 2020, Vol. 41
2020, Vol. 41

2. 稻米品质安全控制湖南省工程实验室, 长沙 410004;
3. 有色金属矿区耕地重金属污染生态阻抗技术研究衡阳市重点实验室, 衡阳 421001
 , ZHOU Hang1,2
, ZHOU Hang1,2 , GU Jiao-feng1,2 , HUO Yang1 , DENG Peng-hui1 , WEI Bin-yun1 , LIU Jun3 , LIAO Bo-han1,2
, GU Jiao-feng1,2 , HUO Yang1 , DENG Peng-hui1 , WEI Bin-yun1 , LIU Jun3 , LIAO Bo-han1,2
2. Hunan Engineering Laboratory for Control of Rice Quality and Safety, Changsha 410004, China;
3. Key Laboratory of Hengyang City on Ecological Impedance Technology of Heavy Metal Pollution in Cultivated Soil of Nonferrous Metal Mining Area, Hengyang 421001, China
全球土壤中重金属铅(Pb)污染问题严重, 采矿冶炼、固废堆置和污水灌溉等人类活动加剧了土壤重金属污染[1].Pb具有高毒性, 不可被生物降解[2], 土壤中过量Pb会对水稻品质产生不利影响, 并通过食物链对人类健康构成威胁[3].据报道, 我国华南和华中地区15个水稻主产区产出的稻米中, 有12.9%的样品Pb含量超过Pb≤0.2 mg·kg-1的标准限值[4, 5].因此, 土壤Pb污染导致的稻米Pb超标问题亟需解决.
近年来, 水稻对重金属的吸收、转运和累积受到广泛关注.水稻对重金属的吸收很大程度上取决于重金属在土壤中的有效性和植株中的迁移转运过程[6].有研究发现, 水稻不同生育期由于生长状况和营养需求等因素的影响, 对重金属的吸收与转运差异明显.王倩倩等[7]和叶长城等[8]通过水培试验结果表明, 水稻糙米Cd累积的关键生育期为灌浆期和成熟期.彭华等[9]通过在水稻分蘖期和孕穗期施用硅肥发现, 水稻糙米中Cd含量比未施硅肥下降25.3%. Zhou等[10]通过测定不同生育时期水稻各部位Cd含量表明, 叶片中累积的Cd在孕穗期到成熟期进行了再转运, 对糙米中Cd累积的贡献率为22.5%~30.0%.刘昭兵等[11]通过采集不同生育期水稻研究发现, 根系、茎和叶中Cd和Pb含量为:分蘖期>成熟期>抽穗期.还有研究表明, 糙米中Pb的累积是通过水稻籽粒发育过程中根的吸收、根-茎转运及茎-谷粒转运而来[12, 13].当前大量研究都聚焦于水稻不同生育期Cd的吸收转运, 部分研究讨论水稻植株Pb吸收累积的关键生育期, 但鲜有研究关注水稻不同生育期Pb胁迫对成熟期水稻植株和糙米Pb累积贡献.
本研究通过水培试验对水稻不同生育期单独供给Pb, 分析水稻不同生育期对Pb吸收累积的差异, 通过探讨水稻吸收累积Pb的关键生育时期, 明确不同生育期吸收累积的Pb对糙米中Pb累积的贡献, 以期为Pb污染稻田安全生产提供科学依据.
1 材料与方法 1.1 供试材料试供水稻为湘晚籼13号, 常规中熟晚稻品种, 全生育期103 d, 由湖南省水稻研究所提供.水稻培养营养液采用木村B营养液[14].水培容器为可装2.5 L营养液的圆柱形塑料桶(内径20 cm, 高24 cm).
1.2 试验设计水培试验在中南林业科技大学生命科学楼三楼露台进行, 光、气、热均为自然状态, 亚热带季风湿润气候, 雨热同期, 四季分明. 2018年7月4日, 选取颗粒饱满、性状大小无明显差异的水稻种子用30% H2O2消毒15 min, 用去离子水洗净, 再用湿润的纱布包裹, 置于人工培养箱(温度30℃, 湿度50%)中培养发芽, 芽长至1 cm左右, 播种于未受Pb污染的土壤中. 2018年7月16日, 选取培育后株高为15 cm左右、长势良好的水稻幼苗(三叶一心), 根系用自来水冲洗, 去除根系附着的黏土, 用泡沫浮板固定海绵包裹水稻幼苗, 放入长方体塑料盆中进行前期培养, 使水稻幼苗正常生长, 满足后期试验要求. 2018年7月26日选择长势相近的水稻幼苗, 五叶一心, 移入圆柱形塑料桶进行水培试验.
