 2020, Vol. 41
2020, Vol. 41

2. 湖南省灌溉水源水质污染净化工程技术研究中心, 长沙 410128;
3. 重金属污染耕地安全高效利用湖南省工程研究中心, 长沙 410128
 , ZHU Hu-cheng1 , PENG Ou1,2,3 , LI Dan-yang1,2,3 , YANG Rui-jia1,2,3 , PENG Jing1,2,3 , TIE Bai-qing1,2,3
, ZHU Hu-cheng1 , PENG Ou1,2,3 , LI Dan-yang1,2,3 , YANG Rui-jia1,2,3 , PENG Jing1,2,3 , TIE Bai-qing1,2,3
2. Engineering Research Center for Water Pollution Purification of Irrigation Source in Hunan Province, Changsha 410128, China;
3. Hunan Engineering Research Center for Safe and High-Efficient Utilization of Heavy Metal Pollution Farmland, Changsha 410128, China
土壤重金属污染已成为社会发展过程中亟待解决的问题, 因其具有不可降解、累积性和高毒性等特点[1], 进入土壤后会造成作物质量和产量下降, 同时通过食物链在生物体内累积, 对生物体造成不可逆损害[2, 3].人类的金属与类金属的生产与消费活动会造成土壤污染, 且我国的重金属污染大多为伴生性、复杂性和综合性的复合污染[4], Cd和As是常被发现在土壤中共存的Ⅰ致癌物[5], 文献[6]的数据显示, 全国土壤污染物点位超标率中Cd达到7.0%, 砷达到2.7%.“十三五”期间, “农业面源和重金属污染农田综合防治与修复技术研发”专项中设置了14个农田Cd和As污染防治领域项目, 将Cd和As的防治列为重要研究内容[7].刘春早等[8]对湘江流域土壤中重金属含量的研究结果表明, Cd和As总量分别在0.13~76.84 mg ·kg-1和4.25~549.67 mg ·kg-1之间.徐启胜等[9]对湖南省土壤中重金属含量调查发现, Cd和As含量分别超过背景值87.58倍和15.89倍.
微生物修复重金属污染土壤主要通过生物吸附、转化、矿化及溶解作用实现.已有研究表明微生物对重金属的修复作用, 如Bacillus megaterium、Pseudomonas stutzeri、Halomonas和Delftia等已被证明能够减少土壤中有效态Cd, 并减少作物各部位积累Cd[10~12]. Xu等[13]研究的抗砷菌Acidithiobacillus ferrooxidans DLC-5, 对去除低浓度As具有很大潜力; 共生的Halomonas sp.可促进Chlorella salina对As的吸收和富集, 增强对As污染水体的修复效果[14].段靖禹等[15]研究生物碳复合青霉菌可固定土壤中27%的As(Ⅳ).然而, 土壤中生存环境复杂, 重金属及其他污染物的毒害作用和土著微生物菌群的竞争作用等均会降低外源添加微生物的存活效率, 使之难以形成稳定的群落结构, 降低修复效果[16, 17].
微生物固定化技术发展于20世纪60年代后期, 已有研究表明生物炭、活性炭、棉纤维和海藻酸钙等都是微生物良好的载体[18~21].生物炭比表面积较大, 含有多种含氧官能团, 如羧基、酚羟基和羰基等, 能够固定重金属[22~24].生物炭的孔隙结构可减少细胞的分散性并提供更多的生存空间[25]; 生物炭中存在的C、O、K、N和P等元素可为微生物提供营养元素[26].目前, 微生物固定化技术在土壤中主要应用于有机污染物治理, 对于重金属污染治理研究相对较少.戚鑫等[27]利用生物炭固定微生物可显著降低土壤中可提取态U和Cd含量, 钝化效果最为显著; 张松等[28]利用生物炭固定化融合菌株F14提高该菌株对芘的去除能力, 去除率增加了58%.
本研究以玉米秸秆生物炭作为细菌固定化载体, 比较生物炭-细菌复合体与单一细菌及生物炭对镉和砷的吸附特性与去除效果, 并将其应用于土壤中, 探究对复合污染土壤中镉砷形态的影响, 以期为生物炭固定化细菌高效修复镉砷复合污染土壤的实际应用提供理论基础.
1 材料与方法 1.1 材料 1.1.1 供试生物炭玉米秸秆产自河南省郑州市, 用去离子水将玉米秸秆清洗、干燥并粉碎后过10目筛, 放入坩埚中并置于马弗炉内500℃限氧条件下热解2 h, 待冷却至室温后取出, 用0.1 mol ·L-1的HCl洗涤至洗液呈中性后烘干, 磨碎, 过100目筛得到玉米秸秆生物炭(CSB).
1.1.2 菌体制备本研究所用菌株为Delftia sp. B9(B9), 基因登录号:MF679148, 菌种保藏号:CGMCC No.16073.
液体培养基:蛋白胨10 g、牛肉膏3 g和氯化钠5 g用蒸馏水定容至1 L, 调节pH 7.2~7.4.121℃高压灭菌30 min.
菌体的制备:将B9接种在液体培养基中30℃、120 r ·min-1振荡培养24 h, 用无菌水洗涤两次后制成D600=1.6(7.6×108 cfu ·mL-1)的菌悬液.
