 2021, Vol. 42
2021, Vol. 42

2. 中国环境监测总站, 北京 100012
 , CHEN Wei1 , ZHAO Ran1 , LI Zhong-yu1 , DONG Ying-li1 , ZHANG Rui1 , WANG Ye-yao2 , JIN Xiao-wei2
, CHEN Wei1 , ZHAO Ran1 , LI Zhong-yu1 , DONG Ying-li1 , ZHANG Rui1 , WANG Ye-yao2 , JIN Xiao-wei2
2. China National Environment Monitoring Centre, Beijing 100012, China
随着经济的高速发展、人口的迅猛增长、城市化过程的加快, 人类活动对河流及区域生态系统的影响日益严重, 水生态系统遭受了严重破坏, 导致河流污染加重、生态环境和生物栖息地的退化、水体生物多样性丧失等问题[1, 2].着生藻类作为河流生态系统中食物链最底端的初级生产者, 具有生产迅速、分布广泛、对生境条件要求低、适应性强、种类较多和采集便捷等特点[3, 4], 是组成水生态系统中的物质循环、能量流动和信息传递等功能的重要部分[5, 6].藻类的群落与环境因子有着紧密的关联性, 其种类的组成和分布变化对环境的变化具有指示作用.因此, 着生藻类的群落特征常被用作评价水生态环境质量的重要指标[7~9].
我国重要七大水系之一的松花江, 地跨黑龙江、辽宁、吉林和内蒙古这4个省区, 流域广阔.流域地处北温带季风气候, 四季分明, 夏季温热多雨且短促, 冬季寒冷干燥而又漫长, 年内温差大, 冰封期长达5~6个月.随着流域内的工业化发展、城镇化提速和农业发展的不断扩大, 水体受到了不同程度的污染. “十二五”以来, 松花江流域被国家列为重点流域水污染防治规划的重要部分, 水环境质量得到了明显的改善, 水生态系统逐渐得到恢复, Ⅲ类水体比例增长10.4%, Ⅳ类和Ⅴ类水体占比下降5.2%, 劣Ⅴ类水体占比下降6.7%.而常规开展的理化监测指标(如COD、氨氮和BOD5)很难满足水生态环境管理的需求, 不能准确反映复杂水生态环境质量的变化趋势, 需要通过开展生物条件以及环境因子的综合监测与评价, 准确评价流域水生态环境质量现状和预测变化趋势[10].关于松花江着生藻类与水质特征的研究较少, 仅有针对个别年份或部分河段的报道, 部分研究显示松花江干流藻类评价结果水体呈中污染状态[11].目前鲜见对松花江流域着生藻类长期群落演变规律及其与环境因子相关性分析的研究, 而时间尺度的演替规律更能真实和直观地反映流域水生态环境质量的变化趋势和治理成效[12].
本文首次通过对松花江干流2014~2019年夏季着生藻类的群落结构与年际变化情况进行分析, 并基于冗余分析(redundancy analysis, RDA)解释了藻类植物对环境变量之间的响应关系, 以期为松花江流域水生态环境质量评价与水生态修复保护提供基础支撑.
1 材料与方法 1.1 采样点设置松花江干流指松花江南源(正源), 自长白山天池流下的西流, 从吉林省松原市嫩江注入, 汇入黑龙江的河口, 全长958 km, 流域面积7.34万km2, 占流域总面积的14.33%, 其供给松花江39%的水量.上游江段由三岔河至哈尔滨市, 全长240 km, 哈尔滨段人为干扰严重, 流域内支流较少, 附近右岸有大支流拉林河汇入; 中游江段由哈尔滨市至佳木斯市, 河道长432 km, 江道比较平缓, 左岸有最大的支流呼兰河汇入; 下游江段由佳木斯至同江, 全长267 km, 穿行于三江平原地区, 两岸为冲积平原, 地势平坦, 杂草丛生, 河道和滩地比较开阔[13].
2014~2019年, 按照每月采样1次的频率对松花江干流着生藻类开展4个月的样品采集并同时采集平行样, 流域自上而下共设置11个采样点位如(图 1), 分别为肇源(D1)、朱顺屯(D2)、阿什河口下(D3)、呼兰河口下(D4)、大顶子山(D5)、摆渡镇(D6)、牡丹江口下(D7)、佳木斯上(D8)、佳木斯下(D9)、江南屯(D10)和同江(D11), 中上游点位D2、D3、D5、D6和D7受到采沙场、村庄、农田灌溉及生活污水等不同人为因素的干扰, 其它点位D1和D4、D8、D9、D10和D11植被覆盖率高, 水质较清洁, 受人为干扰因素小.

|
图 1 松花江干流采样点示意 Fig. 1 Sampling sites in the main stream of the Songhua River |
本文数据来源于黑龙江省生态环境监测中心2014~2019年“松花江流域水生生物试点监测”工作.定性采样安排在放样当天, 采用天然基质作为定性采样器材, 以采样点周围的植物叶片、石块和木块等作为天然基质, 尽可能多地采集不同的基质, 要记录基质的名称, 并填写采样记录.定量采样采用人工基质硅藻计-载玻片法(用木材制作的硅藻计固定载玻片(26 mm×76 mm, 8片).人工基质作为定量采样器材必须固定好, 可缚在钉入河流底部的钢筋或其他结构上.采样器的放置深度一般为5~10 cm, 使之得到合适的光照.每个采样点至少放置2个人工基质, 确保样品采集成功.放置时间一般为14 d, 人工基质经过2~4周的培养可以代表天然基质上着生藻类群落[14].定性样品的保存装入盛有少量水的塑料袋里, 定量样品采样现场将所取基质放入装有采样点水样的广口瓶中, 贴好标签, 做好记录, 带回实验室.用牙刷、毛刷或者硬胶皮将基质上的藻类全部刮到盛有蒸馏水的烧杯中.当基质的手感从光滑、粘腻变为粗糙、不黏时, 才能判断着生藻类已经被完全取下, 并用蒸馏水将基质冲洗多次, 用鲁哥氏液固定, 经24 h沉淀, 弃去上清液, 将剩余的液体搅动, 虹吸法吸取沉淀.定性样品无需定容, 定量样品定容至30 mL, 取0.1 mL置于0.1 mL的计数框内进行计算. 40×显微镜下, 将藻类鉴定至属.种类鉴定主要参考文献[15].
