 2021, Vol. 42
2021, Vol. 42



2. 苏州科技大学城市生活污水资源化利用技术国家地方联合工程实验室, 苏州 215009
2. National and Local Joint Engineering Laboratory of Municipal Sewage Resource Utilization Technology, Suzhou University of Science and Technology, Suzhou 215009, China
ANAMMOX细菌(AnAOB)具备的代谢底物多样性引起了广泛关注, 除了以氨(NH4+)和亚硝酸盐(NO2-)为底物, AnAOB还可以利用具有氧化性的Fe3+[1]和Mn4+[2]等为电子受体氧化NH4+或利用还原性物质Fe0[3]、Fe2+[4]和小分子有机酸[5]等为电子供体还原NO2-/NO3-.在ANAMMOX污泥反应器中还发现了NH4+与SO42-同步转化现象, 有研究者认为这是由AnAOB完成的生物转化过程, 并将其命名为硫酸盐型厌氧氨氧化(sulfate reduction ammonium oxidation, SRAO), 即: 硫酸盐的生物还原与氨的生物氧化相耦合, 最终通过产生N2实现生物脱氮的生物反应系统.
国内外数十位学者在自然环境和实验室反应器中相继对ANAMMOX微生物进行SRAO生物转化进行研究.美国罗德岛大学的Schrum等[6]在进行海底沉积物间隙水的生物地球化学通量与吉布斯能的原位测定时, 发现在缺氧和好氧交界处存在明显的氮和硫损失, 并将其归因于SRAO过程.随后, 丹麦的Canfield等[7]报道在智利海岸“低氧带”, 发现了不同寻常的硫酸盐还原和硫化物氧化, 推测该过程与包括ANAMMOX在内的其它氮循环过程密切关联.2001年, Fdz-Polanco等[8]最先在处理甜菜糖蜜废水的厌氧生物反应器中发现了“不寻常”氮和硫损失, 提出了在厌氧条件下SRAO反应的方程式为:
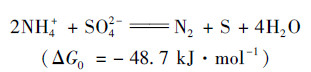
|
(1) |
从热力学的角度, 式(1)的ΔG0为-48.7 kJ·mol-1, 故该反应是可能自发进行的.作者进一步提出该反应是由3个分步反应组成, 氨与硫酸盐转化的中间产物分别为NO2-和S2-:
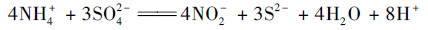
|
(2) |
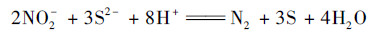
|
(3) |
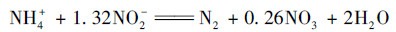
|
(4) |
然而, 在随后的研究中, 该研究团队并未能重现上述实验现象[9].随后, 董凌霄等[10]和Sabumon等[11~13]也在含硫酸盐和氨的有机废水中发现了这一现象, 且检测到有S2-生成; Liu等[14]和Yang等[15]分别在无机营养条件下观察到了ANAMMOX培养物中的SRAO生物转化现象, 检测到氨与硫酸盐转化后的产物为N2和单质硫, 并以“Anammoxoglobus sulfate”命名所用的ANAMMOX培养物, 基于此提出SRAO的反应方程式应为:
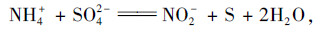
|
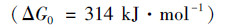
|
(5) |
综合上述研究结果, 不少研究者认为SRAO现象是由ANAMMOX微生物完成的生物转化过程.课题组前期对ANAMMOX培养物中的SRAO现象进行了数年研究, 得到了部分与上述报道相似的结果, 即:以硫酸盐和氨作为主要底物的ANAMMOX培养物中, SRAO现象是客观存在的[16~20].但同时发现, 已有文献对ANAMMOX培养物中SRAO现象的基质转化途径和反应机制的解释仍然存在不合理之处, 不同研究所得出的结论也并不一致.集中体现在以下问题:其一, NH4+与SO42-的摩尔转化比(N/S)远远大于2, 而依据Fdz-Polano等[8]假设提出的反应方程式[式(1)], N/S比应为2.事实上, 在其他文献中N/S比也存在明显差异, 其数值在2~20范围内不等, 这并不符合特定生物转化反应中反应物摩尔比为定值的特征, 也暗示了分步反应[式(2)]可能是由几个平行反应构成的, 亦或是在反应中存在着硫酸盐还原和还原产物被重新氧化成为硫酸盐的循环.其二, NH4+与SO42-转化的中间产物不明确:①依照方程[式(1)], NH4+氧化的中间产物为NO2-, 但相关研究中只有Liu等[14]检测到浓度极低的NO2-, 而其他研究者均并未检测到NO2-生成.②在无机营养条件下观察到SRAO现象时, 未检测到S2-, 也几乎检测不到NO2-.对比其他文献可以看出, 只有在有机营养条件下进行的研究观察到S2-产生, 而在无机营养条件下进行的研究均未检测到S2-, 上述现象也并不符合某一特定生物转化反应其转化产物确定性的特征.
依据课题组近期的研究结果[19, 20], SRAO很可能是NH4+与SO42-盐各自独立转化的结果, 该现象的发生取决于环境中的溶解氧、有机物等条件, 可能的转化途径为:ANAMMOX培养物中存在的氨氧化细菌(aerobic ammonium oxidation bacteria, AOB)利用环境中的微量溶解氧(dissolved oxygen, DO)将氨氧化; 同时, 异养硫酸盐还原菌(sulfate reduction bacteria, SRB)利用污泥微生物衰亡释放出的有机物将SO42-还原.这一观点可以解释实验中N/S比差异、氨与硫酸盐转化不同步的现象, 但仍缺少相关证据以明确SRAO反应中氨与硫酸盐转化途径以及产ANAMMOX微生物在其中所起的作用.为了解决上述问题, 在不同DO、氧化还原电位(ORP)以及不同微生物组成的条件下进行批次实验和连续流实验, 借助16S rDNA高通量测序和功能基因表达定量分析获取分子生物学信息, 以更清晰地解析ANAMMOX培养物中氨与硫酸盐同步转化现象背后的反应机制.本研究主要回答以下3个关键问题:①SRAO现象发生的条件; ②SRAO生物转化过程是否由AnAOB完成; ③ ANAMMOX培养物中氨与硫酸盐的转化途径.
