原能力减弱是大型溞产生氧化应激的主要原因, 过量的活性氧会对不饱和脂肪酸氧化或者过氧化, 生成脂质过氧化产物MDA[36, 37].生物膜中不饱和脂肪酸减少, 膜的流动性降低, 通透性发生改变, 进一步造成营养物吸收效率降低, 维生素、氨基酸和无机盐等营养失衡, 最终造成生长抑制、生殖能力降低和免疫力低下等[38].
有研究发现暴露于5 μg·L-1 ROX中大型溞的SOD和CAT活性下降[39].高浓度ROX暴露后, 大型溞体内产生了过量的活性氧, 导致SOD、CAT、POD、GST和GSH-Px活性被显著抑制, 无法抵抗氧化胁迫, 这时ROS产生的速率大于抗氧化酶清除的速率, 导致ROS积累过多对机体产生损害, 会抑制抗氧化酶的生成[39].Wang等[15]的研究发现暴露于50 μg·L-1布洛芬6 h和48 h后MDA含量显著增加[15], 说明有一定程度的脂质过氧化损伤.抗氧化剂在中断脂质过氧化和过氧化物的转化中起着非常重要的作用[15, 40], 但由于ROX浓度的增加, 破坏了抗氧化剂清除和活性氧的产生之间的平衡, 因此过多的活性氧积累, 引起细胞膜损伤[15, 41].
2.6 对大型溞基因转录的影响在环境污染物的作用下, 生物体在基因水平、蛋白水平、细胞水平和组织水平等方面均会有响应[42].基因表达水平检测是表征环境污染物毒性作用的有效手段.本文研究了ROX在大型溞抗氧化酶GST、过氧化物酶POD相关以及和防御机制有关的基因表达水平的毒性作用.现有研究发现抗生素类污染物会影响水生生物甾体生成途径, 导致其分泌紊乱[43].为此, 本研究还检测了甲状腺激素和保幼激素相关基因表达.
2.6.1 对vg1、vg2、和vit-2转录的影响卵黄蛋白(vitellogenin)常被用作水生物种的内分泌干扰生物标志物, 是胚胎发育的能量储备[44]和主要内源性营养[45], 雌激素控制卵黄蛋白的产生, 因此卵黄蛋白的产生及其编码基因(vg1和vg2)的表达常被用作雌性激素暴露的生物标记物[46].卵黄蛋白vg1有助于卵黄的形成, 卵黄蛋白vg2除了有助于卵黄的形成之外, 还可以促进其他的生理功能[47].vit-2是卵黄蛋白原的编码基因之一, 卵黄蛋白原是卵黄蛋白的前体[19].DePina等[26]的研究发现秀丽隐杆线虫在正常生长的情况下, vit-2的相对表达量会随着寿命的增长而减少.本实验发现两个浓度组vg1和vg2表达下调(图 5), 高浓度组vit-2表达下调了440%.虽然随着生殖的进行, 对照组vit、vg1和vg2相对表达量也会减少, 但仍处于上调, ROX暴露组21 d时大型溞vg1、vg2和vit-2的相对表达量减为负值(下调), 说明ROX在大型溞成年早期vg1、vg2和vit-2的转录水平可能相对偏高, 以满足生殖需求[26], 暴露后期21 d时, 因ROX负面效应, vg1、vg2和vit-2的表达下调[26], 生殖力下降.在成年早期, 过量的vit转录可以确保充分的卵黄蛋白合成能力, 以达到最大的生殖率[26].