现场测定部分水体的理化参数并现场采集其它理化指标的水样. pH、水温、电导率和溶解氧用Hydrolab DS5分析仪测定, 其它参数测定依据文献[16]进行测定.
1.3 数据分析本文采用Shannon-Wiener和Pielou描述群落结构的多样性.详细公式如下[17, 18].
Shannon-Wiener多样性指数(H′):
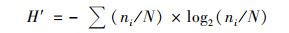
|
式中, H′表示物种多样性指数; ni表示第i种的藻类植物个体数; N表示藻类植物总数.
Pielou均匀度指数(J):
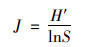
|
式中, J表示均匀度指数; H′表示Shannon-Wiener多样性指数; S表示藻类植物属数.
2个指数的评价标准如表 1所示.
|
|
表 1 Shannon-Wiener多样性指数和Pielou均匀度指数评价标准[18] Table 1 Evaluation criteria of Shannon-Wiener diversity index and Pielou evenness index |
藻类植物的优势度(Y )[19]:
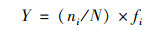
|
式中, Y表示优势度; ni表示第i种的总个数; fi表示第i种在各样点出现的频率.
采用Canoco 4.5软件对物种数据和环境因子进行去趋势对应的分析(DCA), 由于最大长度均小于3为RDA, 3~4之间取RDA和CCA均可, 故本研究选择了典范对应分析(RDA)来进行分析, 数据分析过程中将着生藻类的细胞密度与环境因子数据进行lg(x+1)转换, 用于保证获得正态分布[20].
2 结果与分析 2.1 水质变化整体趋势2014~2019年松花江干流水质理化指标变化趋势显示(图 2), 年平均值最高温度为2017年的13.73℃, 年平均值最低温度为2015年的11.02℃; 水体pH值年际间差异较小, 平均值为7.7; COD在6 a总体平稳, 最高值为2014年的19.03 mg·L-1, 最低值为2016年的17.35 mg·L-1, 2016年后呈逐年上升趋势; BOD峰值出现在2015年, 为3.30 mg·L-1, 2015年后呈逐年下降趋势, 最低值为2019年的2.01 mg·L-1; DO在6 a保持在较高水平, 最高值为2019年的9.00 mg·L-1, 最低值为2017年的8.53 mg·L-1, 6 a平均值为8.82 mg·L-1; NH4+-N指标呈总体下降趋势, 其中2014年出现峰值为0.57 mg·L-1, 其后除2018年较2017年有所上升外, 各年指标均较上一年下降, 2019年为最低值0.25 mg·L-1; TP峰值为2014年的0.14mg·L-1, 最低值为2016年的0.10 mg·L-1, 该指标呈现先降低后升高再降低趋势, 2015年与2016年下降, 于2016年达到低点后, 2017年和2018年数值升高, 而2019年则较2018年有所降低. TN与TP变化趋势相似, 2015年较2014年有所降低, 2016年后逐年上升, 2018年达到峰值为2.17 mg·L-1, 2019年下降并达到最低值1.87 mg·L-1.

|
图 2 2014~2019年松花江干流主要水质指标变化趋势 Fig. 2 Variations in the water quality indexes in the main stream of the Songhua River from 2014 to 2019 |
以2014年为基准, 6 a的松花江干流水域主要水质指标呈波动向好趋势. NH4+-N、DO、TP和TN均在前期下降, 但2017和2018年各项指标较其余4 a较差, 而在2019年得到改善.目前松花江干流水域主要污染物为氮和磷, 该类污染物在2019年均呈现下降趋势, NH4+-N较上一年下降50.98%; TN较上一年下降13.82%; TP较上一年下降23.08%, 理化指标虽相比2014年有所好转, 但营养盐浓度值并未达到降低水体中藻类生物量的程度[21].
2.2 着生藻类群落结构2014~2019年夏季松花江干流共鉴定出着生藻类4门、6纲、17目、26科和58属.其中硅藻门28属, 占总数48.28%;绿藻门17属, 占总数29.31%;蓝藻门10属, 占总数17.24%;裸藻门3属, 占总数5.17%.松花江干流着生藻类的属级分类数呈现为先上升后下降的趋势, 其中2016年最高(49属), 2014年最低(32属).各年均以硅藻门植物居多, 绿藻门植物居中, 蓝藻门种类数量最少(图 3).优势属多为指示中污染或轻污染的种类, 如小环藻属(Cyclotella)、直链藻属(Melosira)、针杆藻属(Synedra)、异极藻属(Gomphonema)、舟形藻属(Navicula)和小球藻属(Chlorella)等, 其中2015、2016和2018年种类构成较2014、2017和2019年丰富.