1 材料与方法 1.1 实验装置、用水与接种污泥连续流反应器装置(continuous flow stirred tank reactor, CFSTR)采用厌氧发酵罐(No. Labfors 5 Bacteria, Switzerland)[19, 20], 设置水力停留时间为10 h.批次实验(Batch)使用总体积120 mL的厌氧血清瓶, 将其固定在转速为150 r·min-1的恒温空气浴振荡器内.实验用水主要含60 mg·L-1NH4+-N(以NH4Cl形式添加)和90 mg·L-1SO42--S(以Na2SO4形式添加).其他主要成分包括:125 mg·L-1 KHCO3、54 mg·L-1 KH2PO4和大量元素(包括CaCl2·2H2O、NaCl、MgSO4·7H2O、KCl)[21].所有实验用水均以纯度为99.5%的氮气曝气15~30 min, 使DO浓度低于0.5 mg·L-1, pH调节为7.0±0.2.
所用污泥有两种, 分别为:ANAMMOX污泥1 L, 取自本实验室稳定运行的亚硝酸盐型ANAMMOX反应器, 氮负荷约2 kg·(m3·d)-1. 活性污泥1 L, 取自苏州市某城市污水厂的污泥浓缩池.污泥预处理方法如下:上述两种污泥先分别用0.9% NaCl清洗数次, 12 000 r·min-1离心15 min后撇去上清液; 再分别称取100 g上述污泥重新悬于1 L的0.9% NaCl溶液, 待用.下文所述的ANAMMOX污泥即直接量取预处理后的ANAMMOX培养物(600 mL), 混合污泥由预处理后的ANAMMOX污泥与活性污泥各300 mL混合得到.
1.2 实验方案本实验分为ANAMMOX组与混合污泥组, 方案设计如图 1所示.首先, 在两个相同的发酵罐中, 分别装入上述ANAMMOX污泥(CSTRAn)和混合污泥(CSTRMix), 进行连续流实验, 反应器运行期均分别为30 d和40 d.待连续流实验结束, 从每个发酵罐中取出相同体积的污泥, 分别转移至厌氧血清瓶中进行两组批次实验, 分别记为BatchAn和BatchMix; 每组又依据ORP不同, 分别记为BatchAn-1、BatchAn-2和BatchMix-1和BatchMix-2.本研究中, 不同的厌氧条件通过调控氮气(纯度99.5%)的曝气时长, 并以ORP和DO为指标进行表征.具体实验条件如表 1所述.

|
图 1 实验设计流程 Fig. 1 Schematic diagram of the up-flow bioreactor |
|
|
表 1 连续流反应器和批次实验条件 Table 1 Operating conditions of the CSTR and batch tests |
1.3 测定指标和方法
|
|
表 2 测定项目与方法 Table 2 Measurement items and methods |
1.4 16S rDNA高通量测序和功能基因实时荧光定量PCR分析
提取使用Omega基因组DNA抽提试剂盒Mag-Bind Soil DNA Kit抽提微生物样品DNA, 并进行定量分析(Qubit3.0 DNA试剂盒).PCR扩增采用细菌16S V3-V4可变区通用引物[23], 每个PCR产物有3个重复, 将重复的PCR产物等量混合后用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司, 美国) 纯化并切胶回收.用QuantiFluorTM-ST蓝色荧光定量系统(Promega, USA)定量检测PCR产物, 最后, 得到的PCR产物采用Illumina MiSeq测序仪进行PE300测序(Majorbio Co.; Ltd., Shanghai, China).
对污泥样品的总细菌16S rDNA[2]、氨氧化酶标记基因amoA[24]、硝酸盐异化还原酶标记基因narG[25]、联氨合成酶标记基因hzsB[26]和硫酸盐还原酶标记基因apsA[27]进行SYBRⅡ法荧光定量PCR(qPCR)分析.
2 结果与分析 2.1 混合污泥体系中氨与硫酸盐转化在接种混合污泥的CSTRMix反应器中, 依据ORP值不同, 运行分为Ⅰ、Ⅱ、Ⅲ和Ⅳ阶段, 图 2为CSTRMix运行中NH4+、SO42-、TOC、S2-浓度及体系内ORP、pH变化.阶段Ⅰ(1~10 d)期间ORP值稳定在约(-100±10) mV, NH4+-N平均转化量为16.1 mg·L-1, 但SO42-未发生转化且出水浓度高于进水浓度.阶段Ⅱ(11~20 d)期间, ORP值维持在(-200±10) mV水平, 反应器中观察到了NH4+与SO42-浓度同时降低的现象, 但二者转化量变化趋势并不一致, NH4+-N转化量由8.2 mg·L-1提高至20.7 mg·L-1, SO42--S转化量由22.4 mg·L-1降低至8.7 mg·L-1, 此时体系中TOC浓度较低, 且未检测到S2-.同时, 在这一阶段进、出水的pH差值增大.在阶段Ⅲ(21~30 d), CSTRMix内ORP平均值为(-490±15) mV, 此时NH4+几乎不转化, SO42-平均转化量为14.79 mg·L-1, TOC和S2- 平均浓度为29.0 mg·L-1和8.8 mg·L-1.阶段Ⅳ(31~40 d), ORP恢复至(-200±10) mV水平, NH4+与SO42-浓度同时降低的现象再次出现, NH4+-N与SO42--S平均转化量分别15.55 mg·L-1和6.96 mg·L-1, TOC浓度较低但仍未检测到S2-, 这与阶段Ⅱ内的反应现象类似.