|
图 5 21 d暴露后ROX对大型溞vg1、vg2和vit-2基因相对转录量的影响 Fig. 5 Effect of ROX on relative gene transcription of vg1, vg2, and vit-2 in Daphnia magna after exposure for 21 d |
甲壳类动物的生殖发育与蜕皮有关, 主要受蜕皮激素(20-hydroxyecdysone)和保幼激素(JH)以及ecr核受体和usp的转录调控[48].保幼激素可以调控大型溞卵黄蛋白的形成[48], 主要参与生物的生殖、变态和发育过程[49], jhe(juvenile hormone esterase)是保幼激素的编码基因.此外, ecra (ecdysone receptor a)、ecrb (ecdysone receptor b)和rxr也是影响大型溞生长的基因[18].ecrb参与调节大型溞的蜕皮过程[50], 在苏拉威西秀体溞中ecra参与成虫期的蜕皮和生殖过程[51].rxr是视黄醇类x受体(RXR)的编码基因, RXR在甲状腺激素受体对下丘脑-垂体轴基因的调控中起协同调节作用[52].从图 6可以看出, 和对照组相比, 高浓度组jhe、ecra、ecrb和rxr表达下调, 表明大型溞的蜕皮受到抑制, 体内的保幼激素、甲状腺激素分泌也会受到影响.在正常环境下, 大型溞每3 d进行一次单性生殖, 生殖周期与蜕皮周期同步[53].蜕皮后, 母溞将新卵从卵巢排出到孵育囊[53], 生殖周期结束时, 母溞会将幼崽从孵育囊中释放到周围的水中.随后, 母溞再次蜕皮, 并重复这一循环[53].甲壳类动物的蜕皮周期对它们的生理过程十分重要, 如生长、成年和生殖[54].本实验表明在暴露初期大型溞的生殖力被显著提升, 导致生殖需求下降, 后期的生殖力大大减弱, 体长也显著小于对照组, 生长发育受到影响.

|
图 6 21 d暴露后ROX对大型溞jhe、ecra、ecrb和rxr基因相对转录量的影响 Fig. 6 Effect of ROX on relative gene transcription of jhe, ecra, ecrb, and rxr in Daphnia magna after exposure for 21 d |
一氧化氮(NO)是一种重要的效应分子, 参与多种生物过程, 包括对寄生虫、病毒和细菌等病原体的免疫应答[18, 55].在有核细胞中的诱导型一氧化氮合酶(iNOS)会产生NO来抑制或杀死细胞内的病原体[55].药物通过刺激iNOS的编码基因(nos2)参与iNOS的上调, 加剧NO介导的DNA损伤[56].从图 7可以看出, 暴露于ROX后, 大型溞nos2表达上调, 意味着iNOS上调, NO会不断生成, 发挥对病原体的免疫应答作用, 但是NO的持续生成会导致缺氧, 神经元死亡[57].

|
图 7 21 d暴露后ROX对大型溞nos2、gst、gst-theta和per06基因相对转录量的影响 Fig. 7 Effect of ROX on relative gene transcription of nos2, gst, gst-theta and per06 in Daphnia magna after exposure for 21 d |
GST是水生生物中氧化应激的敏感性指标, 其与谷胱甘肽(GSH)结合, 参与细胞解毒[58].高浓度组ROX暴露导致大型溞gst表达下调, gst-theta下调了249%, 而低浓度组变化不显著, 表明高浓度的ROX导致大型溞体内解毒作用减弱[18], 这与高浓度组导致GST和GSH-Px活性被抑制的结果相对应.ROX的污染胁迫超过一个限值后, 开始抑制抗氧化酶的生成[39].per06是过氧化物酶(POD)的编码基因[19], 对有毒物质所引起的氧化应激具有保护作用[59], 可以清除磷脂氢过氧化物[60], 在抗氧化防御中发挥重要作用[60].高低浓度组per06表达均显著下调, 高浓度组甚至下调了581%, 意味着大型溞体内积累过多ROS抑制了过氧化物酶的活性, 最终会降低生殖能力, 抑制生长发育, 导致免疫力低下.
3 结论(1) ROX对大型溞的急性毒性等级为Ⅲ级, 48h-LC50为60.26 mg·L-1, 96h-LC50为39.81 mg·L-1.
(2) 在暴露初期ROX导致大型溞内禀增长率显著升高, 生殖能力显著升高, 暴露后期大型溞vg1、vg2和vit-2表达下调, 个体的生殖能力降低, 影响大型溞的寿命.