|
图 3 2014~2019年夏季松花江干流着生藻类的种类组成和群落结构特征 Fig. 3 Summer-time species composition and community structure of periphytic algae in the main stream of the Songhua River from 2014 to 2019 |
调查期间松花江干流着生藻类细胞密度变化范围为1.29×104~8.42×104 ind·cm-3(图 4), 总平均细胞密度为4.35×104ind·cm-3, 其中硅藻门为2.34×104 ind·cm-3, 占总平均细胞密度的53.79%;绿藻门为1.12×104 ind·cm-3, 占总平均细胞密度的25.75%;蓝藻门为6.3×103 ind·cm-3, 占总平均细胞密度的14.48%;裸藻门为2.64×103 ind·cm-3, 占总平均细胞密度的5.98%.硅藻门密度最高出现在2016年为4.18×104 ind·cm-3, 最低值出现在2014年为6.12×103 ind·cm-3; 蓝藻门密度最高出现在2015年为1.22×104 ind·cm-3, 最低值出现在2014年为1.98×103 ind·cm-3.细胞密度变化整体呈先上升后下降的趋势, 其中2015~2017年密度处于较高水平, 各年间细胞密度差异性明显.

|
图 4 2014~2019年夏季松花江干流着生藻类细胞密度变化趋势 Fig. 4 Summer-time cell density of periphytic algae in the main stream of the Songhua River from 2014 to 2019 |
本文对2014~2019年年均细胞密度进行调查分析, 显示优势属主要由硅藻门、绿藻门和蓝藻门组成(表 2). 2014~2019年出现频率为100%的优势属为硅藻门小环藻属(Cyclotella)、直链藻属(Melosira)、星杆藻属(Asterionella)、桥弯藻属(Cymbella)、针杆藻属(Synedra)、羽纹藻属(Pinnularia)、舟形藻属(Navicula)和绿藻门的栅藻属(Scenedesmus).其中占绝对优势的小环藻属(Cyclotella)优势度由2014~2016年的0.436~0.513降至2017~2019年的0.110~0.300;直链藻属(Melosira)、星杆藻属(Asterionella)和舟形藻属(Navicula)优势度呈逐年下降趋势; 桥弯藻属(Cymbella)和针杆藻属(Synedra)呈先下降再上升的趋势; 羽纹藻属(Pinnularia)和栅藻属(Scenedesmus)呈先上升后下降的趋势.星杆藻属(Asterionella)在2019年优势度为0.099, 降幅较大; 栅藻属(Scenedesmus)在2017年优势度为0.335, 上升幅度较大; 其余藻类历年优势度变化不明显. 2015~2019年100%出现的优势属为硅藻门的平板藻属(Tabellaria)和脆杆藻属(Fragilaria), 绿藻门的小球藻属(Chlorella)、丝藻属(Ulothrix)和鼓藻属(Cosmarium), 蓝藻门的集胞藻属(Synechocystis), 5a的小球藻属(Chlorella)和丝藻属(Ulothrix)优势度降低, 其余藻类优势度变化幅度平缓. 2017~2019年蓝藻门的念珠藻属(Nostoc)和鱼腥藻属(Anabaena)的出现率分别为100%和66.67%, 优势度分别在0.051~0.023和0.021~0.037之间; 其余年际间交替出现不同种类的硅藻和绿藻, 个别年份如2018年出现过蓝藻门的细鞘丝藻属(Leptolyngbya)作为优势属, 优势度为0.030.优势属以淡水硅藻中指示轻-中污染的习见种类为主.
|
|
表 2 2014~2019年夏季松花江干流着生藻类优势属及优势度 Table 2 Dominant periphytic algal genera in the main stream of the Songhua River in summer from 2014 to 2019 |
2.4 松花江干流水质生物学评价
本文使用藻类植物群落的Shannon-Wiener多样性指数和Pielou均匀度指数对松花江干流点位监测的年平均评价结果值进行水质生物评价发现(表 1), 干流水体目前处于寡污型向β-中污型过渡的状态, 但不同的时间, 不同的点位仍有较大差异性如(图 5).其中Shannon-Wiener多样性指数均介于1~3之间, 属于β-中污型水质; Pielou均匀度指数介于0.5~0.8之间, 少部分高于0.8, 说明松花江干流处于清洁-寡污型的状态.

|
图 5 2014~2019年夏季松花江干流着生藻类多样性指数 Fig. 5 Average diversity index of periphytic algae in the main stream of the Songhua River in summer from 2014 to 2019 |
从点位来看, 松花江干流中上游点位多样性指数较低, 最低值出现在D3点位, 该点位多样性指数和均匀度指数均为最低, 总体而言着生藻类植物群落简单且不均匀, 以水质生物学评价该位点的水质, 属于β-中污型水体.而松花江干流下游点位多样性指数较上游高, 最高值出现在D9点位, 该点位多样性指数和均匀度指数均为最高, 说明该点着生藻类植物群落较复杂且分布较均匀, 水质生物性评价结果为β-中污型水体, 接近寡污型水体.
从不同时间来看, 2014~2016年多样性指数和均匀度指数均较高, 在2016年达到峰值. 2017~2019年多样性指数和均匀度指数开始下降, 且在2018年达到1个低值.多样性指数和均匀度指数均为年平均值, 6 a的总体变化趋势为先上升后下降.