菌悬液在5 000 r ·min-1离心后弃上清液, 菌体用0.85%生理盐水稀释后再离心, 如此反复3次, 以去除培养基成分.最后一次离心的菌体用0.85%生理盐水制成菌悬液, 备用.同时取混匀后菌悬液离心烘干, 测定细菌干重.
1.1.3 生物炭-细菌复合体制备复合体制备:生物炭和菌悬液以固液比1:4(干重比约为25:1)的比例混合于锥形瓶中, 在30℃, 120 r ·min-1条件下振荡2 h, 制得复合体悬浮液(B-CSB).
1.1.4 吸附材料结构表征B9、CSB和B-CSB这3种吸附材料表面特征测定:样品形貌特征用场发射扫描电子显微镜(SEM, QUANTA430, FEI公司, 美国); 吸附前后材料分析采用傅里叶变换红外光谱仪(FT-IR Nixolet iS50, ThermoFisher科技有限公司, 美国)和X射线光电子能谱仪(XPS ESCALAB 250Xi, ThermoFisher科技有限公司, 美国).
1.1.5 供试土壤供试土壤采自湖南省株洲市, 表 1为理化性质.
|
|
表 1 土壤理化性质 Table 1 Physical and chemical properties of soil |
1.2 B9、CSB和B-CSB对Cd和As吸附实验 1.2.1 等温吸附实验
设置吸附体系中B9、CSB和B-CSB的投加量均为0.1 g ·L-1, 准确设置Cd2+/As(Ⅴ)质量浓度梯度为0、1、5、10、20和50 mg ·L-1(每个质量浓度梯度设置3个平行样), 用0.1 mol ·L-1 HNO3/NaOH溶液调节吸附体系pH为6.0±0.1, 在25℃, 120 r ·min-1振荡12 h, 振荡结束后在5 000 r ·min-1的条件下离心10 min, 上清液用0.45 μm水系滤膜过滤, 滤液Cd2+质量浓度采用电感耦合等离子体发射光谱(ICP Optima 8300, PE公司, 美国)测定, As(Ⅴ)质量浓度采用原子荧光光度计(BAF-2000, 宝德仪器有限公司, 中国)测定.
1.2.2 pH对吸附效果的影响吸附体系中B9、CSB和B-CSB的投加量均为0.1 g ·L-1, Cd2+/As(Ⅴ)初始质量浓度为10 mg ·L-1, 吸附体系pH分别调节为3、4、5、6、7、8和9(用0.1 mol ·L-1 HNO3/NaOH溶液调节, 每个pH梯度设置3个平行样), 在25℃, 120 r ·min-1振荡12 h, 振荡结束后在5 000 r ·min-1的条件下离心10 min, 上清液用0.45 μm水系滤膜过滤, 测定方法同1.2.1节.
1.3 B9、CSB和B-CSB对土壤中Cd和As形态的影响 1.3.1 土壤培养实验本实验用土经阳光暴晒、晾干后过2 mm筛, 每个烧杯中装200 g干土, 设置4个处理, 每个处理3个平行, 各处理见表 2.分别于培养后0、5、10、20、30和60 d取鲜土样20.0 g, 风干后研磨过100目筛, 测定镉和砷各形态含量.
|
|
表 2 土壤培养实验 Table 2 Soil culture experiment |
1.3.2 分析测定方法
Cd形态分析采用优化BCR法[29], 分为弱酸可溶态Cd(包括可交换态和碳酸盐结合态)、可还原态Cd(铁锰氧化物结合态)、可氧化态Cd(包括有机物结合态、硫化物结合态)和残渣态Cd.砷形态分析采用分级测定的方法[30], 分为交换态砷(AE-As)、Al型砷(Al-As)、Fe型砷(Fe-As)、Ca型砷(Ca-As)和残渣态砷(Res-As).所有样品处理过程中同时添加试剂空白、质控样.镉砷各形态含量测定方法同1.2.1节.
1.4 数据分析B9、CSB和B-CSB对Cd和As吸附效果通过对Cd和As吸附量来衡量.其计算公式如式(1)所示.
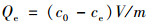
|
(1) |
式中, c0为溶液初始浓度, mg ·L-1; ce为溶液吸附平衡时浓度, mg ·L-1; V为溶液体积, mL; m为吸附剂质量, g.
用Langmuir模型对等温吸附实验数据进行分析拟合.Langmuir模型属单分子层吸附模型如式(2)所示.

|
(2) |
式中, qmax为吸附剂的单层饱和最大吸附量, mg ·g-1; ce为吸附平衡时溶液浓度, mg ·L-1; KL为Langmuir吸附常数, L ·g-1, 当0<KL<1, 表明有利于吸附; KL>1, 表示不利于吸附; KL=1, 属于线性分配; KL趋于0, 表示不可逆吸附.
2 结果与分析 2.1 吸附材料微观形貌及对Cd和As吸附 2.1.1 表征分析通过SEM可直观地反映吸附材料之间的形态变化.图 1(a)显示CSB表面粗糙, 有孔隙均匀分布.如图 1(b)所示, B9菌呈杆状, 长约1~2 μm, 宽约0.4 μm.图 1(c)显示CSB表面及孔隙内附着大量的B9, 说明B9与CSB复合成功.