(3) ROX引起保幼激素、甲状腺激素和蜕皮激素相关的基因jhe、ecra、ecrb和rxr表达下调, 大型溞体长变短, 心跳升高, 游速变慢, 生长发育受到影响.
(4) ROX下调大型溞GST和POD的编码基因gst、gst-theta和per06表达, 上调自由基编码基因nos2表达, 过多自由基积累导致CAT、GST、POD、PPO、SOD、TPX和GSH-Px等酶的活性抑制, MDA的含量增加, 引起细胞膜损伤, 最终导致免疫力低下和生长抑制, 影响种群稳定.
| [1] | Li W, Xu X J, Lyu B L, et al. Degradation of typical macrolide antibiotic roxithromycin by hydroxyl radical: kinetics, products, and toxicity assessment[J]. Environmental Science and Pollution Research, 2019, 26(14): 14570-14582. DOI:10.1007/s11356-019-04713-1 |
| [2] | Dong H Y, Yuan X J, Wang W D, et al. Occurrence and removal of antibiotics in ecological and conventional wastewater treatment processes: a field study[J]. Journal of Environmental Management, 2016, 178: 11-19. |
| [3] | Du J, Fan Y, Qian X. Occurrence and behavior of pharmaceuticals in sewage treatment plants in eastern China[J]. Frontiers of Environmental Science & Engineering, 2015, 9(4): 725-730. DOI:10.1007/s11783-014-0661-1 |
| [4] |
周志洪, 赵建亮, 魏晓东, 等. 珠江广州段水体抗生素的复合污染特征及其生态风险[J]. 生态环境学报, 2017, 26(6): 1034-1041. Zhou Z H, Zhao J L, Wei X D, et al. Co-occurrence and ecological risk of antibiotics in surface water of Guangzhou section of Pearl River[J]. Ecology and Environmental Sciences, 2017, 26(6): 1034-1041. |
| [5] | Xie Z X, Lu G H, Liu J C, et al. Occurrence, bioaccumulation, and trophic magnification of pharmaceutically active compounds in Taihu Lake, China[J]. Chemosphere, 2015, 138: 140-147. DOI:10.1016/j.chemosphere.2015.05.086 |
| [6] | Lin T, Yu S L, Chen W. Occurrence, removal and risk assessment of pharmaceutical and personal care products (PPCPs) in an advanced drinking water treatment plant (ADWTP) around Taihu Lake in China[J]. Chemosphere, 2016, 152: 1-9. DOI:10.1016/j.chemosphere.2016.02.109 |
| [7] | Hu X G, He K X, Zhou Q X. Occurrence, accumulation, attenuation and priority of typical antibiotics in sediments based on long-term field and modeling studies[J]. Journal of Hazardous Materials, 2012, 225-226: 91-98. DOI:10.1016/j.jhazmat.2012.04.062 |
| [8] | Wang J L, Chu L B, Wojnárovits L, et al. Occurrence and fate of antibiotics, antibiotic resistant genes (ARGs) and antibiotic resistant bacteria (ARB) in municipal wastewater treatment plant: an overview[J]. Science of the Total Environment, 2020, 744. DOI:10.1016/j.scitotenv.2020.140997 |
| [9] | Chang X S, Meyer M T, Liu X Y, et al. Determination of antibiotics in sewage from hospitals, nursery and slaughter house, wastewater treatment plant and source water in Chongqing region of Three Gorge Reservoir in China[J]. Environmental Pollution, 2010, 158(5): 1444-1450. DOI:10.1016/j.envpol.2009.12.034 |
| [10] | Yang L H, Ying G G, Su H C, et al. Growth-inhibiting effects of 12 antibacterial agents and their mixtures on the freshwater microalga Pseudokirchneriella subcapitata[J]. Environmental Toxicology and Chemistry, 2008, 27(5): 1201-1208. DOI:10.1897/07-471.1 |
| [11] | Zhang P, Yan Z H, Lu G H, et al. Single and combined effects of microplastics and roxithromycin on Daphnia magna[J]. Environmental Science and Pollution Research, 2019, 26(17): 17010-17020. DOI:10.1007/s11356-019-05031-2 |