2.5 着生藻类与环境因子关系针对松花江干流历年采样数据进行冗余分析(RDA), 各年度数据中第一轴物种与环境相关系数均大于0.93, 第二轴相关系数均大于0.87.各年度第1轴物种-环境解释率平均值为66.28%, 第2轴物种-环境解释率平均值为84.42%, 第3轴物种-环境解释率平均值为92.11%, 第4轴物种-环境解释率平均值为96.07% (表 3).
|
|
表 3 主要优势属与环境因子的RDA分析 Table 3 RDA analysis for dominant species and environment factors |
通过蒙特卡洛单因子检验法对理化影响因子进行检验, 将P < 0.05的环境因子提出并分析, 2014年主要环境影响因子为DO(P=0.002)、TN(P=0.023)、TP(P=0.018)、BOD(P=0.016)和NH4+-N(P=0.036); 2015年主要环境影响因子为DO(P=0.018)、TN(P=0.02)、TP(P=0.034)、BOD(P=0.046)和pH(P=0.023); 2016年主要环境影响因子为DO(P=0.016)、TP(P=0.026)、BOD(P=0.032)、NH4+-N(P=0.024)和COD(P=0.046); 2017年主要环境影响因子为DO(P=0.048)、pH(P=0.013)、TN(P=0.024)、TP(P=0.002)和NH4+-N(P=0.045); 2018年主要环境影响因子为DO(P=0.018)、COD(P=0.05)、BOD(P=0.046)、TN(P=0.006)和NH4+-N(P=0.037); 2019年主要环境影响因子为DO(P=0.032)、BOD(P=0.026)、WT(P=0.008), COD(P=0.032)和NH4+-N(P=0.047).从排序可知不同年份藻类植物群落与环境因子和空间因子之间的变化趋势(图 6), 全部的样品分布在主轴1和主轴2构成的4个象限内, 历年来11个采样点的DO始终与轴1呈显著正相关.其中2014年TN、NH4+-N和BOD与轴2负相关, TP与轴1负相关; 2015年TN、TP和BOD与轴1负相关, pH与轴2负相关; 2016年COD和TP与轴2正相关, NH4+-N和BOD与轴1负相关; 2017年TP和NH4+-N与轴1正相关, TN与轴2负相关, pH与轴1负相关; 2018年TN和NH4+-N与轴1正相关, COD与轴1负相关, BOD与轴2正相关; 2019年NH4+-N与轴1正相关, BOD和COD与轴1负相关, WT与轴2负相关.调查流域中上游河段的优势属主要是一些耐污种和广布种如小环藻属(Cyclotella)、直链藻属(Melosira)、星杆藻属(Asterionella)、针杆藻属(Synedra)、平板藻属(Tabellaria)、脆杆藻属(Fragilaria)、小球藻属(Chlorella)和舟形藻属(Navicula)等且随时间变化分别与含氮污染物和COD呈显著相关, 下游河段的优势属主要是一些寡污带指示种菱形藻属(Nitzschia)、异极藻属(Gomphonema)、舟形藻属(Navicula)和桥弯藻属(Cymbella)等随时间变化分别与DO和NH4+-N呈显著相关.在空间变化上, 不同采样点位之间营养程度不同样点D2、D3、D4、D5和D7较相似, D8、D9和D10较相似; D1和D11较相似, 而D6点位分别不同于其他点位. TN、NH4+-N和COD对D2、D3、D4、D5和D7贡献较大, NH4+-N和DO对D8、D9和D10贡献较大, TP和BOD对D1和D11贡献较大, NH4+-N、DO、BOD、COD、TP和pH对D6贡献较大.

|
缩写表示用于分析的主要藻类植物属种:Cy-cl表示Cyclotella, Me-lo表示Melosira, As-te表示Asterionella, Cy-mb表示Cymbella, Sy-ne表示Synedra, Pi-nn表示Pinnularia, Sc-en表示Scenedesmus, Na-vi表示Navicula, Co-sm表示Cosmarium, Ul-ot表示Ulothrix, Ch-lo表示Chlorella, Fr-ag表示Fragilaria, Ta-be表示Tabellaria, Sy-ne表示Synechocystis, Pe-di表示Pediastrum, Gy-ro表示Gyrosigma, An-ki表示Ankistrodesmus, Ni-tz表示Nitzschia, Su-ri表示Surirella, Co-cc表示Cocconeis, Go-mp表示Gomphonema, Te-tr表示Tetraspora, No-st表示Nostoc, Ha-nt表示Hantzschia, An-ab表示Anabaena, Le-pt表示Leptolyngbya, Eu-no表示Eunotia, Ac-hn表示Achnanthes, Rh-op表示Rhopalodia, Ac-ti表示Actinastrum 图 6 2014~2019年夏季松花江干流着生藻类与环境因子RDA分析 Fig. 6 RDA analysis of periphytic algae and environment factors in the main stream of the Songhua River in summer from 2014 to 2019 |
在自然水体中, 藻类的种类构成以硅藻和绿藻为主要优势种, 兼有少量的蓝藻、裸藻和隐藻等.以隐藻门为优势种的水体, 一般为贫-中营养; 以硅藻为优势种的水体, 一般为中-富营养; 而以蓝藻为优势种的水体, 一般为重富营养.当水体受到一定程度污染时, 藻类植物的群落组成会发生明显的变化, 转化为蓝-绿藻为主的群落[12].李欢等[22]在2014年夏季对长江中下游武汉-河口江段的浮游藻类空间分布特征进行调查, 共检出浮游藻类6门66种, 其中硅藻门24种和绿藻23种, 该流域主要由硅藻和绿藻组成.苟婷等[23]对珠江流域西红支流贺江浮游藻类群落特征及水质分析结果显示丰水期浮游藻类群落组成以绿藻门、硅藻门和蓝藻门为主, 其中以绿藻门居多, 硅藻门次之; 枯水期硅藻门最多.王宏伟等[24]在2010年9月到2011年9月, 4次对太子河流域的藻类植物群落组成、丰度及优势种进行调查分析, 藻类植物群落以硅藻最多, 占总体的43.77%;其次是绿藻, 占总体的35.56%, 蓝藻居第3位, 占总体的11.85%.笔者对2014~2019年夏季松花江干流着生藻类群落结构特征进行调查研究, 共鉴定出着生藻类4门、6纲、17目、26科和58属.其中硅藻门28属, 占总数48.28%;绿藻门17属, 占总数29.31%;蓝藻门10属, 占总数17.24%;裸藻门3属, 占总数5.17%.群落结构的组成类型为硅藻-绿藻型, 与其他大型河流藻类群落结构组成相似, 符合河流的群落分布特征.
