 2021, Vol. 42
2021, Vol. 42

2. 中国科学院亚热带农业生态研究所, 亚热带农业生态过程重点实验室, 长沙 410125;
3. 贵州大学农学院, 贵阳 550025
 , XIAO Mou-liang2 , JIANG Jia-bin3 , WEI Liang2 , LI Ke-lin1
, XIAO Mou-liang2 , JIANG Jia-bin3 , WEI Liang2 , LI Ke-lin1 , LIU Shou-long2 , ZHU Zhen-ke2 , GE Ti-da2 , WU Jin-shui2
, LIU Shou-long2 , ZHU Zhen-ke2 , GE Ti-da2 , WU Jin-shui2
2. Key Laboratory of Subtropical Agro-Ecological Processes, Institute of Subtropical Agro-Ecology, Chinese Academy of Sciences, Changsha 410125, China;
3. College of Agriculture, Guizhou University, Guiyang 550025, China
土壤有机碳是陆地碳库的重要组成部分, 其积累和分解的变化直接影响全球的碳平衡.土壤有机碳库的大小主要受到外源有机物质输入和土壤有机质(SOM)分解过程的共同调节[1].植物光合碳是土壤有机碳库的重要来源, 每年约有40%的植物光合碳通过根际沉积作用输入土壤, 使土壤有机碳库不断更新累积[2].根际沉积物中包含有碳水化合物、氨基酸和有机酸等可溶物, 为土壤微生物提供有效的碳源和氮源, 改变根际微环境和土壤微生物活性, 并改变SOM在根际土和非根际土之间的分解矿化速率, 即为根际激发效应(RPE)[3, 4].植物根际激发效应导致原土壤有机质矿化有可能抵消部分土壤固碳作用.因此, 研究植物生长过程中的激发效应及其主要影响因子, 对于理解土壤碳矿化与平衡具有重要意义.
根际激发效应的产生是植物根系和微生物以及土壤有机质之间相互作用的结果, 是由根系分泌物驱动, 通过改变微生物量及其活性来实现的, 同时受其他生物与非生物因子的调控[5, 6].根际沉积和根系活性的大小取决于植物的生长, 而植物的生长又反过来影响根际的物理和化学条件, 如水分、氧气浓度、pH和氧化还原电位(Eh)[7, 8].对于农田土壤, 田间管理方式包括灌溉、施氮、秸秆还田等能够通过改变土壤水分和养分条件等非生物因素而影响微生物活性及根系分泌等过程, 从而直接或间接地影响根际激发效应的方向和强度[9~12].稻田土壤水分可以直接改变植物的生长状况、根际沉积量和微生物活性, 进而影响根际的激发效应的强度, 通常较高的土壤含水量会引起更强烈的根际激发效应[13].植物的强烈的蒸腾作用也能使根际发生干湿交替, 短时间和高频率的干湿交替可以促进土壤有机质的矿化[13~15]. Cheng[14]的研究指出大豆的根际激发效应比小麦强烈, 可能是由于大豆比小麦具有持续、更高的蒸腾速率.土壤水分还能通过影响土壤微生物和植物对氮素的竞争[16]、根系分泌物的含量[17]和酶活性[10, 11, 18]等对根际激发效应产生影响.而水稻土中由于长期的干湿交替过程和水稻较强的蒸腾作用, 使得根际沉积作用和微生物活性会随土壤水分状况变化而显著改变.因此, 探讨水分管理对于水稻根际沉积碳的输入与激发效应, 对于研究水稻土固碳机制和优化田间水分管理措施起到积极的促进作用.
目前, 淹水灌溉是我国水稻种植的常规灌溉方式, 人们为了节水减排, 开始探究不同水分管理的灌溉模式.近年来关于水分状况对水稻生长的影响与机制已有较多的报道, 然而不同水分管理会影响植物光合碳在地上/地下的分布, 以及根系微生物的数量和活性, 而水分管理对水稻根际激发效应的方向与强度的影响特征还尚不明确.因此, 本研究采用13C-CO2连续标记技术, 通过设置不同水分管理条件(干湿交替和持续淹水), 探究根际沉积碳输入过程中, 水分管理对水稻光合碳分配及其根际激发效应的影响特征.