| [12] | Liu J C, Lu G H, Wang Y H, et al. Bioconcentration, metabolism, and biomarker responses in freshwater fish Carassius auratus exposed to roxithromycin[J]. Chemosphere, 2014, 99: 102-108. DOI:10.1016/j.chemosphere.2013.10.036 |
| [13] | Chen H, Liu S, Xu X R, et al. Antibiotics in typical marine aquaculture farms surrounding Hailing Island, south China: occurrence, bioaccumulation and human dietary exposure[J]. Marine Pollution Bulletin, 2015, 90(1-2): 181-187. DOI:10.1016/j.marpolbul.2014.10.053 |
| [14] | Lu G H, Li Z H, Liu J C. Effects of selected pharmaceuticals on growth, reproduction and feeding of Daphnia magna[J]. Fresenius Environmental Bulletin, 2013, 22(9): 2583-2589. |
| [15] | Wang L, Peng Y, Nie X P, et al. Gene response of CYP360A, CYP314, and GST and whole-organism changes in Daphnia magna exposed to ibuprofen[J]. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2016, 179: 49-56. |
| [16] | Aksakal F I. Evaluation of boscalid toxicity on Daphnia magna by using antioxidant enzyme activities, the expression of genes related to antioxidant and detoxification systems, and life-history parameters[J]. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2020, 237. DOI:10.1016/j.cbpc.2020.108830 |
| [17] |
翁显龙, 左迪, 李嘉尧, 等. 温度及pH对隆线溞生长和生殖的影响[J]. 动物学杂志, 2011, 46(1): 1-6. Weng X L, Zuo D, Li J Y, et al. Effects of temperature and pH on the population dynamics parameters of Daphnia carinata[J]. Chinese Journal of Zoology, 2011, 46(1): 1-6. DOI:10.3969/j.issn.1671.7856.2011.01 |
| [18] | Houde M, Douville M, Gagnon P, et al. Exposure of Daphnia magna to trichloroethylene (TCE) and vinyl chloride (VC): evaluation of gene transcription, cellular activity, and life-history parameters[J]. Ecotoxicology and Environmental Safety, 2015, 116: 10-18. DOI:10.1016/j.ecoenv.2015.02.031 |
| [19] | Vandegehuchte M B, Vandenbrouck T, De Coninck D, et al. Can metal stress induce transferable changes in gene transcription in Daphnia magna?[J]. Aquatic Toxicology, 2010, 97(3): 188-195. DOI:10.1016/j.aquatox.2009.07.013 |
| [20] |
郭婧颖, 刘建超, 李帅衡, 等. 双酚AF对大型溞生殖、生长等生态行为的影响[J]. 中国环境科学, 2019, 39(10): 4394-4400. Guo J Y, Liu J C, Li S H, et al. Influences of bisphenol AF on the reproduction and growth of Daphnia magna[J]. China Environmental Science, 2019, 39(10): 4394-4400. DOI:10.3969/j.issn.1000-6923.2019.10.044 |
| [21] | Ding H J, Wu Y X, Zhang W H, et al. Occurrence, distribution, and risk assessment of antibiotics in the surface water of Poyang Lake, the largest freshwater lake in China[J]. Chemosphere, 2017, 184: 137-147. DOI:10.1016/j.chemosphere.2017.05.148 |
| [22] | Choi K, Kim Y, Jung J, et al. Occurrences and ecological risks of roxithromycin, trimethoprim, and chloramphenicol in the Han River, Korea[J]. Environmental Toxicology and Chemistry, 2008, 27(3): 711-719. DOI:10.1897/07-143.1 |
| [23] |
张洪昌, 陈良燕, 刘树深, 等. BPA与3种双酚类化合物的联合作用雌激素效应[J]. 环境科学, 2009, 30(1): 260-265. Zhang H C, Chen L Y, Liu S S, et al. Jointed estrogenic activities of Bisphenol A and three of its analogs[J]. Environmental Science, 2009, 30(1): 260-265. DOI:10.3321/j.issn:0250-3301.2009.01.044 |