本研究期间内流域着生藻类的群落结构组成发生了显著变化.陈威等[11]于2015年7~10月对松花江干流哈尔滨段进行了藻类的调查研究显示该流域藻类以硅藻门为主, 优势属为小环藻属(Cyclotella)、直链藻属(Melosira)、脆杆藻属(Fragilaria)和针杆藻属(Synedra)等寡污带-中污染带的指示种, 表明该江段水体受到污染但程度较轻.本研究调查发现2014~2016年干流着生藻类优势属由硅藻和绿藻组成, 其中人为活动干扰相对较多的上游采样点位优势属为小环藻属(Cyclotella)、直链藻属(Melosira)、小球藻属(Chlorella)、舟形藻属(Navicula)、脆杆藻属(Fragilaria)和针杆藻属(Synedra)等, 种类组成与陈威等调查结果相近, 但2017~2019年松花江干流哈尔滨段优势属类出现了蓝藻门中显示水体富营养化状态的念珠藻属(Nostoc)和鱼腥藻属(Anabaena)等.陈家厚等[25]对松花江干流哈尔滨段藻类植物的研究结果表明:大顶子山航电枢纽工程蓄水导致着生藻类的种类增加、生物密度增加和蓝绿藻比例增高.芦宴生[26]对松花江污染对浮游生物和着生藻类影响的初步研究结果显示, 1980~1982年松花江干流佳木斯江段着生藻类主要出现绿藻、裸藻、衣藻及硅藻门中的一些耐污种类, 寡污-清水型的星杆藻和黄丝藻等未见.但2014~2019年调查期间显示佳木斯江段硅藻占比由46%提高到78%, 每年都可检出作为主要优势属100%出现的星杆藻属(Asterionella).由藻类群落演替结果可知调查期间松花江干流下游水质好于松花江干流上游水质, 哈尔滨江段水质富营养化程度和污染程度都有一定增加, 佳木斯江段优势属类组成变化显示水质得到改善.
3.2 松花江干流水质生物学评价近年来, 生物多样性指数被广泛用于水质评价, 但有部分学者认为单纯的使用一种多样性指数来评价藻类群落的多样性易造成较大偏差, 因此本文将Shannon-Wiener指数和Pielou均匀度指数综合使用, 来反映松花江干流着生藻类群落结构个体分布情况、水体污染程度及变化趋势.松花江干流下游点位Shannon-Wiener多样性指数和Pielou均匀度指数值均较中上游高, 中上游水质类别为β中污型, D1~D7点位离城市比较近, 人类活动较多, 受干扰较大, 下游水质类别为清洁-寡污型, D8~D11点位远离城市, 人为干扰小, 植物覆盖率高, 水质较清洁. 2014~2019年水体总体呈寡污型-β中污型水质过度状态, 6 a的多样性指数呈现为先升高后下降的趋势, 其中2016年各指数达到顶峰, 其余年际间波动不大.藻类植物多样性的分布格局与密度及种类数分布相关, 2014~2019年细胞密度变化范围为1.29×104~8.42×104 ind·cm-3, 总平均细胞密度为4.35×104 ind·cm-3.最高值出现在2016年, 最低值出现在2014年, 6 a的细胞密度值呈先上升后下降趋势, 与多样性指数变化趋势相一致.陈威等[11]对松花江干流哈尔滨段的藻类植物进行研究调查, 采用指数评价法和相关性分析统计的方法评价该流域的水质状况和生物状态, 有研究表明Margalef和Shannon-Wiener等指数都可以很好地评价藻类植物的多样性, 总体评价为松花江哈尔滨段近人为干扰点水体为重污染, 远离市区和人为活动较少的点位水体为中污染.基于大型底栖动物对松花江干流2012~2015年的水生态环境质量分析结果显示[27, 28], 该流域水体总体呈良好-中度干扰的状态, 与本研究的着生藻类评价的结果相近.综上所述, 虽然随着松花江流域水污染控制与管理的加强, 流域整体水生态状态逐步恢复, 但部分江段如哈尔滨江段依然受污染威胁, 水质富营养化程度及污染程度有一定增加.