1 材料与方法 1.1 供试土壤供试土壤采集自中国科学院长沙农业环境观测研究站(113°19′52″E, 28°33′04″N)连续种稻10年的稻田表层土壤, 该地属亚热带季风湿润气候, 平均气温16~18℃, 年降雨量1200~1700 mm, 无霜期250~300 d.土壤采集时间为2018年5月, 采集耕作层(0~20 cm)土壤, 除去土壤可见植物残体(如根、茎和叶)、石块以及土壤动物等可见杂质, 取1.00 kg原始土样在室内进行风干破碎, 过2 cm筛, 供用于测定土壤的基本理化性质(表 1).供试作物为水稻, 水稻品种为常规迟熟晚稻——华润2号.
|
|
表 1 供试土壤基本理化性质1) Table 1 Soil physiochemical properties |
1.2 水稻种植与13C-CO2连续培养
向预培养后的土壤中施入21 mg·kg-1的(NH4)2SO4、30 mg·kg-1的KH2PO4和122.5 mg·kg-1的KCl作为基肥, 其中以每千克土壤中施入的(NH4)2SO4、NaH2PO4和KCl中的N、P和K的质量(mg)来计, 并加入硝化抑制剂DCD(25 mg·kg-1), 充分混匀.水稻标记组设计2组处理, 每个处理5个重复:①持续淹水(CF); ②干湿交替(AWD); 每组包括:不种植水稻的处理、种植水稻但是不标记和种植水稻且标记.其中, 持续淹水维持水位2~3 cm, 落干处理要求土壤表面无水层或是有轻微土壤裂隙即可, 持续淹水和落干周期为24 d, 设置3次干湿交替循环, 采用原位Eh计检测土壤氧化还原电位变化动态(图 1).将土壤样品混匀分装于PVC盆(直径17.2 cm、高20 cm), 每盆装入相当于1.8 kg干土的土壤, 2018年5月14日播种, 待水稻秧苗长到25 d后, 将其移栽在PVC盆中, 每盆2株, 每个处理12个重复, 用称重法分别控制土壤水分为田间持水量的105%.水稻生长期间, 根据病虫害情况, 喷洒必要的农药.培养装置和方法参考文献[19, 20].

|
CF表示持续淹水, AWD表示干湿交替, 下同 图 1 土壤的氧化还原点位 Fig. 1 Soil redox potential |
待水稻秧苗在PVC盆生长20 d左右, 在每个处理12个重复里面各选择10个长势相当的秧苗作为试验材料.选择在2018年6月29日对水稻进行连续标记; 另设不标记组作为对照试验, 盆栽放置在离标记箱10 m以外, 以防止同位素污染. 13C-CO2通过13C-NaHCO3 (99% atom 13C, 100 mL、1 mol·L-1)和过量的H2SO4(50 mL、0.5 mol·L-1)反应产生, 通过控制13C-NaHCO3加入速度使标记箱内CO2含量维持在350~400 μL·L-1.标记培养箱内外安置两个温度传感器(智能型精密数显温度控制器, SNT-96S, 杭州), 使标记过程中培养箱内外温度控制在±1℃.本试验共设计2次采样, 分别在最后一个干湿交替循环的水稻淹水期结束(63 d)和落干期结束时(75 d)采集气体和植株与土壤样品.
CO2和CH4采用改进的静态箱采气法(图 2)[12].采气时提前1 d, 每个处理随机选取5盆, 检查PVC盆中的水位, 然后将采气装置的P1(根际呼吸CO2和CH4收集室)固定于PVC盆上, 水稻秸秆与P1盖子中间的间隙用硅胶密封严实, 放置1 d后使其固化.采气时调节标记箱的光照与温度, 保证此后每次采样时条件基本一致, 将采气装置的P2(水稻呼吸采气室)固定于P1上, 打开两个风扇, 向采气装置内通入高纯N2 10 min(以排除装置内的CO2和CH4, 同时也保证采气起始点气体浓度一致), 然后封死采气装置的所有加水口和换气口、采气口, 待装置内气体累积1 h后, 在次日的09:00~15:00, 分别抽取两个气室中的气体, 并分别进行气体浓度测试和同位素丰度测试.采气结束后, 对土壤和水稻植株采用破坏性采样, 先将水稻地上部分剪断, 将水稻根系与土壤分离, 用去离子水冲洗干净, 然后将水稻地上部和及其根系置于105℃的烘箱内杀青30 min, 在60℃下烘干至恒量、称重.土壤样品一部分新鲜的土样提取可溶性有机碳氮等基本理化指标, 另一部分自然风干后待测.