| [24] | Xi Y L, Hagiwara A, Sakakura Y. Combined effects of food level and temperature on life table demography of Moina macrocopaStraus (Cladocera)[J]. International Review of Hydrobiology, 2005, 90(5-6): 546-554. DOI:10.1002/iroh.200510809 |
| [25] | Tessier A J, Woodruff P. Trading off the ability to exploit rich versus poor food quality[J]. Ecology Letters, 2002, 5(5): 685-692. DOI:10.1046/j.1461-0248.2002.00373.x |
| [26] | DePina A S, Iser W B, Park S S, et al. Regulation of Caenorhabditis elegans vitellogenesis by DAF-2/ⅡS through separable transcriptional and posttranscriptional mechanisms[J]. BMC Physiology, 2011, 11. DOI:10.1186/1472-6793-11-11 |
| [27] | Huang Q S, Fang C, Wu X L, et al. Perfluorooctane sulfonate impairs the cardiac development of a marine medaka (Oryzias melastigma)[J]. Aquatic Toxicology, 2011, 105(1-2): 71-77. DOI:10.1016/j.aquatox.2011.05.012 |
| [28] | Pan Y, Yan S W, Li R Z, et al. Lethal/sublethal responses of Daphnia magna to acute norfloxacin contamination and changes in phytoplankton-zooplankton interactions induced by this antibiotic[J]. Scientific Reports, 2017, 7. DOI:10.1038/srep40385 |
| [29] | Bownik A, S'laska B, Bochra J, et al. Procaine penicillin alters swimming behaviour and physiological parameters of Daphnia magna[J]. Environmental Science and Pollution Research, 2019, 26(18): 18662-18673. DOI:10.1007/s11356-019-05255-2 |
| [30] | Imhof H K, Rusek J, Thiel M, et al. Do microplastic particles affect Daphnia magna at the morphological, life history and molecular level?[J]. PLoS One, 2017, 12(11). DOI:10.1371/journal.pone.0187590 |
| [31] | Meinertz J R, Schreier T M, Bernardy J A. Chronic toxicity of erythromycin thiocyanate to Daphnia magna in a flow-through, continuous exposure test system[J]. Bulletin of Environmental Contamination and Toxicology, 2011, 87(6): 621-625. DOI:10.1007/s00128-011-0415-8 |
| [32] | Kim H Y, Lee M J, Yu S H, et al. The individual and population effects of tetracycline on Daphnia magna in multigenerational exposure[J]. Ecotoxicology, 2012, 21(4): 993-1002. DOI:10.1007/s10646-012-0853-z |
| [33] | Hall A, Karplus P A, Poole L B. Typical 2-Cys peroxiredoxins-structures, mechanisms and functions[J]. The FEBS Journal, 2009, 276(9): 2469-2477. DOI:10.1111/j.1742-4658.2009.06985.x |
| [34] | Zhang Q F, Li Y W, Liu Z H, et al. Exposure to mercuric chloride induces developmental damage, oxidative stress and immunotoxicity in zebrafish embryos-larvae[J]. Aquatic Toxicology, 2016, 181: 76-85. DOI:10.1016/j.aquatox.2016.10.029 |
| [35] | Jia W, Mao L, Zhang L, et al. Effects of two strobilurins (azoxystrobin and picoxystrobin) on embryonic development and enzyme activities in juveniles and adult fish livers of zebrafish (Danio rerio)[J]. Chemosphere, 2018, 207: 573-580. DOI:10.1016/j.chemosphere.2018.05.138 |
| [36] |