|
图 2 不同ORP条件下CSTRMix反应器中氨与硫酸盐的转化 Fig. 2 Conversion of ammonium and sulfate in CSTRMixunder different ORP conditions |
为了进一步明确ORP对NH4+与SO42-的同步转化现象的影响, 取上述CSTRMix反应器中的部分污泥在ORP分别为-100 mV(BatchMix-1)和-480 mV(BatchMix-2)条件下进行10 d的批次实验, 结果如图 3所示.在BatchMix-1中, NH4+浓度在前4 d由55.4 mg·L-1降至约25.7 mg·L-1, 但随后上升, 第10 d时氨的最终浓度为45.5 mg·L-1.在BatchMix-2中, ORP值远远低于BatchMix-1, 期间NH4+没有转化, 且在第2 d之后浓度持续升高.然而, 硫酸盐转化速率明显高于BatchMix-1, 在第6 d其浓度仅为2.3 mg·L-1.由图 3可以看出, 在第6 d之后, NH4+与SO42-转化速率相当缓慢, 故选取前6 d实验数据计算ORP分别为-100 mV(BatchMix-1)和-480 mV(BatchMix-2)时, 氨的转化速率(以VSS计)分别为0.12 mg·(g·d)-1和-0.09 mg·(g·d)-1; 硫酸盐转化速率(以VSS计)分别为0.55 mg·(g·d)-1和0.78 mg·(g·d)-1.

|
图 3 不同ORP条件下BatchMix中氨与硫酸盐的转化 Fig. 3 Conversion of ammonium and sulfate in BatchMix under different ORP conditions |
与CSTRMix反应器运行类似, 在接种ANAMMOX污泥的CSTRAn反应器中同样以NH4+和SO42-为基质, 连续运行30 d, 在3个运行阶段内ORP分别控制在(-100±15)、(-220±10)和(-477±10)mV水平.由图 4可以看出, 在ANAMMOX污泥体系中, NH4+转化量与ORP值密切相关.在阶段Ⅰ(1~10 d), 氨转化量最大, 平均转化速率(以VSS计)为0.90 mg·(g·d)-1; 进入阶段Ⅱ(11~20 d), 随着ORP降至-200 mV以下, NH4+平均转化速率明显降低; 阶段Ⅲ(21~30 d)ORP降至-477 mV时, NH4+停止转化.在上述3个阶段的运行期间, 进、出水中SO42-浓度几乎无差别, 也未检测到S2-, 表明SO42-始终未转化.此外, 出水中也未检测到TOC.

|
图 4 不同ORP条件下CSTRAn反应器中氨与硫酸盐的转化 Fig. 4 Conversion of ammonium and sulfate in CSTRAn under different ORP conditions |
鉴于CSTRMix实验结果, 较低的ORP值可能更有利于SO42-的转化, 因此取上述CSTRAn反应器中的部分ANAMMOX污泥在ORP分别为-100 mV(BatchAn-1)和-480 mV(BatchAn-2)条件下进行10 d的批次实验.由图 5(a)可以看出, 当ORP处于极低的-480 mV水平时, NH4+停止转化, 其浓度无变化, 这一现象与BatchMix-2中所观察到表现一致; SO42-仍无明显转化, 在实验后期浓度有所上升.图 5(b)显示了2组实验中, ORP和TOC随时间的变化情况.随着反应时间的延长, BatchAn-1中ORP由-100 mV快速下降至-350 mV以下, 而BatchAn-2中的ORP值变化不大.同时, 2组实验中TOC浓度均有不同程度地升高, 相比之前, ORP在-480 mV水平下的BatchAn-2中TOC增幅更大.

|
图 5 不同ORP条件下, BatchAn中氨与硫酸盐的转化以及TOC浓度变化 Fig. 5 Conversion of ammonium and sulfate in BatchAn under different ORP conditions |
为了进一步探究氨与硫酸盐同步转化过程中的主要功能微生物及其丰度变化, 取接种时的ANAMMOX污泥(SAn-1)、混合污泥(SMix-1)和连续流实验后的ANAMMOX污泥(SAn-2)、混合污泥(SMix-2)样品进行16S rDNA高通量测序, 测序结果如表 3所示.
|
|
表 3 α多样性指数 Table 3 Indexes of α diversity |
各样品的有效序列数分别为77 707、80 598、70 996和75 033, 有效操作单元(OTUs)数在4 666~5 694之间, 覆盖度(coverage)大于94%, 以Shannon指数为纵坐标的稀释曲线图说明各个样品的测序数据量合理.Chao1、Shannon和Simpson指数描述了反应器运行期间ANAMMOX污泥和混合污泥样品中微生物群落结构的演变.以Shannon指数为例, 可以看出SAn-1较SAn-2该指数升高, 而SMix-1较SMix-2该指数下降.这表明以NH4+和SO42-为基质时, ANAMMOX污泥的微生物群落多样性增加, 物种总数降低, 而混合污泥的微生物群落多样性下降, 物种总数增多.