|
1.小风扇, 2.温度计, 3.橡胶塞, 4.P1盖子的中孔, 5.水稻植株, 6.螺丝, 7.橡胶塞, 8.水层, 9.水稻根, 10.PVC盆, 11.水稻土; 采气装置包括以下3个部分, PVC盆:水稻种植PVC盆; P1:根际呼吸的CH4收集室; P2:根际呼吸的CO2和CH4收集室 图 2 水稻根际呼吸气体(CO2和CH4)采集装置 Fig. 2 Rice rhizosphere emitted gas (CO2 and CH4) trapping system |
土壤pH(土水体积比为1:2.5)采用pH计(Mettler-toledo 320, 瑞士)测定; 土壤氧化还原电位(Eh)采用Eh计(PRN-41, 日本)测定; 土壤可溶性有机碳采用0.05 mol·L-1 K2SO4浸提, 土壤微生物量采用氯仿熏蒸, 均采用TOC分析仪测定法(TOC-VWP, 日本)[21]; 土壤铵态氮(NH4+)采用0.5 mol·L-1 K2SO4溶液提取, 流动注射仪测定(AA3, 德国). SOC的测定采用干烧法-碳氮元素分析仪(Vario MAX C/N, 德国)[22]; 土壤速效磷(Olsen-P)采用0.5 mol·L-1 NaHCO3溶液浸提, 钼锑抗比色-紫外分光光度计测定(UV2300, 上海). CO2、CH4用温室气相色谱仪直接测定(7890A, 美国); 水稻干物质量(地上部、根系)及其根冠比测定采用65℃烘干后称取干物质质量; 水稻植株(地上部、根系)与土壤样品的13C丰度以及植物碳含量测定采用稳定性同位素质谱仪(MAT253, 美国).
1.4 数据处理通过添加外源13C于稻田土壤中, 土壤原有碳和外源13C矿化作用产生CO2气体, 算式为:
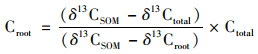
|
(1) |
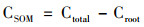
|
(2) |

|
(3) |
式中, Ctotal为来源于水稻体系中总释放的CO2; δ13Ctotal为来源于水稻体系中总释放的13C丰度; δ13CSOM为来源于土壤有机质释放的13C-CO2丰度; δ13Croot为来源于根呼吸释放的13C-CO2丰度; CSOM, Planted为来源于种植水稻土壤有机质释放的CO2; CSOM, Unplanted为来源于未种植水稻土壤有机质释放的CO2.
当来源于种植水稻土壤有机质释放的CO2小于来源于未种植水稻土壤有机质释放的CO2, 即公式(3)中RPE小于零则为负激发效应, 反之为正激发效应.
土壤原有碳和添加外源13C矿化作用的CH4产生量计算原理同上.
方差分析用SPSS 17.0软件计算.同一组分不同生育期的干物质累积量、根冠比、地上部占植株总质量的比例、土壤可溶性有机碳、微生物生物量碳、CO2和CH4激发效应与土壤环境因子间的关系显著性差异分析均设为P < 0.05水平, 且均值比较采用最小显著差异法(least significant difference, LSD).制图采用Origin 9.1软件.
2 结果与分析 2.1 水分管理对水稻干物质量及其根冠比的影响水稻移栽63 d后, 持续淹水条件下的水稻地上部和根系的生物量分别为329 kg·hm-2和98 kg·hm-2, 而干湿交替比持续淹水条件下的水稻地上部和根系的生物量分别增加了22 kg·hm-2和19 kg·hm-2.在经过最后一个落干周期后(75 d), 水稻地上部生物量基本没有增加, 但是干湿交替处理的根系的生物量增加了14 kg·hm-2.干湿交替处理显著增加了水稻根冠比, 而且在75 d时, 干湿交替相比持续淹水使水稻根冠比增加了5%(图 3).

|
图 3 水分管理对水稻干物质量及根冠比 Fig. 3 Rice dry biomass and the ratio of rice root to shoot |
土壤可溶性有机碳含量随水稻生长快速增加[图 4(a)].持续淹水条件下, 土壤可溶性有机碳由12.5 mg·kg-1(63 d)增加到29.6 mg·kg-1(75 d), 增加了137%.而干湿交替条件下土壤可溶性有机碳由1.8 mg·kg-1(63 d)快速增加到19.8 mg·kg-1(75 d), 增加了约10倍.土壤微生物生物量碳随水稻生长的变化主要受到水分管理的影响[图 4(b)].土壤微生物生物量碳受水分管理的影响与可溶性有机碳相反, 干湿交替处理增加了微生物生物量碳.持续淹水条件下, 微生物生物量碳在两个采样点变化不大; 但是, 干湿交替处理下, 微生物生物量碳由332.3 mg·kg-1(63 d)增加到516.2 mg·kg-1(75 d), 增加了55%.