|
(a)CSB; (b)B9; (c)B-CSB 图 1 CSB、B9和B-CSB的SEM图 Fig. 1 SEM images of CSB, B9, and B-CSB |
表 3是B9、CSB和B-CSB对Cd2+/As(Ⅴ)等温吸附模型的拟合参数; 根据式(2)拟合的Langmuir模型如图 2所示.通过分析B9、CSB和B-CSB的拟合度R2可知, 3种材料的吸附Cd2+/As(Ⅴ)过程符合Langmuir模型, 可推测B9、CSB和B-CSB表面的吸附位点有限, 吸附机制属于单分子层吸附[31].B9、CSB和B-CSB这3种材料对Cd2+的最大吸附量分别为49.43、82.68和75.38 mg ·g-1; 3种材料对As(Ⅴ)的最大吸附量分别为24.67、42.92和34.03 mg ·g-1(表 3).

|
图 2 B9、CSB和B-CSB对Cd和As吸附等温线 Fig. 2 B9, CSB, and B-CSB on the Cd and As adsorption isotherm |
|
|
表 3 Cd2+/As(Ⅴ)等温吸附模型拟合参数 Table 3 Fitness of isotherm models and corresponding parameters of Cd2+ and As(Ⅴ) |
2.1.3 pH对吸附效果的影响
由图 3可知, 在Cd2+初始质量浓度为10 mg ·L-1时, B9、CSB和B-CSB对Cd2+吸附量随溶液pH的增大而增加.当pH>5时, B-CSB吸附量趋近平稳, 吸附量约为70 mg ·g-1.当pH为3~7时, B9和CSB的吸附量明显增加, 当pH>7时, 吸附量缓慢增加, B9最大吸附量约为60 mg ·g-1, CSB的吸附量约为80 mg ·g-1.

|
图 3 pH对B9、CSB和B-CSB吸附Cd2+的影响 Fig. 3 Effects of pH on the adsorption of Cd2+ by B9, CSB, and B-CSB |
由图 4可知, 当pH为3~6时, B9、CSB和B-CSB对溶液中As(Ⅴ)的吸附量略有增加, 但增加量不显著; pH=6时, B9、CSB和B-CSB的吸附量达到最大值, 分别为9.79、19.30和18.65 mg ·g-1; 当pH为7~9时, 3种材料对As(Ⅴ)的吸附量随着pH的增加明显减少.

|
图 4 pH对B9、CSB和B-CSB吸附As(Ⅴ)的影响 Fig. 4 Effects of pH on the adsorption of As(Ⅴ) by B9, CSB, and B-CSB |
傅里叶红外光谱可检测材料中所含化合物的化学结构, 包括各种官能团, 如氨基、羧基和羰基等.B9、CSB和B-CSB吸附重金属离子前后的FTIR谱图如图 5所示.B9[图 5(a)]细胞壁表面上基团主要有:3 291.6 cm-1来自蛋白质的C—H键、N—H键的伸缩振动以及碳水化合物中结合水的O—H键伸展振动; 2 931.5 cm-1处为N—H键的伸缩振动; 1 656.7、1 536.3和1 390.8 cm-1处的谱峰分别为蛋白质酰胺Ⅰ带(C O的伸展振动)、酰胺Ⅱ带(N—H键的弯曲振动与C—N键的伸展振动)和酰胺Ⅲ带; 1 234.6 cm-1处的峰主要为P O和C S的伸缩振动及C O和O—H的叠加; 1 074.8 cm-1处的峰为胺基中C—N的伸缩振动、磷酸酯的振动[32].当B9吸附Cd2+后, 3 291.6、1 656.7、1 536.3和1 390.8 cm-1处的来自蛋白质的C—H键、N—H键以及酰胺Ⅰ、Ⅱ、Ⅲ带的峰强均明显增加, 表明蛋白质上的官能团对B9吸附Cd2+起重要作用.当B9吸附As(Ⅴ)后, B9细胞壁上的基团峰强均明显增加, 并且强度大于B9对Cd2+吸附.B9吸附Cd2+和As(Ⅴ)后官能团增加表明金属离子促进菌株生长和产生更多的活性基团.

|
图 5 B9、CSB和B-CSB吸附Cd2+和As(Ⅴ)后的FTIR图谱 Fig. 5 FTIR spectrograms of B9, CSB, and B-CSB before and after the absorption of Cd2+ and As(Ⅴ) |
CSB[图 5(b)]主要特征吸收峰出现在3 327.8、1 694.2、1 586.1、1 095.5、887.0和812.1 cm-1波数位置, 其中3 327.8 cm-1处吸收峰是由CSB表面的C—H键的伸缩振动; 1 694.2 cm-1处吸收峰为芳香酸类—COOH; 1 586.1 cm-1处吸收峰是由—COOH、—OH和内酯基的C—O和C C的伸缩振动; 1 095.5 cm-1处为醚键R—O—R′; 887.0 cm-1和812.1 cm-1处均为吡啶、吲哚等芳香化和杂环化合物的不饱和烃基C—H弯曲振动[33].当CSB吸附Cd2+后, 3 327.8、1 586.1、1 095.5、887.0和812.1 cm-1处峰强均明显减弱, 表明CSB的C—H、—COOH、—OH、C—O、C C和R—O—R′与Cd2+发生配位络合、离子交换, 含氧官能团、芳香类化合物提供π电子与Cd2+形成稳定结构.当CSB吸附As(Ⅴ)后, CSB的官能团峰强均明显增加, —COOH、—OH、—C—O、—C C等的变化表明As(Ⅴ)与CSB表面发生羟基络合作用及与生物炭表面芳香族化合物发生亲电取代反应.