在门分类水平上(图 6), 4个样品中共检测到37种细菌及古菌, 其中相对丰度大于1%的有9个门, 包括:Chloroflexi(绿弯菌门)、Proteobacteria(变形菌门)、Planctomycetes(浮霉菌门)和Bacteroidetes(拟杆菌门)等.在混合污泥中, ANAMMOX菌所属的Planctomycetes丰度分别由26.1%(SMix-1)增加至33.9%(SMix-2); 相反地, 在ANAMMOX污泥中, ANAMMOX菌丰度由34.1%(SAn-1)略提高至35.7%(SAn-2).Proteobacteria微生物在混合污泥中的丰度略有上升, 但其在ANAMMOX污泥中的丰度由34.1%降至17.5%, 变化更为显著.拟杆菌门(Bacteroidetes)始终是混合污泥中丰度最高的一类微生物, 相对丰度约43.1%.然而, 此类微生物在ANAMMOX污泥中的丰度由23.8%(SAn-2)下降至21.5%(SAn-1).此外, 其他主要微生物中, Firmicutes在两种污泥中的丰度均有所增加; Acidobacteria以及Chloroflexi的相对丰度在混合污泥中有均所增加, 但在ANAMMOX污泥中减少.

|
图 6 各污泥样品门水平微生物群落Heatmap图 Fig. 6 Heatmap analysis of mixed sludge and ANAMMOX sludge at the phyla level |
在属水平上丰度前20的物种如图 7所示.在混合污泥样品SMix-1和SMix-2中, 可以看出以AnAOB Candidatus Brocadia (Ca. Brocadia)相对丰度由20.2%增大到25.5%, 增幅显著; 在ANAMMOX污泥中其相对丰度由初始的32.1%提高至34.6%, 增幅较小.相比之下, 好氧硝化菌Nitrospira(硝化螺菌属)以及与硫酸盐还原相关的Thiobacillus(硫杆菌属)、Sulfurisoma(硫体属)和Desulfomicrobium(脱硫化微菌属)等微生物的丰度均明显提高.而在ANAMMOX污泥样品SAn-1和SAn-2中, AnAOB(Ca. Brocadia)丰度变化较小, Thiobacillus属、Nitrosomonas(亚硝化单胞菌属)和Desulfovibrio(脱硫弧菌属)等丰度明显降低.

|
红色字体为重点关注物种, 下同 图 7 各污泥样品属水平微生物丰度比例差异 Fig. 7 Abundance of ANAMMOX sludge and mixed sludge at the gene level |
为了明确ANAMMOX污泥和混合污泥中NH4+氧化和SO42-盐还原的生物转化途径, 选取了好氧氨氧化标记基因amoA、异养硫酸盐还原标记基因apsA、异养硝酸盐还原标记基因narG以及ANAMMOX标记基因hzsB的丰度变化进行定量qPCR分析.
(1) amoA amoA(氨单加氧酶)是亚硝化过程的关键酶编码基因, 被广泛应用于AOB菌的研究[28].图 8所示为混合污泥样品(SMix-1、SMix-2)和ANAMMOX污泥样品(SAn-1、SAn-2)中携带amoA基因的微生物(属水平)和其中amoA基因的相对丰度, 以及相应丰度值在95%置信区间内的差异显著性(P < 0.05).

|
图 8 混合污泥(SMix-1和SMix-2)与ANAMMOX污泥(SAn-1与SAn-2)中携带amoA基因的微生物(属水平)丰度变化 Fig. 8 Abundance of amoA gene expressed microorganisms at the genera level in mixed sludge (SMix-1/SMix-2) and ANAMMOX sludge (SAn-1/SAn-2) |
从图 8中可以看出, 在混合污泥样品以及ANAMMOX污泥样品中, 携带amoA基因的微生物类别并无差异, 其中绝大部分微生物为Nitrosomonas, 其余少量为Nitrosospira.在混合污泥SMix-2与SMix-1中, Nitrosomonas丰度随着实验进行明显增加、Nitrosospira丰度下降; 而对比ANAMMOX污泥SAn-2与SAn-1, Nitrosomonas和Nitrosospira的相对丰度均随反应进行有所提高.
(2) apsA apsA是腺苷酰硫酸还原酶的功能基因, 常用于鉴别SRB的存在[29].图 9是混合污泥(SMix-1和SMix-2)和ANAMMOX污泥(SAn-1和SAn-2)中携带apsA基因的微生物在属水平分布情况及其相对丰度.对比SMix-1和SMix-2样品可以看出, Desulfomicrobium、Desulfovibrio和Desulfonatronum(脱硫化泡碱菌属)丰度增幅约为1%~3%, 这与CSTRMix反应器中逐渐增强的硫酸盐转化现象一致.其中, Desulfovibrio属和Desulfomicrobium属主要参与硫化合物还原, 而Desulfonatronum属主要参与硫及硫化物的氧化过程[29~31].在SAn-1和SAn-2中, Desulfovibrio的丰度比例降幅约为90%, 这一变化可能由于CSTRAn反应器中ORP值始终在(-100±15)mV水平, 高于硫酸盐还原菌所适宜的ORP值环境(< -100 mV)导致, 这与CSTRAn、BatchAn中的硫酸盐无转化现象一致.

|
图 9 混合污泥(SMix-1和SMix-2)与ANAMMOX污泥(SAn-1与SAn-2)中携带apsA基因的微生物(属水平)丰度变化 Fig. 9 Abundance of apsA gene expressed microorganisms at the genera level in mixed sludge (SMix-1/SMix-2) and ANAMMOX sludge (SAn-1/SAn-2) |
(3) narG narG是编码Nar(nitrate reductase, 硝酸盐还原酶)的功能基因, 图 10是混合污泥(SMix-1和SMix-2)和ANAMMOX污泥(SAn-1和SAn-2)中携带narG基因的微生.在SMix-1和SMix-2中携带narG基因的微生物为常见的反硝化细菌, 包括:Denitronmonas、Haloferax、Sterolibactterium和Stenotrophomonas, 除去Stenotrophomonas为好养反硝化菌[32], 其他均为厌氧反硝化菌[33].其中, 只有Stenotrophomonas丰度下降, 其余narG基因携带微生物的丰度均增加, 原因在于CSTRMix中NO3-浓度始终低于检测限, 且实验过程中反应器内DO和ORP均较低, 不利于好氧反硝化菌的生长.