|
图 4 不同水分管理条件下土壤可溶性有机碳和微生物生物量碳 Fig. 4 Soil dissolved C and microbial biomass C in response to water management |
在水稻-土壤体系中, 不同水分管理方式对CO2和CH4排放通量有显著的影响(图 5).持续淹水条件下的CO2和CH4排放通量随水稻生长显著增加, 而干湿交替处理下, 在最后一个落干循环中CO2和CH4排放通量随水稻生长却显著降低.持续淹水条件下, 来源于根际沉积碳的13CO2和13CH4排放通量, 在随水稻生长由10.2 μg·(kg·h)-1和2.8 μg·(kg·h)-1(63 d)增加到16.0 μg·(kg·h)-1和3.2 μg·(kg·h)-1(75 d).而在12 d的落干处理过程中, 来源于根际沉积碳的13CO2和13CH4排放通量分别降低了57.5%和88.1%.

|
图 5 CO2和CH4排放通量对水分管理的响应 Fig. 5 CO2 and CH4 fluxes in response to water management |
水稻根际沉积碳对土壤原有有机碳矿化的激发效应, 因水分管理方式而差异明显.持续淹水条件下, CO2和CH4的根际激发效应表现为正激发效应, 而且随水稻的生长由0.06 mg·(kg·h)-1和7.7 μg·(kg·h)-1(63 d)增加到0.32 mg·(kg·h)-1和24.0 μg·(kg·h)-1(75 d).而干湿交替处理下, 经过12 d的落干处理(图 6), CO2和CH4的根际激发效应分别由0.29 mg·(kg·h)-1和12.3 μg·(kg·h)-1(63 d)降低到-0.39 mg·(kg·h)-1和0.07 μg·(kg·h)-1(75 d).相关性分析发现, CO2根际激发效应与Fe2+、NH4+和MBN呈显著的正相关关系; CH4根际激发效应与Fe2+、NH4+、MBN和CO2激发效应呈显著的正相关关系(表 2).

|
图 6 CO2和CH4根际激发效应对水分管理的响应 Fig. 6 Response of CO2 and CH4 rhizosphere priming effects to water management |
|
|
表 2 CO2和CH4激发效应与土壤环境因子间的相关性分析1) Table 2 Relationships between CO2 and CH4 priming effects and soil environmental factors |
3 讨论 3.1 水分管理对水稻植株干生物量及根冠比的影响
水分在作物的生长发育过程中起着重要作用, 它通过水稻植株本身复杂的生理过程而影响植物的生长状况[23].与传统农业灌溉方式相比, 干湿交替灌溉对作物产量的影响结果将会产生差异[24], 但是结果也不尽相同, 可能是因为土壤和气候条件, 以及水稻品种特性等, 干湿交替灌溉会降低作物生长和产量[25, 26], 或是促进作物生长和产量提升[27].本研究结果表明干湿交替水分管理能显著提高水稻的地上部和地下部生物量以及根冠比, 促进了水稻的生长.这主要是因为干湿交替处理下改变了水稻土壤的通气性和氧化还原条件, 一方面促进水稻根尖伸长和细根发育, 增加根系与土壤的接触面积, 促进根系对养分的吸收能力, 加快植物的生长发育; 另一方面是增加了土壤微生物数量和活性, 促进了土壤养分的活化与供给, 进而加快了对养分的吸收和有机质分解[28~30].而持续淹水会导致土壤通气性差、水稻根系发育不良、无效分蘖增加而抑制水稻的生长[31, 32].因此, 相对于淹水处理, 干湿交替更易加快根系生长和土壤养分活化, 从而促进水稻植株的生长发育.
3.2 水分管理对水稻根际CO2激发效应的影响不同的水分管理方式通过改变土壤的水分条件而影响土壤氧化还原电位、土壤含氧量、硝化与反硝化作用强度以及气体传输途径影响温室气体的产生与释放量[28].干湿交替的淹水周期结束时(63 d), 干湿交替条件下CO2的排放量高于持续淹水下的CO2的排放量[(图 5(a)和图 5(b)].这主要是因为干湿交替处理, 促进了水稻的生长, 增加了根冠比, 提高了根系活力(图 1).根系的生长也会促进根系分泌物的增加以及微生物的生长[图 4(b)], 从而促进了微生物呼吸产生更多的CO2[12].但在落干周期结束时(75 d), 干湿交替下CO2的排放量显著降低, 而低于持续淹水下的CO2的排放量.研究表明随落干和复水次数的增加, 土壤CO2释放速率会随之减少[33], 可能的原因是水稻管理方式将会影响土壤中微生物呼吸作用, 使其强度减弱, 因而导致CO2释放量减少[34].此外, 土壤落干过程中, 由于土壤中会形成新的弱结晶态矿物, 增加了土壤可溶性有机碳的吸附, 降低了微生物的可接触性, 进而减少了CO2的排放量[35, 36].