B9与CSB复合[B-CSB, 图 5(c)]后大部分基团波峰变宽, 与B9相比, 1 656.7 cm-1处酰胺Ⅰ带的C O峰消失, 3 291.6、2 931.5、1 390.8和1 234.6 cm-1处峰强均减弱, 1 536.3 cm-1处酰胺Ⅱ带峰宽峰强增强.与CSB相比, 增加了3 291.7 cm-1处的O—H, 1 694.2 cm-1处芳香类—COOH峰消失, 其他处峰强有所增强.B-CSB吸附Cd2+和As(Ⅴ)后, 3 291.7、2 924.4、1 231.0和1 086.7 cm-1处的O—H、N—H、P O、C S和C O等峰强均增加, 均增加1 637.6 cm-1处的酰胺Ⅰ带C O的伸展振动峰.B-CSB吸附As(Ⅴ)后, 800.5 cm-1处峰强增加, Hu等[34]的研究表明, 在650~950 cm-1间出现As—O伸缩振动带.
2.2.2 X射线光电子能谱(XPS)分析通过对B9、CSB和B-CSB吸附Cd2+和As(Ⅴ)前后的表面元素结合能的变化来进一步研究3种吸附材料对Cd2+和As(Ⅴ)的吸附机制(图 6). 3种吸附材料的O 1s/C 1s:CSB>B-CSB>B9, B9、B-CSB吸附Cd2+和As(Ⅴ)后, O 1s/C 1s均增加, 表明吸附Cd2+和As(Ⅴ)后芳香化程度增强, 含氧官能团增多, 但含氧官能团数量减少.CSB吸附As(Ⅴ)后, O 1s/C 1s降低, O 1s含量减少, C 1s含量没有明显变化, 表明含氧官能团在吸附As(Ⅴ)时起作用.因此, 利用XPS Peak软件对3种吸附材料的O 1s峰面积及RWA值进行了分析.

|
图 6 B9、CSB和B-CSB吸附Cd2+和As(Ⅴ)后的XPS图谱 Fig. 6 XPS spectrograms of B9, CSB, and B-CSB before and after the absorption of Cd2+ and As(Ⅴ) |
图 7是B9、CSB和B-CSB吸附Cd2+和As(Ⅴ)前后O 1s谱图.B9吸附Cd2+和As(Ⅴ)前后峰值分别在531.29、531.52和531.53 eV处可归类为C O/P O, 峰值分别在532.58、532.80和532.74 eV处可归类为C—O—C/C—O—H. B9吸附Cd2+和As(Ⅴ)后C O/P O的峰面积从37.88%分别增加到41.63%和52.99%; C—O—C/C—O—H的峰面积从62.12%分别减少到58.37%、47.01%.CSB吸附Cd2+和As(Ⅴ)前后峰值在531.60、531.45和531.49 eV处可归类为C O, 峰值在533.30、533.32和533.18 eV处可归类为C—O—C/C—O—H.CSB吸附Cd2+后C O和C—O—C/C—O—H的峰面积未发生明显改变; 吸附As(Ⅴ)后, C O的峰面积从23.72%减少到19.60%, C—O—C/C—O—H的峰面积从76.28%增加到80.4%.B-CSB吸附Cd2+和As(Ⅴ)前后峰值在531.39、531.9和531.39 eV处可归类为C O/P O, 峰值在532.61、533.14和532.80 eV处可归类为C—O—C/C—O—H.B-CSB吸附Cd2+和As(Ⅴ)后C O/P O的峰面积从22.17%分别增加到47.67%和29.16%; C—O—C/C—O—H的峰面积从77.83%分别减少到52.33%和70.84%.

|
图 7 氧元素核心轨道X射线光电子能谱图 Fig. 7 Oxygen element core orbit XPS image |
土壤培养60 d后镉形态变化如图 8所示, 与CK相比, B9(T1)、CSB(T2)和B-CSB(T3)的添加均能降低土壤中弱酸可溶态Cd含量, 可还原态和残渣态Cd含量增加, 可氧化态Cd含量变化不明显.在培养60 d后, T1、T2和T3处理中弱酸可溶态Cd含量降至最小值, 分别减少了8.85%、10.14%和13.36%, 并且减少量是T3>T2>T1;可还原态Cd含量增加明显, 培养至第60 d时分别达到9.85%、10.08%和9.55%, 比初始培养时分别增加了2.04%、2.27%和1.74%;残渣态Cd含量明显增加, 培养至60 d时分别增加了5.86%、6.37%和10.35%.

|
图 8 B9、CSB和B-CSB对复合污染土壤中镉形态的影响 Fig. 8 Effects of the addition of B9, CSB, and B-CSB on the cadmium form in cadmium-arsenic complex contaminated soil |
土壤培养60 d后, 砷形态变化如图 9所示, 与CK相比, 添加3种材料后均能减少土壤中AE-As、Al-As和Fe-As含量, 增加Ca-As和Res-As含量.培养60 d后, T1、T2和T3处理中Fe-As含量与初始相比分别减少了1.56%、2.21%和2.86%, 并且减少量是T3>T2>T1;土壤中AE-As的降幅为0.90% ~1.04%; Al-As的降幅为1.57% ~2.91%.T1、T2和T3处理中Ca-As含量分别增加5.89%、6.27%和7.19%; Res-As含量分别增加3.90%、4.33%和5.35%, 且增加量是T3>T2>T1.培养60 d后CK中各形态砷含量变化不明显.