3.3 松花江干流着生藻类和环境因子的关系着生藻类对水体环境变化十分敏感, 其结构与水体环境具有直接的关系[29].本文通过冗余分析(RDA)探究着生藻类与环境因子间关系, 发现驱动松花江干流着生藻类群落结构演替的主要环境因子随时间推移而有所变化.相关性结果表明: DO、TN、NH4+-N、TP、COD和BOD为影响松花江干流藻类群落分布的重要因素, 与阴琨等[30]对松花江干流环境影响因子调查结果相一致.许多研究表明水质状况与着生藻类群落结构显著相关:Dalu等[31]的研究发现DO、浊度与着生藻类群落结构的变化具有显著的关系. DO是表征水体有机污染程度的重要性指标, 清洁水体DO趋向于饱和, 当水环境受到污染后DO浓度会大幅降低[32]. COD和BOD是评价水体污染的主要因子, COD和BOD多表征水体受到有机污染,可以说BOD降解量等于DO减少量, COD和DO没有直接联系.本研究发现松花江干流着生藻类与DO呈显著正相关, 与BOD和COD大体呈负相关趋势, 说明该流域着生藻类适宜在较高DO和较低COD和BOD的环境下生存, 且不同点位DO、BOD和COD的含量影响着生藻类群落结构, 使的上游和下游江段的藻类群落结构及其组成存在一定的差异性.李巧玉等[33]的研究结果表明, 变异直链藻和披针形舟形藻等与DO呈负相关, 文中调查显示上游点位人为干扰较大污染程度较重优势属为直链藻属(Melosira)、针杆藻属(Synedra)、平板藻属(Tabellaria)、脆杆藻属(Fragilaria)、小球藻属(Chlorella)和舟形藻属(Navicula)等与DO呈显著负相关, 与前人环境因子影响结果相一致.周贤杰等[34]的研究表明, 随着氮和磷等营养盐浓度的增加, 藻类植物种类由清洁型向耐污型转换, 细胞的密度、叶绿素浓度和比增长率随着氮和磷浓度的变化有着相同的增减趋势.本研究结果显示TN的变化浓度为1.87~2.17 mg·L-1, TP的变化浓度为0.1~0.14 mg·L-1, NH4+-N的变化浓度为0.25~0.57 mg·L-1, 由此可知松花江流域营养盐含量较高, 将在一定程度上影响藻类结构组成[35].杨宋琪等[36]的研究结果表明, 黑河张掖段藻类植物总密度与TN显著相关, 且TN浓度与优势种的分布呈正相关.本研究发现松花江干流中上游点位的优势属与TN呈正相关, 除2015年和2016年外下游点位的优势种与NH4+-N呈显著正相关. Pan等[37]的研究发现, 激流环境中的总磷对着生藻类多样性和物种丰富度有着重要的影响.总磷与着生藻类, 尤其与硅藻物种丰度和生物多样性有着密切联系[38].本研究发现处于松花江干流两端伴有支流汇入且流速较快的肇源和同江点位与TP呈显著正相关.一直以来松花江水系都有三大污染问题:有机污染、甲基汞污染和冰封期水质恶化, 存在的污染物主要是NH4+-N、TN和TP, 其来源主要为工业废水、生活污水、化肥农药、水土流失及流域内所释放的沉积物等.松花江作为黑龙江的母亲河, 部分流域作为哈尔滨市重要的饮用水水源地, 水污染防治迫在眉睫.需严格控制有机污染物和含磷生活污水排入松花江, 合理施肥; 提高人类对环境保护的意识, 控制并规范人们的行为; 加强重点流域环境管理和水环境的研究, 利用生物与理化相结合的手段进行长期的监控.
4 结论(1) 通过对2014~2019年夏季松花江干流着生藻类的群落结构特征及其对环境因子的响应关系分析, 对松花江干流的水生态环境质量状况及变化趋势进行探讨.
(2) 2014~2019年夏季松花江干流共鉴定着生藻类4门和58属.其中硅藻门28属、绿藻门17属、蓝藻门10属和裸藻门3属.出现的优势属大部分为轻-中污染指示属, 如小环藻属(Cyclotella)、直链藻属(Melosira)、针杆藻属(Synedra)、异极藻属(Gomphonema)、舟形藻属(Navicula)和小球藻属(Chlorella)等.
(3) 多样性指数评价松花江干流上游为中污染, 下游为轻污染状态; RDA结果显示, 驱动松花江干流着生藻类群落结构演替的主要环境因子随时间推移而有所变化, 并且因河段不同有所差异, 溶解氧、氨氮、总氮、总磷和生化需氧量等指标是这一区域着生藻类种类分布的重要环境影响因子.
| [1] | Malaj E, Von Der Ohe P C, Grote M, et al. Organic chemicals jeopardize the health of freshwater ecosystems on the continental scale[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(26): 9549-9554. DOI:10.1073/pnas.1321082111 |
| [2] | Török P, T-Krasznai E, B-Béres V, et al. Functional diversity supports the biomass-diversity humped-back relationship in phytoplankton assemblages[J]. Functional Ecology, 2016, 30(9): 1593-1602. DOI:10.1111/1365-2435.12631 |
| [3] | Chessman B, Growns I, Currey J, et al. Predicting diatom communities at the genus level for the rapid biological assessment of rivers[J]. Freshwater Biology, 1999, 41(2): 317-331. DOI:10.1046/j.1365-2427.1999.00433.x |
| [4] | Leland H V. Distribution of phytobenthos in the Yakima River basin, Washington, in relation to geology, land use and other environmental factors[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1995, 52(5): 1108-1129. DOI:10.1139/f95-108 |
| [5] | Round F E. Diatoms in river water-monitoring studies[J]. Journal of Applied Phycology, 1991, 3(2): 129-145. DOI:10.1007/BF00003695 |
| [6] | Tang T, Cai Q H, Liu J K. Using epilithic diatom communities to assess ecological condition of Xiangxi River system[J]. Environmental Monitoring and Assessment, 2006, 112(1-3): 347-361. DOI:10.1007/s10661-006-7666-6 |
| [7] | Martínez De Fabricius A L, Maidana N, Gómez N, et al. Distribution patterns of benthic diatoms in a pampean river exposed to seasonal floods: the Cuarto River (Argentina)[J]. Biodiversity & Conservation, 2003, 12(12): 2443-2454. |
| [8] | Uehlinger U, Kawecka B, Robinson C T. Effects of experimental floods on periphyton and stream metabolism below a high dam in the Swiss Alps (River Spl)[J]. Aquatic Sciences, 2003, 65(3): 199-209. DOI:10.1007/s00027-003-0664-7 |
| [9] | Luce J J, Cattaneo A, Lapointe M F. Spatial patterns in periphyton biomass after low-magnitude flow spates: geomorphic factors affecting patchiness across gravel-cobble riffles[J]. Journal of the North American Benthological Society, 2010, 29(2): 614-626. DOI:10.1899/09-059.1 |