CO2根际激发效应的规律与CO2排放通量的规律一致; 即持续淹水过程中, 随着水稻的生长CO2根际激发效应逐渐增加, 而干湿交替处理中, 在落干期CO2根际激发效应由正降低为负.在持续淹水过程中, 水稻的生长增加了土壤可溶性有机碳的含量, 促进微生物的生长, 增加了对根系分泌物和土壤原有有机碳的分解矿化[12, 37].而且, 厌氧条件下土壤矿物还原溶解过程中会促进土壤原有有机质的溶出, 使得易于被微生物利用, 因而表现出逐渐增加的正激发效应[38].而土壤在由淹水逐渐落干的过程中, 土壤含水率急剧降低, 分泌物难以快速扩散以供应土壤微生物利用, 使微生物活性降低, 抑制了其对土壤原有有机质的分解速率[38, 39].而落干过程中, 新形成的土壤矿物对有机碳的封存作用进一步降低了微生物对原有有机碳的矿化, 因而导致了在落干结束时CO2根际激发效应表现为负值[40~42].
3.3 水分管理对水稻根际CH4激发效应的影响CH4的产生是厌氧分解的最后一步, 其产生速率与土壤氧化还原电位密切相关, 较低的氧化还原电位可以促进甲烷产生; 在持续淹水的水稻土中由于较低的氧气含量使得土壤氧化还原电位较低(图 1)[43, 44], 因而产甲烷量和甲烷的激发效应都高于干湿交替处理的土壤[图 5(c)、图 5(d)和图 6].而且, 持续淹水处理中, 随着水稻的生长, 铁还原速率增加, 促进了甲烷的产生.但是干湿交替处理中, 落干过程中由于土壤氧气含量增加, 抑制了甲烷产生.持续淹水处理的CH4根际激发效应始终表现为正激发效应.这与之前的研究结果一致, 根系分泌物是厌氧条件下微生物基质的重要来源[45].相对于干湿交替, 持续淹水下来源于土壤原有有机质的CH4的释放和来源于水稻新输入的根际沉积碳的CH4的释放都显著高于干湿交替下CH4的释放.这可能是:①持续淹水条件下形成的缺氧环境, 有利于土壤中还原型微生物的生长, 可促进水稻土中铁还原过程, 加快土壤矿物固定的养分元素释放到土壤, 使产甲烷菌活性得以提高, 因此提高了微生物对土壤有机质的分解矿化作用, 增强了CH4释放的根际激发效应[38, 40, 42]. ② CH4排放是由土壤中CH4产生、氧化及向大气传输3个过程相互作用的结果. CH4产生是土壤厌氧条件下产甲烷菌作用于产甲烷基质的结果, 但在干湿交替条件下, 充足的氧分会使甲烷氧化菌对多数的CH4进行氧化, 就造成了稻田土壤中生成的CH4在排放到大气前约80%~94%被氧化[46, 47]. Yagi等[48]研究了持续淹水及水分排干对稻田CH4排放的影响, 结果表明水稻生长期几次短时间的水分排干大大降低了CH4排放量.蔡祖聪[49]通过培养试验研究淹水、好气(70%WHC)和淹水好气交替的不同水分类型对土壤温室气体排放的影响, 结果表明连续淹水条件下土壤排放出大量的CH4, 好气条件下土壤不排放CH4, 淹水好气交替处理的土壤排放的CH4在好气和连续淹水之间.因此, 干湿交替水分管理可以抑制甲烷产生并降低根际激发效应.
4 结论(1) 干湿交替相对于持续淹水增加了水稻植株干物质量和根冠比.干湿交替过程增加了CO2排放通量, 却降低了CH4排放通量.
(2) 持续淹水条件下, CO2的根际激发效应, 表现为持续的正激发效应, 而在土壤落干处理后表现为持续的负激发效应. CO2根际激发效应主要受Fe2+、NH4+和MBN的影响.
(3) 干湿交替条件下, 水稻土产CH4显著低于持续淹水处理, 而且落干处理过程中降低了CH4的根际激发效应.持续淹水条件下, CH4的根际激发效应表现出了较强的正激发效应.