|
图 10 混合污泥(SMix-1和SMix-2)与ANAMMOX污泥(SAn-1与SAn-2)中携带narG基因的微生物(属水平)丰度变化 Fig. 10 Abundance of narG gene expressed microorganisms at the genera level in mixed sludge (SMix-1/SMix-2) and ANAMMOX sludge (SAn-1/SAn-2) |
(4) hzsB hzsB基因编码联氨合成酶, 常用作AnAOB的标记基因.图 11是混合污泥(SMix-1和SMix-2)和ANAMMOX污泥(SAn-1和SAn-2)中携带hzsB基因的菌属及其中hzsB基因的丰度.可以看出随着反应的持续, Candidatus Jettenia(Ca. Jettenia)、Azoarcus、Sphingomonas和Ca. Broocadia所占丰度均略有增加, 因此体系中存在ANAMMOX反应.但由于体系内NH4+和NO2-始终浓度较低, 因此AnAOB处于营养贫乏的环境中, 其相对丰度没有明显提高.

|
图 11 混合污泥(SMix-1和SMix-2)与ANAMMOX污泥(SAn-1与SAn-2)中携带hzsB基因的微生物(属水平)丰度变化 Fig. 11 Abundance of hzsB gene expressed microorganisms at the genera level in mixed sludge (SMix-1/SMix-2) and ANAMMOX sludge (SAn-1/SAn-2) |
本研究通过连续流实验和批次实验, 在不同厌氧状态下(微氧:-100 mV < ORP < 0 mV、0.5 mg·L-1 < DO < 1 mg·L-1; 缺氧:-300 mV < ORP < -100 mV、0.2 mg·L-1 < DO < 0.5 mg·L-1; 厌氧:ORP < -300 mV、DO < 0.2 mg·L-1), 分别接种ANAMMOX污泥与混合污泥对SRAO现象的发生条件进行识别, 并结合16S rDNA高通量测序与相关功能基因表达解析上述实验条件下氨与硫酸盐的转化途径.结果显示, 氨与硫酸盐可以各自独立转化, 只有在某些特定条件下SRAO现象才会发生, 这主要取决于与厌氧条件和微生物群落组成.
3.1 氨与硫酸盐同步转化发生的条件CSTRAn实验的接种污泥为ANAMMOX污泥, 在微氧和缺氧条件下, 无论是连续流实验(ORP平均分别为-100 mV和-200 mV)还是批次实验(ORP平均为-100 mV), 均可以观察到氨氧化的现象, 且微氧条件下氨转化速率更快.而在厌氧条件下, 无论是连续流实验或批次实验, 氨都不会转化.上述所有使用ANAMMOX污泥的实验中均未观察到硫酸盐的转化.在接种混合污泥的CSTRMix实验中, 氨与硫酸盐的转化同样与厌氧状态密切相关.微氧条件下, 氨被转化但硫酸盐不转化; 而当ORP降至约-490 mV达到厌氧条件时, 硫酸盐转化加快, 氨停止转化.实验过程中发现, 只有在缺氧状态下, 氨与硫酸盐同步转化的现象发生, 但相比微氧条件时, 氨的转换速率减缓.在接种混合污泥的批次实验过程中, 也观察到相同的实验规律.显然, 氨与硫酸盐是各自独立转化的, 并非是由同一个生物反应过程完成的; 而二者的同步转化现象只有在特定条件下(混合污泥和缺氧条件)才发生, 这与本课题组前期的研究结果吻合[18~20].
3.2 氨转化条件与途径通过分析ANAMMOX污泥(SAn-1和SAn-2)中氨氧化功能基因amoA表达变化以及微生物群落结构, 可以发现:混合污泥和ANAMMOX污泥中均存在携带amoA功能基因的氨氧化细菌(Nitrosomonas属和Nitrosospira属), 且amoA基因的表达在连续流实验CSTRAn与CSTRMix中被强化(图 8); 微生物群落结构分析也证实上述微生物的相对丰度有所增加(图 7).上述实验结果表明:在混合污泥和ANAMMOX污泥中发生了氨氧化反应, 以Nitrosomonas和Nitrosospira为主的氨氧化细菌是完成这一过程的主要功能微生物.显然, 氨氧化过程受到厌氧条件的直接影响:厌氧程度越高(ORP和DO越低)、氨转化速率越慢, 微氧条件下的氨转化速率明显高于缺氧条件下的氨转化速率, 而厌氧条件下氨无法被转化.由于在实验过程中, 进水未添加硝酸盐、亚硝酸盐等氧化态化合物, 故氨氧化的电子受体应为氧气, 而不是硫酸盐.正是依赖于氨氧化细菌的作用, 氨被氧化为亚硝酸盐, 为ANAMMOX细菌的生长代谢提供了必要的底物.微生物群落结构分析也可为这一结论提供直接证据.在图 7中可以看出, 相比接种初期, 活性污泥与ANAMMOX污泥中ANAMMOX细菌的相对丰度均有所增加.经氨氧化反应[式(6)]与ANAMMOX反应[式(7)], 氨最终被转化为氮气和少量硝酸盐, 然而在实现过程中硝酸盐浓度始终较低, 相关讨论在3.3部分展开.