本试验设计7个外源Pb添加处理(见表 1), 分别且仅在分蘖期(S分蘖, 29 d)、拔节期(S拔节, 16 d)、孕穗期(S孕穗, 13 d)、灌浆期(S灌浆, 17 d)、蜡熟期(S蜡熟, 14 d)和成熟期(S成熟, 14 d)添加外源Pb, 以及全生育期(S全, 103 d)添加外源Pb, 同时设置全生育期不添加外源Pb作为空白(CK)对照.外源Pb选用醋酸铅[(CH3COO)2Pb], 使水培中营养液Pb浓度为0.5 mg·L-1.本试验期间, 每3 d更换一次营养液, 使用稀HNO3和NaOH溶液调节营养液的pH, 使pH维持在5.5.试验各处理均重复3次.水稻种植期间, 根据实际情况喷洒农药, 防止病虫害.
|
|
表 1 试验设计 Table 1 Experimental design |
1.3 水稻样品采集
水稻成熟后, 采集整株水稻, 用自来水去除水稻表面尘土, 再用超纯水清洗3次后分剪.将水稻植株分为稻谷、穗轴、叶、茎、茎节1(茎节为茎与叶的连结点, 茎节1为茎连接穗和旗叶的最上面节点)、其他茎节(除了茎节1以外的茎节)、根基茎(根与茎连结2 cm处)和根[15, 16].根系放入5 mmol·L-1的CaCl2溶液浸泡15 min[17], 用于去除根系表面吸附的Pb2+, 再和其他部位一同自然风干24 h后, 放入烘箱105℃杀青30 min, 再70℃烘干至恒重.水稻各部位分别称量, 记录干重.稻谷使用小型砻谷机(JLGJ4.5, 浙江台州市粮仪厂)脱壳后, 分离谷壳和糙米.水稻各部位均用粉碎机(JFSJ-100-1, 浙江台州市粮仪厂)粉碎, 装入密封袋保存待测.
1.4 样品分析水稻各部位样品采用干灰化法处理, 具体为称取3.0 g水稻样品粉末于30 mL坩埚中, 在马弗炉中碳化至无黑烟(250℃), 然后500℃高温灰化6~8 h.取出冷却, 加入0.5 mol·L-1 HNO3溶解灰分, 并多次洗涤坩埚, 定容到50 mL容量瓶中, 保存备测.用石墨炉原子吸收分光光度计(iCE-3500, Thermo-Fisher)测定水稻样品中Pb含量.在分析过程中, 所有水稻样品以湖南大米[GBW10045(GSB-23)]进行质量控制, Pb的回收率为94.1%~104.9%.
1.5 数据统计与分析不同生育期Pb胁迫对水稻植株、地上部位和糙米Pb累积的相对贡献率计算方式见下式:
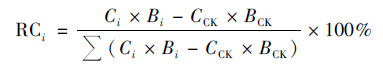
|
式中, RC为相对贡献率(%), Ci为不同生育期Pb胁迫下某部位Pb含量(mg·kg-1), CCK为对照下某部位Pb含量(mg·kg-1), Bi为不同生育期Pb胁迫下某部位生物量(干重, g·株-1), BCK为对照下某部位生物量(干重, g·株-1), i为不同生育期Pb胁迫(分蘖期、拔节期、孕穗期、灌浆期、蜡熟期和成熟期).
本研究数据统计与分析采用Excel 2013和SPSS 22.0单因素ANOVA中Duncan多重比较法分析各处理间差异(P<0.05);水稻各部位Pb含量表示为平均值±标准偏差(n=3), 图形采用Origin 2018进行绘制.
2 结果与分析 2.1 不同生育期Pb胁迫对水稻生长的影响对水稻分蘖数、株高和生物量进行分析测定, 结果如表 2和图 1.可以看出, 0.5 mg·L-1的Pb胁迫对水稻生长有一定的影响.从水稻植株分蘖数来看, 6个单一时期Pb胁迫处理, 分蘖数范围为17~21蘖·株-1, 与CK之间均不存在显著差异, 而全生育期Pb胁迫(S全)处理时水稻分蘖数(23蘖·株-1)显著高于对照CK处理(19蘖·株-1).从株高来看, 拔节期Pb胁迫(S拔节)处理时水稻株高明显增加, 与CK(70.5 cm)相比高出9.9%, 而其他各单一时期Pb胁迫处理均与CK无显著差异.S全处理时水稻株高最低, 仅为59.0 cm, 与CK和各单一时期Pb胁迫处理之间均存在显著差异(P<0.05).从生物量来看, CK处理的总生物量最大, 为61.0 g·株-1, S全处理的总生物量最小, 仅为47.4 g·株-1, 比CK下降了22.3%.与CK相比, 各单一时期Pb胁迫处理使总生物量降低了6.3%~15.7%.显然, 不同生育时期0.5 mg·L-1的Pb胁迫对水稻植株的生物量具有一定的抑制作用.