李杜文, 华智杰, 张文香, 等. 金属蛋白酶对大菱鲆血清抗氧化酶活性及丙二醛含量的影响[J]. 海洋科学, 2019, 43(4): 52-60. Li D W, Hua Z J, Zhang W X, et al. Effects of metalloprotease on antioxidant system enzyme activities and malondialdehyde content in the serum of Scophthalmus maximus[J]. Marine Sciences, 2019, 43(4): 52-60. |
| [37] | Viarengo A, Canesi L, Garcia Martinez P, et al. Pro-oxidant processes and antioxidant defence systems in the tissues of the Antarctic scallop (Adamussium colbecki) compared with the Mediterranean scallop (Pecten jacobaeus)[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 1995, 111(1): 119-126. DOI:10.1016/0305-0491(94)00228-M |
| [38] |
胡文琴, 王恬, 孟庆利. 动物中活性氧的产生及清除机制[J]. 家畜生态, 2004, 25(3): 64-67. Hu W Q, Wang T, Meng Q L. Mechanisms of generating and scavenging reactive oxygen species in animals[J]. Ecology of Domestic Animal, 2004, 25(3): 64-67. DOI:10.3969/j.issn.1673-1182.2004.03.020 |
| [39] |
姜航, 丁剑楠, 黄叶菁, 等. 聚苯乙烯微塑料和罗红霉素对斜生栅藻(Scenedesmus obliquus)和大型溞(Daphnia magna)的联合效应研究[J]. 生态环境学报, 2019, 28(7): 1457-1465. Jiang H, Ding J N, Huang Y J, et al. Combined effects of polystyrene microplastics and roxithromycin on the green algae (Scenedesmus obliquus) and waterflea (Daphnia magna)[J]. Ecology and Environmental Sciences, 2019, 28(7): 1457-1465. |
| [40] | Sevanian A, Ursini F. Lipid peroxidation in membranes and low-density lipoproteins: similarities and differences[J]. Free Radical Biology and Medicine, 2000, 29(3-4): 306-311. DOI:10.1016/S0891-5849(00)00342-7 |
| [41] | Parolini M, Binelli A, Provini A. Chronic effects induced by ibuprofen on the freshwater bivalve Dreissena polymorpha[J]. Ecotoxicology and Environmental Safety, 2011, 74(6): 1586-1594. DOI:10.1016/j.ecoenv.2011.04.025 |
| [42] |
王丽, 张耘泽, 耿歌, 等. 高效氯氰菊酯急性暴露中斑马鱼相关酶活性和基因表达变化[J]. 生态毒理学报, 2016, 11(4): 146-154. Wang L, Zhang Y Z, Geng G, et al. Variations of relevant enzyme activities and genes expressions in zebrafish under acute exposure to beta cypermethrin[J]. Asian Journal of Ecotoxicology, 2016, 11(4): 146-154. |
| [43] | Daghrir R, Drogui P. Tetracycline antibiotics in the environment: a review[J]. Environmental Chemistry Letters, 2013, 11(3): 209-227. DOI:10.1007/s10311-013-0404-8 |
| [44] | Subramoniam T. Mechanisms and control of vitellogenesis in crustaceans[J]. Fisheries Science, 2011, 77(1): 1-21. DOI:10.1007/s12562-010-0301-z |
| [45] |
季旭, 黄权. 鱼类卵黄蛋白和卵黄蛋白原的研究进展[J]. 饲料工业, 2016, 37(4): 32-36. Ji X, Huang Q. Research progress of fish vitellin and vitellogenin[J]. Feed Industry, 2016, 37(4): 32-36. |
| [46] | Jones P D, De Coen W M, Tremblay L, et al. Vitellogenin as a biomarker for environmental estrogens[J]. Water Science & Technology, 2000, 42(7-8): 1-14. |
| [47] | Mahapatra S, Kabita S, Bhattacharya D, et al. Purification and development of ELISAs for two forms of vitellogenin in Indian walking catfish, Clarias batrachus (L.)[J]. Fish Physiology and Biochemistry, 2017, 43(2): 477-491. DOI:10.1007/s10695-016-0304-5 |
| [48] | Seyoum A, Pradhan A, Jass J, et al. Perfluorinated alkyl substances impede growth, reproduction, lipid metabolism and lifespan in Daphnia magna[J]. Science of the Total Environment, 2020, 737. DOI:10.1016/j.scitotenv.2020.139682 |