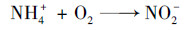
|
(6) |
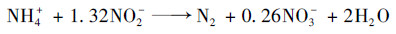
|
(7) |
好氧氨氧化的发生需同时满足两个条件:AOB和氧.在SRAO相关研究中, 不同研究者所使用的接种污泥不同, 包括活性污泥[29]、厌氧污泥[8, 9, 34~36]、以及ANAMMOX污泥[14], 其中大部分研究都发现微生物群落中存在AOB, 这类好氧或兼性好氧微生物可以保护AnAOB免受氧气的抑制作用.此外, ANAMMOX微生物群落中的Armatimonadetes、Acidobacteria、Verrucomicrobia和Nitrosomonas 也可以在厌氧条件下氧化氨[37].
氧是氨氧化过程的电子受体.在课题组前期研究中发现, 在连续流反应器中, 需依靠蠕动泵完成进、出水操作, 搅拌桨与容器间的密封性等问题都会导致氧气进入反应器系统.在这种情况下, 即使通过常规的厌氧实验操作(如采用惰性气体对进水曝气除氧和出水管水封等)也难以维持反应器始终处于严格厌氧条件, 实际上往往是缺氧或微氧状态.在相关研究中, SRAO现象也是在非严格厌氧条件下(ORP值在-300~-150 mV)被观察到的[18, 19].相比之下, 厌氧血清瓶的密封性更优, 因此可以为批次实验提供更为严格的厌氧反应条件.在课题组前期研究中, 就通过控制血清瓶中是否被液体充满来实现厌氧(充满)或缺氧(不充满)条件, 并推测缺氧环境是SRAO现象发生的必要条件之一, 而厌氧条件下SRAO现象不会发生.在此基础上, 本研究通过控制氮气的曝气时长, 以ORP和DO为依据, 实现更精确地调节反应系统的厌氧状态.本实验结果证实, 只有在缺氧条件下, SRAO现象才有可能发生.
除了上述讨论的溶解氧之外, 可能造成氨被氧化的其他氧化态物质包括:羟基自由基(·OH)、Fe3+(进水中的Fe2+被氧化)和NO3-(ANAMMOX反应副产物), 这些氧化性物质的存在也会影响体系内的氧化还原电位(即ORP值).有研究者认为来自于微生物衰亡自由基或者活性氧也可以作为厌氧状态下氨氧化的电子供体[11~13].也有部分研究认为在缺乏电子供体的环境中, 兼性异养微生物可以通过产生H2O2、进而释放O2来实现氧化氨的目的.这部分氧同样可被AOB利用易产生亚硝酸盐, 为AnAOB提供必要的代谢基质[37].
3.3 硫酸盐转化条件与途径通过连续流实验和批次实验结果不难看出, 无论是连续流实验还是批次实验, 混合污泥在厌氧和缺氧条件下均可观察到硫酸盐还原现象, 而ANAMMOX污泥在不同厌氧条件下均无法将硫酸盐还原, 即硫酸盐还原现象的发生需同时满足两个条件:SRB和溶解性有机物(作为硫酸盐还原的电子供体).联合16S rDNA高通量测序(图 7)以及异养硫酸盐还原功能基因apsA表达分析(图 9)的结果, 混合污泥样中存在多种携带apsA基因的微生物, 包括:Desulfomicrobium、Desulfovibrio以及Desulfonatronum, 且上述微生物在反应器运行期间的相对丰度均有所增加, 这些微生物主要涉及硫化合物还原和氧化过程[29~31].因此, 混合污泥中硫酸盐的转化是由SRB完成的.相比之下, ANAMMOX污泥中, Desulfovibrio是唯一携带apsA基因的SRB, 且随着反应进程丰度比例大幅降低, 导致这一结果的主要原因在于在ANAMMOX污泥中缺乏溶解性有机物, 故异养微生物Desulfovibrio因底物匮乏逐渐被淘汰.同时, 由于SRB是专性厌氧微生物, 其生存环境的ORP值往往低于-100 mV[31, 32], 故异养硫酸盐还原这一过程也受限于厌氧状态, 这与本研究观察到的实验现象相吻合.由图 2和图 3可以看出, 厌氧程度越高(ORP和DO越低)有利于硫酸盐还原, 厌氧条件下硫酸盐还原速率明显高于缺氧条件下硫酸盐还原速率, 而微氧条件下硫酸盐几乎难以被还原, 符合异养硫酸盐反应的客观规律.
异养硫酸盐还原发生的另一个必要条件是环境中存在溶解性有机物作为电子供体.由于本研究所有实验进水中均不含有机物, 故其来源有两方面:①实验过程中微生物衰亡导致溶解性有机物释放到环境中; ②在混合污泥系统中, 活性污泥的加入也会带入部分溶解性有机物.但是, 在实验过程中, 混合污泥体系内可被检测到的TOC浓度非常低, 原因是:相对硫酸盐而言, TOC含量较低, 因而成为浓度限制因子, 在SRB的作用下会被迅速消耗.综上, 硫酸盐转化是在SRB的作用下通过异养硫酸盐还原途径实现的, 其反应方程式如下[35]:
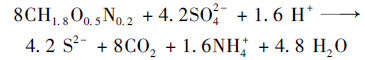
|
(8) |
在观察到SRAO现象的实验中, 异养硫酸盐还原的产物为S2-以及ANAMMOX反应副产物NO3-都始终维持很低浓度水平甚至未检测到.这一现象说明体系内除了好氧氨氧化、ANAMMOX、异养硫酸盐还原之外, 还有其他生物过程发生, 可能性最大的是异养反硝化和硫自养反硝化过程.①异养反硝化:在3.3节中已经讨论了体系内溶解性有机物的来源, 这部分有机物除了为异养硫酸盐还原提供电子供体, 同时也会为异养反硝化过程提供电子供体, 从而将ANAMMOX反应产生的硝酸盐还原为氮气[式(9)].②硫自养反硝化:该生物反应可以利用硝酸盐或亚硝酸盐为电子受体将还原态的S2-或S0氧化成SO42-[式(10)][34, 38], 因此推测这是本研究观察到SRAO现象时并未检测到S2-的原因.这一推论在其他研究中也可以得到证实[34], 上述研究者认为硫自养反硝化的作用是SRAO现象中硫酸盐转化总是不明显的原因.