| [10] |
金小伟, 王业耀, 王备新, 等. 我国流域水生态完整性评价方法构建[J]. 中国环境监测, 2017, 33(1): 75-81. Jin X W, Wang Y Y, Wang B X, et al. Methods development for monitoring and assessment of ecological integrity of surface waters in China[J]. Environmental Monitoring in China, 2017, 33(1): 75-81. |
| [11] |
陈威, 魏南, 金小伟, 等. 松花江哈尔滨段藻类植物分布及其与环境因子的关系[J]. 中国环境监测, 2018, 34(4): 102-110. Chen W, Wei N, Jin X W, et al. The phytoplankton assemblages and its relation to environmental factors in Harbin section of Songhua River, China[J]. Environmental Monitoring in China, 2018, 34(4): 102-110. |
| [12] |
王丑明, 郭晶, 张屹, 等. 1988—2017年洞庭湖浮游植物的群落演变[J]. 中国环境监测, 2018, 34(6): 19-25. Wang C M, Guo J, Zhang Y, et al. Phytoplankton community succession trends of the Dongting Lake from 1988 to 2017[J]. Environmental Monitoring in China, 2018, 34(6): 19-25. |
| [13] | Bao W M, Reimer C W. New taxa of the diatoms from Changbaishan Mountain, China[J]. Bulletin of Botanical Research, 1992, 12(4): 357-361. |
| [14] | Collins G B, Weber C I. Phycoperiphyton (Algae) as indicators of water quality[J]. Transactions of the American Microscopical Society, 1978, 97(1): 36-43. DOI:10.2307/3225682 |
| [15] | 胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [16] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [17] | Spatharis S, Roelke D L, Dimitrakopoulos P G, et al. Analyzing the (mis)behavior of Shannon index in eutrophication studies using field and simulated phytoplankton assemblages[J]. Ecological Indicators, 2011, 11(2): 697-703. DOI:10.1016/j.ecolind.2010.09.009 |
| [18] |
吴天浩, 刘劲松, 邓建明, 等. 大型过水性湖泊——洪泽湖浮游植物群落结构及其水质生物评价[J]. 湖泊科学, 2019, 31(2): 440-448. Wu T H, Liu J S, Deng J M, et al. Community structure of phytoplankton and bioassessment of water quality in a large water-carrying lake, Lake Hongze[J]. Journal of Lake Sciences, 2019, 31(2): 440-448. |
| [19] |
刘金殿, 顾志敏, 杨元杰, 等. 长诏水库浮游植物群落结构及水质评价[J]. 生态学杂志, 2012, 31(11): 2865-2871. Liu J D, Gu Z M, Yang Y J, et al. Phytoplankton community structure and water quality assessment of Changzhao Reservoir, Zhejiang of East China[J]. Chinese Journal of Ecology, 2012, 31(11): 2865-2871. |
| [20] |
贾晓妮, 程积民, 万惠娥, 等. DCA、CCA和DCCA三种排序方法在中国草地植被群落中的应用现状[J]. 中国农学通报, 2007, 23(12): 391-395. Jia X N, Cheng J M, Wan H E, et al. Application present situation of DCA、CCA、and DCCA ordination of grassland vegetation communities in China[J]. Chinese Agricultural Science Bulletin, 2007, 23(12): 391-395. DOI:10.3969/j.issn.1000-6850.2007.12.084 |
| [21] | Xu H, Paer H W, Qin B, et al. Determining critical nutrient thresholds needed to control harmful cyanobacterial blooms in eutrophic Lake Taihu, China[J]. Environmental Science & Technology, 2015, 49(2): 1051-1059. |
| [22] |
李欢, 赵伟华, 林莉, 等. 长江中下游干流浮游藻类空间分布特征初探[J]. 长江科学院院报, 2015, 32(6): 94-99. Li H, Zhao W H, Lin L, et al. Spatial distribution features of phytoplankton community in middle-low reaches of mainstream Yangtze River[J]. Journal of Yangtze River Scientific Research Institute, 2015, 32(6): 94-99. |
| [23] |
苟婷, 许振成, 李杰, 等. 珠江流域西江支流贺江浮游藻类群落特征及水质分析[J]. 湖泊科学, 2015, 27(3): 412-420. Gou T, Xu Z C, Li J, et al. Phytoplankton community structure and water quality assessment of Hejiang River, a branch of Xijiang River, Pearl River drainage basin[J]. Journal of Lake Sciences, 2015, 27(3): 412-420. |
| [24] |
王宏伟, 陈莹, 张晓明, 等. 辽河太子河流域藻类植物群落结构及其季节变化[J]. 湖泊科学, 2013, 25(6): 936-942. Wang H W, Chen Y, Zhang X M, et al. The algal community structure and seasonal variation in Taizi River catchment, Liaohe River basin[J]. Journal of Lake Sciences, 2013, 25(6): 936-942. |
| [25] |
陈家厚, 孙子孟, 白羽军, 等. 大顶子山航电枢纽工程蓄水后对松花江哈尔滨江段着生藻类的影响[J]. 环境科学与管理, 2010, 35(5): 46-50. Chen J H, Sun Z M, Bai Y J, et al. Influence of Dadingzishan closure on attached algae of Songhua River in Harbin Section[J]. Environmental Science and Management, 2010, 35(5): 46-50. DOI:10.3969/j.issn.1673-1212.2010.05.015 |