|
图 9 B9、CSB和B-CSB对复合污染土壤中As形态的影响 Fig. 9 Effects of the addition of B9, CSB, and B-CSB on the form of arsenic in cadmium-arsenic complex contaminated soil |
Delftia sp. B9可以在Cd2+浓度为300 mg ·L-1, As(Ⅴ)浓度为200 mg ·L-1的条件下生长良好, 且对Cd2+和As(Ⅴ)的理论最大吸附量分别达到49.43 mg ·g-1和24.67 mg ·g-1(表 3), 表明B9对Cd2+和As(Ⅴ)均具有较好地吸附效果.B9在土壤修复实际应用中[12], 外源添加到污染土壤后, 对早稻(较短时间)土壤镉形态影响较小, 对晚稻(较长时间)土壤形态影响增大, 这可能是实际应用中土壤及外界环境复杂, 威胁外源添加细菌的生长和存活, 限制细菌发挥作用.生物炭作为细菌固定化的载体修复重金属污染的研究还较少, 生物炭固定细菌后可提供营养元素及生存空间, 增加细菌的团聚性; 生物炭可以有效吸附Cd和As(表 3), 在一定程度上缓解Cd和As对细菌的毒害作用, 增加细菌的存活效率, 同时可为细菌提供更多接触金属离子的机会; 生物炭与细菌之间的电子转移在Cd和As形态转化中起到重要作用, 复合后有增加吸附重金属的潜力[35].同时, 微生物对生物炭的分解速度很慢, 且分解发生在生物炭表面, 表面以下的生物炭会维持稳定的O/C比, 保持生物炭稳定性[36].本实验研究显示, 细菌与生物炭复合(B-CSB)后对Cd2+和As(Ⅴ)理论最大吸附量均增加, 分别达到75.38 mg ·g-1和34.03 mg ·g-1(图 2和表 3), 与B9相比吸附量均增加, 但与CSB相比吸附量却有所减少, 这可能是由于细菌与生物炭复合后占据了部分生物炭上的官能团或者阻隔了生物炭官能团与重金属离子接触[37].
重金属总量可以评价该土壤污染水平, 但不能准确反映该金属元素的生物有效性与生态风险大小, 金属元素的不同存在形态环境行为和生态效应不同, 故分析重金属不同赋存形态具有重要意义[38].土壤中的弱酸可溶态Cd易直接被植物吸收; 残渣态Cd主要赋存于原生、次生矿物与硅酸盐等土壤晶格中, 几乎不能被生物利用[39].土壤交换态砷(AE-As)易迁移, 生物有效性高, 而Al-As、Fe-As、Ca-As和Res-As统称为难溶性砷[40].土壤培养实验结果表明, 经过60 d的培养, 添加B9、CSB及B-CSB后土壤中镉砷形态均发生一定变化(图 8和9), 对于B9和CSB对土壤中镉形态的影响原理已有过探讨[41, 42].土壤中的砷主要以阴离子形式存在, 对铁锰氧化物具有较强的亲和力, 而生物炭有较强的阳离子交换能力, 可以与土壤中的Fe和Mn结合, 形成铁锰氧化物, 从而达到吸附及转化砷的效果[15, 43].B9细胞表面的羟基可被As(Ⅴ)取代或络合, 从而吸附As(Ⅴ)[44]; 细菌由于胞内外As(Ⅴ)浓度差, 也会产生主动吸附将As(Ⅴ)转移至细胞内[45]; Delftia sp. B9有较高的产铁载体的能力[46], 铁载体可以与多种重金属离子进行络合, 形成稳定的络合物, 降低重金属的浓度, Nair等[47]报道铁载体可与镉、镍、铅、钴和砷等络合形成稳定的络合物, 铁载体中的官能团可通过多价阳离子桥接作用或者氨基质子化作用与砷酸根离子螯合[48].本研究中有效态砷、铝型砷和铁型砷含量减少, 钙型砷和残渣态砷含量增加.随着培养时间的延长, B-CSB对土壤中镉砷形态的影响大于B9和CSB, 这可能是由于细菌与生物炭复合后, 生物炭为细菌提供更加适宜的生存空间, Delftia sp. B9生物量比单一添加细菌增加得更多[49], 故增加了对镉砷形态的影响.
4 结论(1) 生物炭-细菌复合体(B-CSB)增加了Delftia sp. B9对Cd2+和As(Ⅴ)的吸附能力, Cd2+和As(Ⅴ)的吸附量分别由49.43 mg ·g-1和24.67 mg ·g-1增加至75.38 mg ·g-1和34.03 mg ·g-1.
(2) 在Cd2+/As(Ⅴ)初始质量浓度为10 mg ·L-1时, 3种吸附材料吸附Cd2+的量随着pH的增加而增加, 吸附As(Ⅴ)的量随着pH的增加而减少.