致谢: 本研究的实施场所以及样品测定工作由中国科学院亚热带农业研究所提供, 谨表感谢.
| [1] | Lal R. Soil carbon sequestration to mitigate climate change[J]. Geoderma, 2004, 123(1-2): 1-22. DOI:10.1016/j.geoderma.2004.01.032 |
| [2] | Gleixner G, Poirier N, Bol R, et al. Molecular dynamics of organic matter in a cultivated soil[J]. Organic Geochemistry, 2002, 33(3): 357-366. DOI:10.1016/S0146-6380(01)00166-8 |
| [3] | Kuzyakov Y. Review:factors affecting rhizosphere priming effects[J]. Journal of Plant Nutrition and Soil Science, 2002, 165(4): 382-396. DOI:10.1002/1522-2624(200208)165:4<382::AID-JPLN382>3.0.CO;2-# |
| [4] | Philippot L, Raaijmakers J M, Lemanceau P, et al. Going back to the roots:the microbial ecology of the rhizosphere[J]. Nature Reviews Microbiology, 2013, 11(11): 789-799. DOI:10.1038/nrmicro3109 |
| [5] | Kemmitt S J, Lanyon C V, Waite I S, et al. Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass-a new perspective[J]. Soil Biology and Biochemistry, 2008, 40(1): 61-73. DOI:10.1016/j.soilbio.2007.06.021 |
| [6] | Kuzyakov Y. Priming effects:interactions between living and dead organic matter[J]. Soil Biology and Biochemistry, 2010, 42(9): 1363-1371. DOI:10.1016/j.soilbio.2010.04.003 |
| [7] | Cheng W X, Johnson D W, Fu S L. Rhizosphere effects on decomposition[J]. Soil Science Society of America Journal, 2003, 67(5): 1418-1427. DOI:10.2136/sssaj2003.1418 |
| [8] | Yuan Q, Pump J, Conrad R. Straw application in paddy soil enhances methane production also from other carbon sources[J]. Biogeosciences Discussions, 2013, 10(8): 14169-14193. DOI:10.5194/bgd-10-14169-2013 |
| [9] | Soon Y K. Crop residue and fertilizer management effects on some biological and chemical properties of a Dark Grey Solod[J]. Canadian Journal of Soil Science, 1998, 78(4): 707-713. DOI:10.4141/S98-016 |
| [10] | Geisseler D, Horwath W R, Scow K M. Soil moisture and plant residue addition interact in their effect on extracellular enzyme activity[J]. Pedobiologia, 2011, 54(2): 71-78. DOI:10.1016/j.pedobi.2010.10.001 |
| [11] | Burns R G, Deforest J L, Marxsen J, et al. Soil enzymes in a changing environment:current knowledge and future directions[J]. Soil Biology and Biochemistry, 2013, 58: 216-234. DOI:10.1016/j.soilbio.2012.11.009 |
| [12] | Zhu Z K, Ge T D, Liu S L, et al. Rice rhizodeposits affect organic matter priming in paddy soil:the role of N fertilization and plant growth for enzyme activities, CO2 and CH4 emissions[J]. Soil Biology and Biochemistry, 2018, 116: 369-377. DOI:10.1016/j.soilbio.2017.11.001 |
| [13] | Dijkstra F A, Cheng W X. Interactions between soil and tree roots accelerate long-term soil carbon decomposition[J]. Ecology Letters, 2007, 10(11): 1046-1053. DOI:10.1111/j.1461-0248.2007.01095.x |
| [14] | Cheng W X. Rhizosphere priming effect:its functional relationships with microbial turnover, evapotranspiration, and C-N budgets[J]. Soil Biology and Biochemistry, 2009, 41(9): 1795-1801. DOI:10.1016/j.soilbio.2008.04.018 |
| [15] | Fierer N, Schimel J P. A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil[J]. Soil Science Society of America Journal, 2003, 67(3): 798-805. DOI:10.2136/sssaj2003.7980 |
| [16] | Liljeroth E, Kuikman P, van Veen J A. Carbon translocation to the rhizosphere of maize and wheat and influence on the turnover of native soil organic matter at different soil nitrogen levels[J]. Plant and Soil, 1994, 161(2): 233-240. DOI:10.1007/BF00046394 |
| [17] | Gorissen A, Tietema A, Joosten N N, et al. Climate change affects carbon allocation to the soil in shrublands[J]. Ecosystems, 2004, 7(6): 650-661. |
| [18] | Bell T H, Henry H A L. Fine scale variability in soil extracellular enzyme activity is insensitive to rain events and temperature in a mesic system[J]. Pedobiologia, 2011, 54(2): 141-146. DOI:10.1016/j.pedobi.2010.12.003 |