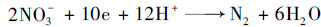
|
(9) |

|
(10) |
(1) SRAO现象发生的必要条件为:存在SRB和缺氧条件(-300 mV < ORP < -100 mV、0.2 mg·L-1 < DO < 0.5 mg·L-1)下才能发生;
(2) 没有证据表明AnAOB可以利用硫酸盐为电子供体氧化氨;
(3) SRAO现象实际上是好氧氨氧化、ANAMMOX和异养硫酸盐还原过程的联合作用的结果, 上述生物转化过程分别由AOB、AnAOB和SRB完成.
| [1] | Sawayama S. Possibility of anoxic ferric ammonium oxidation[J]. Journal of Bioscience and Bioengineering, 2006, 101(1): 70-72. DOI:10.1263/jbb.101.70 |
| [2] | Javanaud C, Michotey V, Guasco S, et al. Anaerobic ammonium oxidation mediated by Mn-oxides: from sediment to strain level[J]. Research in Microbiology, 2011, 162(9): 848-857. DOI:10.1016/j.resmic.2011.01.011 |
| [3] | Suzuki T, Moribe M, Oyama Y, et al. Mechanism of nitrate reduction by zero-valent iron: equilibrium and kinetics studies[J]. Chemical Engineering Journal, 2012, 183: 271-277. DOI:10.1016/j.cej.2011.12.074 |
| [4] | Oshiki M, Ishii S, Yoshida K, et al. Nitrate-dependent ferrous iron oxidation by anaerobic ammonium oxidation (anammox) bacteria[J]. Applied and Environmental Microbiology, 2013, 79(13): 4087-4093. DOI:10.1128/AEM.00743-13 |
| [5] | Sumino T, Isaka K, Ikuta H, et al. Nitrogen removal from wastewater using simultaneous nitrate reduction and anaerobic ammonium oxidation in single reactor[J]. Journal of Bioscience and Bioengineering, 2006, 102(4): 346-351. DOI:10.1263/jbb.102.346 |
| [6] | Schrum H N, Spivack A J, Kastner M, et al. Sulfate-reducing ammonium oxidation: a thermodynamically feasible metabolic pathway in subseafloor sediment[J]. Geology, 2009, 37(10): 939-942. DOI:10.1130/G30238A.1 |
| [7] | Canfield D E, Stewart F J, Thamdrup B, et al. A cryptic sulfur cycle in oxygen-minimum-zone waters off the Chilean coast[J]. Science, 2010, 330(6009): 1375-1378. DOI:10.1126/science.1196889 |
| [8] | Fdz-Polanco F, Fdz-Polanco M, Fernández N, et al. Combining the biological nitrogen and sulfur cycles in anaerobic conditions[J]. Water Science & Technology, 2001, 44(8): 77-84. |
| [9] | Fdz-Polanco F, Fdz-Polanco M, Fernandez N, et al. Simultaneous organic nitrogen and sulfate removal in an anaerobic GAC fluidised bed reactor[J]. Water Science & Technology, 2001, 44(4): 15-22. |
| [10] | 董凌霄, 吕永涛, 韩勤有, 等. 硫酸盐还原对氨氧化的影响及其抑制特性研究[J]. 西安建筑科技大学学报(自然科学版), 2006, 38(3): 425-428, 432. DOI:10.3969/j.issn.1006-7930.2006.03.024 |
| [11] | Sabumon P C. Anaerobic ammonia removal in presence of organic matter: a novel route[J]. Journal of Hazardous Materials, 2007, 149(1): 49-59. DOI:10.1016/j.jhazmat.2007.03.052 |
| [12] | Sabumon P C. Development of a novel process for anoxic ammonia removal with sulphidogenesis[J]. Process Biochemistry, 2008, 43(9): 984-991. DOI:10.1016/j.procbio.2008.05.004 |
| [13] | Sabumon P C. Effect of potential electron acceptors on anoxic ammonia oxidation in the presence of organic carbon[J]. Journal of Hazardous Materials, 2009, 172(1): 280-288. DOI:10.1016/j.jhazmat.2009.07.006 |
| [14] | Liu S T, Yang F L, Gong Z, et al. Application of anaerobic ammonium-oxidizing consortium to achieve completely autotrophic ammonium and sulfate removal[J]. Bioresource Technology, 2008, 99(15): 6817-6825. DOI:10.1016/j.biortech.2008.01.054 |
| [15] | Yang Z Q, Zhou S Q, Sun Y B. Start-up of simultaneous removal of ammonium and sulfate from an anaerobic ammonium oxidation (anammox) process in an anaerobic up-flow bioreactor[J]. Journal of Hazardous Materials, 2009, 169(1-3): 113-118. DOI:10.1016/j.jhazmat.2009.03.067 |
| [16] |
袁怡, 黄勇, 李祥, 等. 硫酸盐还原-氨氧化反应的特性研究[J]. 环境科学, 2013, 34(11): 4362-4369. Yuan Y, Huang Y, Li X, et al. Characteristics of sulfate reduction-ammonia oxidation reaction[J]. Environmental Science, 2013, 34(11): 4362-4369. |
| [17] |
张丽, 黄勇, 袁怡, 等. 硫酸盐/氨的厌氧生物转化试验研究[J]. 环境科学, 2013, 34(11): 4356-4361. Zhang L, Huang Y, Yuan Y, et al. Study on the biotransformation of sulfate and ammonia in anaerobic conditions[J]. Environmental Science, 2013, 34(11): 4356-4361. |
| [18] |
完颜德卿, 黄勇, 毕贞, 等. 硫酸盐还原氨氧化体系中基质转化途径[J]. 环境科学, 2017, 38(8): 3406-3414. Wanyan D Q, Huang Y, Bi Z, et al. Conversion pathways of substrates in sulfate-reducing ammonia oxidation system[J]. Environmental Science, 2017, 38(8): 3406-3414. |
| [19] |
董石语, 毕贞, 张文静, 等. ANAMMOX体系中氨与硫酸盐的同步转化条件[J]. 环境科学, 2019, 40(8): 3691-3698. Dong S Y, Bi Z, Zhang W J, et al. Simultaneous conversion conditions of ammonia and sulfate in anammox systems[J]. Environmental Science, 2019, 40(8): 3691-3698. |