(3) 生物炭-细菌复合体(B-CSB)对土壤中镉砷形态影响更大, 能增加Cd从弱酸可溶态向可还原态和残渣态转化, 增加As从铁型砷向残渣态砷转化.
| [1] |
张静静, 朱爽阁, 朱利楠, 等. 不同钝化剂对微碱性土壤镉, 镍形态及小麦吸收的影响[J]. 环境科学, 2020, 41(1): 460-468. Zhang J J, Zhu S G, Zhu L N, et al. Effects of different amendments on fractions and uptake by winter wheat in slightly alkaline soil contaminated by cadmium and nickel[J]. Environmental Science, 2020, 41(1): 460-468. |
| [2] | Rahman M F, Ghosal A, Alam M F, et al. Remediation of cadmium toxicity in field peas (Pisum sativum L.) through exogenous silicon[J]. Ecotoxicology and Environmental Safety, 2017, 135: 165-172. |
| [3] | O'Connor D, Peng T Y, Zhang J L, et al. Biochar application for the remediation of heavy metal polluted land:A review of in situ field trials[J]. Science of the Total Environment, 2018, 619-620: 815-826. |
| [4] |
张继舟, 王宏韬, 倪红伟, 等. 我国农田土壤重金属污染现状、成因与诊断方法分析[J]. 土壤与作物, 2012, 1(4): 212-218. Zhang J Z, Wang H T, Ni H W, et al. Current situation, sources and diagnosis method analysis of heavy metal contamination in agricultural soils[J]. Soil and Crop, 2012, 1(4): 212-218. |
| [5] | Diacomanolis V, Noller B N, Ng J C. Bioavailability and pharmacokinetics of arsenic are influenced by the presence of cadmium[J]. Chemosphere, 2014, 112: 203-209. |
| [6] | 环境保护部, 国土资源部. 全国土壤污染状况调查公报[J]. 中国环保产业, 2014(5): 10-11. |
| [7] |
徐长春, 郑戈, 林友华. "十三五"国家重点研发计划农田镉砷污染防治领域资助情况概述[J]. 农业环境科学学报, 2018, 37(7): 1321-1325. Xu C C, Zheng G, Lin Y H. Brief introduction to research projects on prevention and control of cadmium and arsenic pollution in crop-lands supported by National Key R&D Program of China in 13th Five-Year Period[J]. Journal of Agro-Environment Science, 2018, 37(7): 1321-1325. |
| [8] |
刘春早, 黄益宗, 雷鸣, 等. 湘江流域土壤重金属污染及其生态环境风险评价[J]. 环境科学, 2012, 33(1): 260-265. Liu C Z, Huang Y Z, Lei M, et al. Soil contamination and assessment of heavy metals of Xiangjiang River basin[J]. Environmental Science, 2012, 33(1): 260-265. |
| [9] |
徐启胜, 李雨晴, 陈燕, 等. 我国中南地区土壤重金属污染状况及其空间分布研究[J]. 北方农业学报, 2018, 46(3): 93-100. Xu Q S, Li Y Q, Chen Y, et al. Study on heavy metal pollution and its spatial distribution in soil in central south China[J]. Journal of Northern Agriculture, 2018, 46(3): 93-100. |
| [10] | Li Y, Pang H D, He L Y, et al. Cd immobilization and reduced tissue Cd accumulation of rice (Oryza sativa wuyun-23) in the presence of heavy metal-resistant bacteria[J]. Ecotoxicology and Environmental Safety, 2017, 138: 56-63. |
| [11] | Siripornadulsil S, Siripornadulsil W. Cadmium-tolerant bacteria reduce the uptake of cadmium in rice:potential for microbial bioremediation[J]. Ecotoxicology and Environmental Safety, 2013, 94: 94-103. |
| [12] | Liu Y L, Tie B Q, Li Y X, et al. Inoculation of soil with cadmium-resistant bacterium Delftia sp. B9 reduces cadmium accumulation in rice (Oryza sativa L.) grains[J]. Ecotoxicology and Environmental Safety, 2018, 163: 223-229. |
| [13] | Xu S H, Xu R X, Nan Z R, et al. Bioadsorption of arsenic from aqueous solution by the extremophilic bacterium Acidithiobacillus ferrooxidans DLC-5[J]. Biocatalysis and Biotransformation, 2018, 37(1): 35-43. |
| [14] |
许平平, 刘聪, 王亚, 等. 共生细菌对盐生小球藻富集和转化砷酸盐的影响[J]. 环境科学, 2016, 37(9): 3438-3446. Xu P P, Liu C, Wang Y, et al. Effects of a symbiotic bacterium on the accumulation and transformation of arsenate by Chlorella salina[J]. Environmental Science, 2016, 37(9): 3438-3446. |
| [15] |
段靖禹, 李华, 马学文, 等. 青霉菌与生物炭复合修复土壤砷污染的研究[J]. 环境科学学报, 2019, 39(6): 1999-2005. Duan J Y, Li H, Ma X W, et al. Study on the remediation of arsenic pollution in soil by the combination of penicillium and biochar[J]. Acta Scientiae Circumstantiae, 2019, 39(6): 1999-2005. |
| [16] | Li X Q, Meng D L, Li J, et al. Response of soil microbial communities and microbial interactions to long-term heavy metal contamination[J]. Environmental Pollution, 2017, 231: 908-917. |