| [19] | Ge T D, Chen X J, Yuan H Z, et al. Microbial biomass, activity, and community structure in horticultural soils under conventional and organic management strategies[J]. European Journal of Soil Biology, 2013, 58: 122-128. DOI:10.1016/j.ejsobi.2013.07.005 |
| [20] | Ge T D, Wu X M, Chen X J, et al. Microbial phototrophic fixation of atmospheric CO2 in China subtropical upland and paddy soils[J]. Geochimica et Cosmochimica Acta, 2013, 113: 70-78. DOI:10.1016/j.gca.2013.03.020 |
| [21] | 吴金水, 林启美, 黄巧云, 等. 土壤微生物生物量测定方法及其应用[M]. 北京: 气象出版社, 2006. |
| [22] | 鲍士旦. 土壤农化分析[M]. ((第三版)). 北京: 中国农业出版社, 2005. |
| [23] |
谭立敏.施氮对光合碳在水稻-土壤-微生物系统中分配的影响[D].长沙: 中南林业科技大学, 2014. Tan L M. Allocation of photosynthesized carbon to rice-soil-microbial community system under nitrogen application[D]. Changsha: Central South University of Forestry and Technology, 2014. |
| [24] | Mishra H S, Rathore T R, Pant R C. Effect of intermittent irrigation on groundwater table contribution, irrigation requirement and yield of rice in Mullisols of the Tarai region[J]. Agricultural Water Management, 1990, 18(3): 231-241. DOI:10.1016/0378-3774(90)90045-Z |
| [25] | Won J G, Choi J S, Lee S P, et al. Water saving by shallow intermittent irrigation and growth of rice[J]. Plant Production Science, 2005, 8(4): 487-492. DOI:10.1626/pps.8.487 |
| [26] | Yao F X, Huang J L, Cui K H, et al. Agronomic performance of high-yielding rice variety grown under alternate wetting and drying irrigation[J]. Field Crops Research, 2012, 126: 16-22. DOI:10.1016/j.fcr.2011.09.018 |
| [27] | Tabbal D F, Bouman B A M, Bhuiyan S I, et al. On-farm strategies for reducing water input in irrigated rice; case studies in the Philippines[J]. Agricultural Water Management, 2002, 56(2): 93-112. DOI:10.1016/S0378-3774(02)00007-0 |
| [28] |
张静, 刘娟, 陈浩, 等. 干湿交替条件下稻田土壤氧气和水分变化规律研究[J]. 中国生态农业学报, 2014, 22(4): 408-413. Zhang J, Liu J, Chen H, et al. Change in soil oxygen and water contents under alternate wetting and drying in paddy fields[J]. Chinese Journal of Eco-Agriculture, 2014, 22(4): 408-413. |
| [29] |
王婷婷, 祝贞科, 朱捍华, 等. 施氮和水分管理对光合碳在土壤-水稻系统间分配的量化研究[J]. 环境科学, 2017, 38(3): 1227-1234. Wang T T, Zhu Z K, Zhu H H, et al. Input and distribution of photosynthesized carbon in soil-rice system affected by water management and nitrogen fertilization[J]. Environmental Science, 2017, 38(3): 1227-1234. |
| [30] |
侯海军, 张文钊, 沈建林, 等. 水分管理对稻田细菌丰度与群落结构的影响[J]. 生态环境学报, 2016, 25(9): 1431-1438. Hou H J, Zhang W Z, Shen J L, et al. Effect of water management on soil bacterial abundance and community in the rice paddy field[J]. Ecology and Environmental Sciences, 2016, 25(9): 1431-1438. |
| [31] | Ye Y S, Liang X Q, Chen Y X, et al. Alternate wetting and drying irrigation and controlled-release nitrogen fertilizer in late-season rice. effects on dry matter accumulation, yield, water and nitrogen use[J]. Field Crops Research, 2013, 144: 212-224. DOI:10.1016/j.fcr.2012.12.003 |
| [32] |
姜萍, 袁永坤, 朱日恒, 等. 节水灌溉条件下稻田氮素径流与渗漏流失特征研究[J]. 农业环境科学学报, 2013, 32(8): 1592-1596. Jiang P, Yuan Y K, Zhu R H, et al. Study on the nitrogen loss from paddy fields on different water management[J]. Journal of Agro-Environment Science, 2013, 32(8): 1592-1596. |
| [33] | Schimel J, Balser T C, Wallenstein M. Microbial stress-response physiology and its implications for ecosystem function[J]. Ecology, 2007, 88(6): 1386-1394. DOI:10.1890/06-0219 |
| [34] | Steenwerth K L, Jackson L E, Calderón F J, et al. Response of microbial community composition and activity in agricultural and grassland soils after a simulated rainfall[J]. Soil Biology and Biochemistry, 2005, 37(12): 2249-2262. DOI:10.1016/j.soilbio.2005.02.038 |