| [26] |
芦晏生. 松花江污染对浮游生物、着生藻类影响的初步研究[J]. 环境科学, 1985, 6(1): 33-39. Lu Y S. A preliminary study on the impact of Songhua River pollution on plankton and phytoplankton[J]. Environmental Science, 1985, 6(1): 33-39. |
| [27] |
李中宇, 金小伟, 刘继凤, 等. 应用底栖动物为指示生物的松花江干流水生态预警研究[J]. 生态毒理学报, 2017, 12(5): 161-169. Li Z Y, Jin X W, Liu J F, et al. Early warning for water ecosystem in main stream of Songhua River with benthic macroinvertebrate[J]. Asian Journal of Ecotoxicology, 2017, 12(5): 161-169. |
| [28] |
刘蕊, 李中宇, 赵然, 等. 中国北方河流底栖动物群落季节性演替对水生态环境质量评价的影响[J]. 中国环境监测, 2018, 34(6): 84-91. Liu R, Li Z Y, Zhao R, et al. Effects of benthic community seasonal dynamics on the aquatic environment quality assessment of rivers in northern China[J]. Environmental Monitoring in China, 2018, 34(6): 84-91. |
| [29] |
米文梅, 施军琼, 杨燕君, 等. 三峡库区支流梅溪河附石藻类群落变化及其与环境因子的关系[J]. 环境科学, 2020, 41(4): 1636-1647. Mi W M, Shi J Q, Yang Y J, et al. Changes in epilithic algae community and its relationship with environmental factors in the Meixi River, a Tributary of the Three Gorges Reservoir[J]. Environmental Science, 2020, 41(4): 1636-1647. |
| [30] |
阴琨, 赵然, 李中宇, 等. 松花江流域水生态环境中生物与生境和化学要素间的关联性研究[J]. 中国环境监测, 2015, 31(1): 17-23. Yin K, Zhao R, Li Z Y, et al. Study on the correlation between biological and habitat, chemical elements of water ecosystem quality in Songhua River basin[J]. Environmental Monitoring in China, 2015, 31(1): 17-23. DOI:10.3969/j.issn.1002-6002.2015.01.004 |
| [31] | Dalu T, Wasserman R J, Magoro M L, et al. Variation partitioning of benthic diatom community matrices: effects of multiple variables on benthic diatom communities in an Austral temperate river system[J]. Science of the Total Environment, 2017, 601-602: 73-82. DOI:10.1016/j.scitotenv.2017.05.162 |
| [32] |
俞焰, 刘德富, 杨正健, 等. 千岛湖溶解氧与浮游植物垂向分层特征及其影响因素[J]. 环境科学, 2017, 38(4): 1393-1401. Yu Y, Liu D F, Yang Z J, et al. Vertical stratification characteristics of dissolved oxygen and phytoplankton in Thousand-Island Lake and their influencing factors[J]. Environmental Science, 2017, 38(4): 1393-1402. |
| [33] |
李巧玉, 刘瑞, 向蓉, 等. 三峡库区支流底栖硅藻功能群特征及其驱动因子分析——以汝溪河为例[J]. 湖泊科学, 2017, 29(6): 1464-1472. Li Q Y, Liu R, Xiang R, et al. Functional group characteristics of benthic diatoms and its driving factors in the branch of Three Gorges Reservoir: a case study of Ruxi River[J]. Journal of Lake Sciences, 2017, 29(6): 1464-1472. |
| [34] |
周贤杰, 罗固源, 杨清玲, 等. 大宁河回水腹心区氮磷浓度对藻类生长影响的模拟试验研究[J]. 农业环境科学学报, 2008, 27(3): 1123-1127. Zhou X J, Luo G Y, Yang Q L, et al. Simulation study of effects of different nitrogen and phosphorus concentrations on algae growth in the central backwater reach of Daning River[J]. Journal of Agro-Environment Science, 2008, 27(3): 1123-1127. DOI:10.3321/j.issn:1672-2043.2008.03.049 |
| [35] | Dixit S S, Cumming B F, Birks H J B, et al. Diatom assemblages from Adirondack lakes (New York, USA) and the development of inference models for retrospective environmental assessment[J]. Journal of Paleolimnology, 1993, 8(1): 27-47. |
| [36] |
杨宋琪, 祖廷勋, 王怀斌, 等. 黑河张掖段浮游植物群落结构及其与环境因子的关系[J]. 湖泊科学, 2019, 31(1): 159-170. Yang S Q, Zu Y X, Wang H B, et al. Relationship between the structure of phytoplankton community and environmental factors in the Zhangye section of Heihe River[J]. Journal of Lake Sciences, 2019, 31(1): 159-170. |
| [37] | Pan Y D, Stevenson R J, Hill B H, et al. Using diatoms as indicators of ecological conditions in lotic systems: a regional assessment[J]. Journal of the North American Benthological Society, 1996, 15(4): 481-495. DOI:10.2307/1467800 |
| [38] | Cooper S R, Huvane J, Vaithiyanathan P, et al. Calibration of diatoms along a nutrient gradient in florida everglades water conservation area-2A, USA[J]. Journal of Paleolimnology, 1999, 22(4): 413-437. DOI:10.1023/A:1008049224045 |