| [17] | Igalavithana A D, Lee S E, Lee Y H, et al. Heavy metal immobilization and microbial community abundance by vegetable waste and pine cone biochar of agricultural soils[J]. Chemosphere, 2017, 174: 593-603. |
| [18] | Ma X S, Li N J, Jiang J, et al. Adsorption-synergic biodegradation of high-concentrated phenolic water by Pseudomonas putida immobilized on activated carbon fiber[J]. Journal of Environmental Chemical Engineering, 2013, 1(3): 466-472. |
| [19] | de-Bashan L E D, Bashan Y. Immobilized microalgae for removing pollutants:review of practical aspects[J]. Bioresource Technology, 2010, 101(6): 1611-1627. |
| [20] | Bergero M F, Liffourrena A S, Opizzo B A, et al. Immobilization of a microbial consortium on Ca-alginate enhances degradation of cationic surfactants in flasks and bioreactor[J]. International Biodeterioration & Biodegradation, 2017, 117: 39-44. |
| [21] | Lonappan L, Liu Y X, Rouissi T, et al. Covalent immobilization of laccase on citric acid functionalized micro-biochars derived from different feedstock and removal of diclofenac[J]. Chemical Engineering Journal, 2018, 351: 985-994. |
| [22] | Shaheen S M, Niazi N K, Hassan N E E, et al. Wood-based biochar for the removal of potentially toxic elements in water and wastewater:a critical review[J]. International Materials Reviews, 2018, 64(4): 216-247. |
| [23] | Archanjo B S, Mendoza M E, Albu M, et al. Nanoscale analyses of the surface structure and composition of biochars extracted from field trials or after co-composting using advanced analytical electron microscopy[J]. Geoderma, 2017, 294: 70-79. |
| [24] |
梁婷, 李莲芳, 朱昌雄, 等. 铈锰改性生物炭对土壤As的固定效应[J]. 环境科学, 2019, 40(11): 5114-5123. Liang T, Li L F, Zhu C X, et al. Cerium-manganese modified biochar immobilizes arsenic in farmland soils[J]. Environmental Science, 2019, 40(11): 5114-5123. |
| [25] | Chen B Y, Chen C Y, Guo W Q, et al. Fixed-bed biosorption of cadmium using immobilized Scenedesmus obliquus CNW-N cells on loofa (Luffa cylindrica) sponge[J]. Bioresource Technology, 2014, 160: 175-181. |
| [26] | Gul S, Whalen J K, Thomas B W, et al. Physico-chemical properties and microbial responses in biochar-amended soils:Mechanisms and future directions[J]. Agriculture, Ecosystems & Environment, 2015, 206: 46-59. |
| [27] |
戚鑫, 陈晓明, 肖诗琦, 等. 生物炭固定化微生物对U、Cd污染土壤的原位钝化修复[J]. 农业环境科学学报, 2018, 37(8): 1683-1689. Qi X, Chen X M, Xiao S Q, et al. In situ remediation of U-and Cd-contaminated soils by immobilized microorganisms and biochar[J]. Journal of Agro-Environment Science, 2018, 37(8): 1683-1689. |
| [28] |
张松, 侯彬, 纪婷婷, 等. 生物质炭固定化融合菌株F14方法的研究及其对芘的去除[J]. 农业环境科学学报, 2018, 37(3): 464-470. Zhang S, Hou B, Ji T T, et al. Degradation of pyrene by immobilized cells of fusant bacterial strain F14 using biochar[J]. Journal of Agro-Environment Science, 2018, 37(3): 464-470. |
| [29] |
张永利, 刘晓文, 陈启敏, 等. Tessier法和改进BCR法提取施加熟污泥后黄土中Cd的对比研究[J]. 环境工程, 2019, 37(5): 34-38, 81. Zhang Y L, Liu X W, Chen Q M, et al. Comparative study of tessier method and modified BCR method for extracting Cd in loess amended by composted sludge[J]. Environmental Engineering, 2019, 37(5): 34-38, 81. |
| [30] | 刘凤枝. 农业环境监测实用手册[M]. 北京: 中国标准出版社, 2001: 97-107. |
| [31] |
李丹阳, 杨蕊嘉, 罗海艳, 等. 十六烷基三甲基溴化铵改性生物炭对水中镉离子吸附性能的影响[J]. 环境工程学报, 2019, 13(8): 1809-1821. Li D Y, Yang R J, Luo H Y, et al. Effect of adsorption of cadmium from aqueous solution by hexadecyl trimethyl ammonium bromide modified biochar[J]. Chinese Journal of Environmental Engineering, 2019, 13(8): 1809-1821. |
| [32] |
刘红娟, 党志, 张慧, 等. 蜡状芽孢杆菌抗重金属性能及对镉的累积[J]. 农业环境科学学报, 2010, 29(1): 25-29. Liu H J, Dang Z, Zhang H, et al. Characterization of heavy metal resistance and Cd-accumulation on a Bacillus cereus strain[J]. Journal of Agro-Environment Science, 2010, 29(1): 25-29. |
| [33] |
谢超然, 王兆炜, 朱俊民, 等. 核桃青皮生物炭对重金属铅、铜的吸附特性研究[J]. 环境科学学报, 2016, 36(4): 1190-1198. Xie C R, Wang Z W, Zhu J M, et al. Adsorption of lead and copper from aqueous solutions on biochar produced from walnut green husk[J]. Acta Scientiae Circumstantiae, 2016, 36(4): 1190-1198. |