| [35] | ThomasArrigo L K, Byrne J M, Kappler A, et al. Impact of organic matter on iron(Ⅱ)-catalyzed mineral transformations in ferrihydrite-organic matter coprecipitates[J]. Environmental Science & Technology, 2018, 52(21): 12316-12326. |
| [36] | Wei X M, Zhu Z K, Wei L, et al. Biogeochemical cycles of key elements in the paddy-rice rhizosphere:Microbial mechanisms and coupling processes[J]. Rhizosphere, 2019, 10. DOI:10.1016/j.rhisph.2019.100145 |
| [37] | Liu X J A, Finley B K, Mau R L, et al. The soil priming effect:consistent across ecosystems, elusive mechanisms[J]. Soil Biology and Biochemistry, 2020, 140. DOI:10.1016/j.soilbio.2019.107617 |
| [38] | Pan W N, Kan J J, Inamdar S, et al. Dissimilatory microbial iron reduction release DOC (dissolved organic carbon) from carbon-ferrihydrite association[J]. Soil Biology and Biochemistry, 2016, 103: 232-240. DOI:10.1016/j.soilbio.2016.08.026 |
| [39] | Gordon H, Haygarth P M, Bardgett R D. Drying and rewetting effects on soil microbial community composition and nutrient leaching[J]. Soil Biology and Biochemistry, 2008, 40(2): 302-311. DOI:10.1016/j.soilbio.2007.08.008 |
| [40] | Canarini A, Dijkstra F A. Dry-rewetting cycles regulate wheat carbon rhizodeposition, stabilization and nitrogen cycling[J]. Soil Biology and Biochemistry, 2015, 81: 195-203. DOI:10.1016/j.soilbio.2014.11.014 |
| [41] | Atere C T, Ge T D, Zhu Z K, et al. Assimilate allocation by rice and carbon stabilisation in soil:effect of water management and phosphorus fertilisation[J]. Plant and Soil, 2019, 445(1-2): 153-167. DOI:10.1007/s11104-018-03905-x |
| [42] | Mikha M M, Rice C W, Milliken G A. Carbon and nitrogen mineralization as affected by drying and wetting cycles[J]. Soil Biology and Biochemistry, 2005, 37(2): 339-347. DOI:10.1016/j.soilbio.2004.08.003 |
| [43] | Conrad R, Klose M, Yuan Q, et al. Stable carbon isotope fractionation, carbon flux partitioning and priming effects in anoxic soils during methanogenic degradation of straw and soil organic matter[J]. Soil Biology and Biochemistry, 2012, 49: 193-199. DOI:10.1016/j.soilbio.2012.02.030 |
| [44] | Ye R Z, Doane T A, Morris J, et al. The effect of rice straw on the priming of soil organic matter and methane production in peat soils[J]. Soil Biology and Biochemistry, 2015, 81: 98-107. DOI:10.1016/j.soilbio.2014.11.007 |
| [45] |
杨智仙, 汤利, 郑毅, 等. 不同品种小麦与蚕豆间作对蚕豆枯萎病发生、根系分泌物和根际微生物群落功能多样性的影响[J]. 植物营养与肥料学报, 2014, 20(3): 570-579. Yang Z X, Tang L, Zheng Y, et al. Effects of different wheat cultivars intercropped with faba bean on faba bean Fusarium wilt, root exudates and rhizosphere microbial community functional diversity[J]. Journal of Plant Nutrition and Fertilizers, 2014, 20(3): 570-579. |
| [46] | Wu X, Yao Z, Brüggemann N, et al. Effects of soil moisture and temperature on CO2 and CH4 soil-atmosphere exchange of various land use/cover types in a semi-arid grassland in Inner Mongolia, China[J]. Soil Biology and Biochemistry, 2010, 42(5): 773-787. DOI:10.1016/j.soilbio.2010.01.013 |
| [47] | Fechner E J, Hemond H F. Methane transport and oxidation in the unsaturated zone of a Sphagnum peatland[J]. Global Biogeochemistry Cycles, 1992, 6(1): 33-34. DOI:10.1029/91GB02989 |
| [48] | Yagi K, Tsuruta H, Kanda K I, et al. Effect of water management on methane emission from a Japanese rice paddy field:automated methane monitoring[J]. Global Biogeochemical Cycles, 1996, 10(2): 255-267. DOI:10.1029/96GB00517 |
| [49] |
蔡祖聪. 水分类型对土壤排放的温室气体组成和综合温室效应的影响[J]. 土壤学报, 1999, 36(4): 484-491. Cai Z C. Effects of water regime on CO2, CH4 and N2O emissions and overall potential for greenhouse effect caused by emitted gases[J]. Acta Pedologica Sinica, 1999, 36(4): 484-491. DOI:10.3321/j.issn:0564-3929.1999.04.008 |