| [20] | Bi Z, Wanyan D Q, Li X, et al. Biological conversion pathways of sulfate reduction ammonium oxidation in anammox consortia[J]. Frontiers of Environmental Science & Engineering, 2020, 14(3). DOI:10.1007/s11783-019-1217-1 |
| [21] | van de Graaf A A, de Bruijn P, Robertson L A, et al. Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor[J]. Microbiology, 1996, 142(8): 2187-2196. DOI:10.1099/13500872-142-8-2187 |
| [22] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [23] | Shu D T, He Y L, Yue H, et al. Effects of Fe(Ⅱ) on microbial communities, nitrogen transformation pathways and iron cycling in the anammox process: kinetics, quantitative molecular mechanism and metagenomic analysis[J]. RSC Advances, 2016, 6(72): 68005-68016. DOI:10.1039/C6RA09209H |
| [24] | Harms G, Layton A C, Dionisi H M, et al. Real-time PCR quantification of nitrifying bacteria in a municipal wastewater treatment plant[J]. Environmental Science & Technology, 2003, 37(2): 343-351. |
| [25] | Bi Z, Zhang W J, Song G, et al. Iron-dependent nitrate reduction by anammox consortia in continuous-flow reactors: a novel prospective scheme for autotrophic nitrogen removal[J]. Science of the Total Environment, 2019, 692: 582-588. DOI:10.1016/j.scitotenv.2019.07.078 |
| [26] | Wang S Y, Zhu G B, Peng Y Z, et al. Anammox bacterial abundance, activity, and contribution in riparian sediments of the Pearl River estuary[J]. Environmental Science & Technology, 2012, 46(16): 8834-8842. |
| [27] | Xu X M, Fu S Y, Zhu Q, et al. Depth-related coupling relation between methane-oxidizing bacteria (MOBs) and sulfate-reducing bacteria (SRBs) in a marine sediment core from the Dongsha region, the South China Sea[J]. Applied Microbiology and Biotechnology, 2014, 98(24): 10223-10230. DOI:10.1007/s00253-014-5958-2 |
| [28] | Pester M, Rattei T, Flechl S, et al. amoA-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions[J]. Environmental Microbiology, 2012, 14(2): 525-539. DOI:10.1111/j.1462-2920.2011.02666.x |
| [29] | Prachakittikul P, Wantawin C, Noophan P, et al. Anammox-like performances for nitrogen removal from ammonium-sulfate-rich wastewater in an anaerobic sequencing batch reactor[J]. Journal of Environmental Science and Health, Part A, 2016, 51(3): 220-228. DOI:10.1080/10934529.2015.1094336 |
| [30] | 王慧. 硫酸盐型厌氧氨氧化同步脱氮除硫实验研究[D]. 沈阳: 沈阳工业大学, 2017. |
| [31] | Rozanova E P, Tourova T P, Kolganova T V, et al. Desulfacinum subterraneum sp. nov., a new thermophilic sulfate-reducing bacterium isolated from a high-temperature oil field[J]. Microbiology, 2001, 70(4): 466-471. DOI:10.1023/A:1010494413559 |
| [32] | Ryan R P, Monchy S, Cardinale M, et al. The versatility and adaptation of bacteria from the genus Stenotrophomonas[J]. Nature Reviews Microbiology, 2009, 7(7): 514-525. DOI:10.1038/nrmicro2163 |
| [33] | Chiang Y R, Ismail W, Heintz D, et al. Study of anoxic and oxic cholesterol metabolism by Sterolibacterium denitrificans[J]. Journal of Bacteriology, 2008, 190(3): 905-914. DOI:10.1128/JB.01525-07 |
| [34] | Rikmann E, Zekker I, Tomingas M, et al. Sulfate-reducing anaerobic ammonium oxidation as a potential treatment method for high nitrogen-content wastewater[J]. Biodegradation, 2012, 23(4): 509-524. DOI:10.1007/s10532-011-9529-2 |
| [35] | Rikmann E, Zekker I, Tomingas M, et al. Comparison of sulfate-reducing and conventional anammox upflow anaerobic sludge blanket reactors[J]. Journal of Bioscience and Bioengineering, 2014, 118(4): 426-433. DOI:10.1016/j.jbiosc.2014.03.012 |
| [36] | Rikmann E, Zekker I, Tomingas M, et al. Sulfate-reducing anammox for sulfate and nitrogen containing wastewaters[J]. Desalination and Water Treatment, 2016, 57(7): 3132-3141. DOI:10.1080/19443994.2014.984339 |
| [37] | Zhang Z, Liu S. Insight into the overconsumption of ammonium by anammox consortia under anaerobic conditions[J]. Journal of Applied Microbiology, 2014, 117(6): 1830-1838. DOI:10.1111/jam.12649 |
| [38] | Li W, Zhao Q L, Liu H. Sulfide removal by simultaneous autotrophic and heterotrophic desulfurization-denitrification process[J]. Journal of Hazardous Materials, 2009, 162(2-3): 848-853. DOI:10.1016/j.jhazmat.2008.05.108 |