| [34] | Hu X, Ding Z H, Zimmerman A R, et al. Batch and column sorption of arsenic onto iron-impregnated biochar synthesized through hydrolysis[J]. Water Research, 2015, 68: 206-216. |
| [35] | Zhu X M, Chen B L, Zhu L Z, et al. Effects and mechanisms of biochar-microbe interactions in soil improvement and pollution remediation:A review[J]. Environmental Pollution, 2017, 227: 98-115. |
| [36] |
陈温福, 张伟明, 孟军. 农用生物炭研究进展与前景[J]. 中国农业科学, 2013, 46(16): 3324-3333. Chen W F, Zhang W M, Meng J. Advances and prospects in research of biochar utilization in agriculture[J]. Scientia Agricultura Sinica, 2013, 46(16): 3324-3333. |
| [37] |
王婷.高效诱变菌与生物炭复合修复重金属污染土壤的研究[D].天津: 南开大学, 2013. Wang T. The bioremediation of heavy metal contaminated soil by bioaugmentation of an active mutant bacterium assisted by biochar[D]. Tianjin: Nankai University, 2013. |
| [38] |
罗海艳, 李丹阳, 刘寿涛, 等. 铁锰改性椰壳炭对土壤镉形态及水稻吸收积累镉的影响[J]. 环境科学研究, 2019, 32(5): 139-147. Luo H Y, Li D Y, Liu S T, et al. Effects of Fe-Mn modified coconut shell biochar on cadmium speciation and accumulation in rice[J]. Research of Environmental Sciences, 2019, 32(5): 139-147. |
| [39] | Jayarathne A, Egodawatta P, Ayoko G A, et al. Role of residence time on the transformation of Zn, Cu, Pb and Cd attached to road dust in different land uses[J]. Ecotoxicology and Environmental Safety, 2018, 153: 195-203. |
| [40] |
杨文弢, 王英杰, 周航, 等. 水稻不同生育期根际及非根际土壤砷形态迁移转化规律[J]. 环境科学, 2015, 36(2): 694-699. Yang W T, Wang Y J, Zhou H, et al. Transformation and mobility of arsenic in the rhizosphere and non-rhizosphere soils at different growth stages of rice[J]. Environmental Science, 2015, 36(2): 694-699. |
| [41] |
刘玉玲, 铁柏清, 李园星露, 等. 耐镉细菌的分离及其对土壤中镉的形态影响[J]. 农业环境科学学报, 2018, 37(2): 250-258. Liu Y L, Tie B Q, Li Y X L, et al. Isolation of a Cd-resistant bacterium and its effect on the speciation of Cd in soil[J]. Journal of Agro-Environment Science, 2018, 37(2): 250-258. |
| [42] |
张燕, 铁柏清, 刘孝利, 等. 玉米秸秆生物炭对稻田土壤砷、镉形态的影响[J]. 环境科学学报, 2018, 38(2): 715-721. Zhang Y, Tie B Q, Liu X L, et al. Effects of waterlogging and application of bio-carbon from corn stalks on the physico-chemical properties and the forms of arsenic and cadmium in arsenic and cadmium-contaminated soils[J]. Acta Scientiae Circumstantiae, 2018, 38(2): 715-721. |
| [43] | Yin D X, Wang X, Peng B, et al. Effect of biochar and Fe-biochar on Cd and As mobility and transfer in soil-rice system[J]. Chemosphere, 2017, 186: 928-937. |
| [44] | Sari A, Tuzen M. Biosorption of As(Ⅲ) and As(Ⅴ) from aqueous solution by macrofungus (Inonotus hispidus) biomass:Equilibrium and kinetic studies[J]. Journal of Hazardous Materials, 2009, 164(2-3): 1372-1378. |
| [45] |
杨婷.微生物细胞表面的化学/基因改性调控用于重金属分离及(形态)分析[D].沈阳: 东北大学, 2013. Yang T. Chemical/genetical modification of microbial cell for heavy metal separation and analysis/speciation[D]. Shenyang: Northeastern University, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10145-1016011625.htm |
| [46] |
刘玉玲, 彭鸥, 铁柏清, 等. Delftia sp. B9对镉胁迫下水稻种子萌发及幼苗镉积累的影响[J]. 农业环境科学学报, 2019, 38(8): 1855-1863. Liu Y L, Peng O, Tie B Q, et al. Effects of Delftia sp. B9 on rice seed germination and cadmium accumulation in rice seedlings under Cd stress[J]. Journal of Agro-Environment Science, 2019, 38(8): 1855-1863. |
| [47] | Nair A, Juwarkar A A, Singh S K. Production and characterization of siderophores and its application in arsenic removal from contaminated soil[J]. Water, Air, and Soil Pollution, 2007, 180(1-4): 199-212. |
| [48] | Crans D C, Smee J J, Gaidamauskas E, et al. The chemistry and biochemistry of vanadium and the biological activities exerted by vanadium compounds[J]. Chemical Reviews, 2004, 104(2): 849-902. |
| [49] | Liu Y L, Tie B Q, Peng O, et al. Inoculation of Cd-contaminated paddy soil with biochar-supported microbial cell composite:A novel approach to reducing cadmium accumulation in rice grains[J]. Chemosphere, 2020, 247: 125850. |