| [49] | Nagaraju G P C. Reproductive regulators in decapod crustaceans: an overview[J]. Journal of Experimental Biology, 2011, 214: 3-16. DOI:10.1242/jeb.047183 |
| [50] | Kato Y, Kobayashi K, Oda S, et al. Cloning and characterization of the ecdysone receptor and ultraspiracle protein from the water flea Daphnia magna[J]. Journal of Endocrinology, 2007, 193(1): 183-194. DOI:10.1677/JOE-06-0228 |
| [51] | In S, Yoon H W, Yoo J W, et al. Acute toxicity of bisphenol A and its structural analogues and transcriptional modulation of the ecdysone-mediated pathway in the brackish water flea Diaphanosoma celebensis[J]. Ecotoxicology and Environmental Safety, 2019, 179: 310-317. DOI:10.1016/j.ecoenv.2019.04.065 |
| [52] | Laflamme L, Hamann G, Messier N, et al. RXR acts as a coregulator in the regulation of genes of the hypothalamo-pituitary axis by thyroid hormone receptors[J]. Journal of Molecular Endocrinology, 2002, 29(1): 61-72. DOI:10.1677/jme.0.0290061 |
| [53] | Sumiya E, Ogino Y, Miyakawa H, et al. Roles of ecdysteroids for progression of reproductive cycle in the fresh water crustacean Daphnia magna[J]. Frontiers in Zoology, 2014, 11. DOI:10.1186/s12983-014-0060-2 |
| [54] | Street S M, Eytcheson S A, LeBlanc G A. The role of nuclear receptor E75 in regulating the molt cycle of Daphnia magna and consequences of its disruption[J]. PLoS One, 2019, 14(8). DOI:10.1371/journal.pone.0221642 |
| [55] | Carriero M M, Henrique-Silva F, Caetano A R, et al. Characterization and gene expression analysis of pacu (Piaractus mesopotamicus) inducible nitric oxide synthase (iNOS) following Aeromonas dhakensis infection[J]. Fish & Shellfish Immunology, 2018, 74: 94-100. |
| [56] | Cabrié A, Guittet O, Tomasini R, et al. Crosstalk between TAp73 and TGF-β in fibroblast regulates iNOS expression and Nrf2-dependent gene transcription[J]. Free Radical Biology and Medicine, 2019, 134: 617-629. DOI:10.1016/j.freeradbiomed.2019.02.008 |
| [57] | Mishra O P, Mishra R, Ashraf Q M, et al. Nitric oxide-mediated mechanism of neuronal nitric oxide synthase and inducible nitric oxide synthase expression during hypoxia in the cerebral cortex of newborn piglets[J]. Neuroscience, 2006, 140(3): 857-863. DOI:10.1016/j.neuroscience.2006.02.060 |
| [58] | Hellou J, Ross N W, Moon T W. Glutathione, glutathione S-transferase, and glutathione conjugates, complementary markers of oxidative stress in aquatic biota[J]. Environmental Science and Pollution Research, 2012, 19(6): 2007-2023. DOI:10.1007/s11356-012-0909-x |
| [59] | Xia X C, Yu R X, Li M B, et al. Molecular cloning and characterization of two genes encoding peroxiredoxins from freshwater bivalve Anodonta woodiana: Antioxidative effect and immune defense[J]. Fish & Shellfish Immunology, 2018, 82: 476-491. |
| [60] | Liu G, Feinstein S I, Wang Y, et al. Comparison of glutathione peroxidase 1 and peroxiredoxin 6 in protection against oxidative stress in the mouse lung[J]. Free Radical Biology and Medicine, 2010, 49(7): 1172-1181. DOI:10.1016/j.freeradbiomed.2010.07.002 |