|
|
表 2 不同生育期Pb胁迫下水稻成熟期分蘖数和株高1) Table 2 Effects of exogenous Pb stress at different growth stages on tiller numbers and heights of rice plants at the maturing stage |

|
图 1 不同生育期Pb胁迫下水稻不同部位生物量 Fig. 1 Effects of exogenous Pb stress at different growth stages on the biomass of different rice tissues |
表 3是不同生育期Pb胁迫对水稻各部位Pb含量的影响.从中可知, 水稻各部位Pb含量在不同生育期Pb胁迫处理下存在明显差异.除S全处理时水稻地上部位和根系中Pb含量最大外, 其余6个单一时期处理中, S分蘖、S拔节和S孕穗处理水稻地上部位Pb含量显著高于S灌浆、S蜡熟和S成熟处理, 而根系Pb含量与地上部位相反, 即S灌浆、S蜡熟和S成熟处理根中Pb含量显著高于S分蘖、S拔节和S孕穗.
|
|
表 3 不同生育期Pb胁迫下水稻成熟期各部位Pb含量1) Table 3 Effects of exogenous Pb stress at different growth stages on Pb content in different rice tissues at the maturing stage |
对于不同部位Pb含量, 7个处理各部位中Pb含量均显著高于CK(P < 0.05), S全处理各部位中Pb含量远高于其他6个单一时期Pb胁迫处理.各处理根系Pb含量变化范围较大, 为30.8~936.2 mg·kg-1; 其次为茎节1、其他茎节和根基茎中Pb含量, 变化范围分别为0.3~162.9、2.4~103.1和5.4~27.0 mg·kg-1, 其中茎节1的Pb含量平均值高于其他茎节Pb含量平均值; 茎、叶和穗轴Pb含量变化范围相对较小, 分别为0.4~15.8、1.4~8.4和0.7~34.1 mg·kg-1.在单一时期Pb胁迫处理中, S孕穗处理糙米Pb含量最高为1.2 mg·kg-1, 显著高于其他处理(P < 0.05);其次为S拔节和S分蘖处理, 糙米Pb含量分别为0.7 mg·kg-1和0.5 mg·kg-1, S灌浆、S蜡熟和S成熟处理糙米中Pb含量较低.
2.3 不同生育期Pb胁迫对水稻各部位Pb累积的影响图 2(a)是不同生育期Pb胁迫对水稻各部位Pb累积的影响, 图 2(b)是不同生育期Pb胁迫对水稻糙米Pb累积的影响.水稻各部位(糙米、谷壳、穗轴、叶、茎、茎节1、其他茎节、根基茎和根)Pb累积量为水稻各部位Pb含量与各部位生物量的乘积.

|
图 2 不同生育期Pb胁迫下水稻成熟期各部位Pb累积量 Fig. 2 Effects of exogenous Pb stress at different growth stages on Pb accumulation amounts in different rice tissues at the maturing stage |
由图 2(a)可知, 不同生育期Pb胁迫处理下水稻植株Pb累积量显著高于对照CK处理, 变化范围为357.2~5 785.6 μg·株-1, 其中根系Pb累积量占比最大.根系Pb累积量变化范围为172.6~5 113.5 μg·株-1, 占植株Pb总累积量的48.3%~98.2%, 平均为75.3%. 7个不同处理根系Pb累积量从高到低依次为:S全>S蜡熟>S成熟>S灌浆>S拔节>S分蘖≈S孕穗.其中, S全处理时根Pb累积量最高, 为5 113.5 μg·株-1, 其次是S蜡熟处理, 其根Pb累积量为1969.1 μg·株-1, 是地上部位Pb总累积量的55倍; S分蘖和S孕穗处理下根系Pb累积量相对较低, 分别为172.6 μg·株-1和187.3 μg·株-1. 7个不同处理地上部位Pb累积量变化范围为36.2~276.9 μg·株-1, Pb累积量排序为:S全>S拔节>S分蘖≈S孕穗>S灌浆>S蜡熟≈S成熟.由图 2(b)可以看出, 糙米Pb累积量变化范围为0.6~6.6 μg·株-1, 在单一时期Pb胁迫处理中, 糙米Pb累积量在S孕穗处理时最大, 为5.3 μg·株-1, 其次是S拔节和S分蘖处理, 分别为3.2 μg·株-1和2.8 μg·株-1.
2.4 不同生育期Pb胁迫对水稻Pb累积的相对贡献率表 4为不同生育期Pb胁迫对水稻植株、地上部位和糙米Pb累积的相对贡献率.不同生育期Pb胁迫下, 水稻植株的相对贡献率大小顺序为:S蜡熟>S成熟>S灌浆>S拔节>S孕穗>S分蘖, 相对贡献率分别为37.7%、27.4%、14.9%、11.3%、4.6%和4.1%.不同生育期Pb胁迫下水稻地上部位的相对贡献率大小顺序为:S拔节>S孕穗>S分蘖>S灌浆>S成熟>S蜡熟, 相对贡献率分别为34.5%、23.6%、22.1%、13.9%、3.9%和2.0%.这说明水稻地上部位中Pb主要来源于分蘖期、拔节期和孕穗期的吸收和转运, 水稻根系中Pb主要在灌浆期、蜡熟期和成熟期累积, 并且向地上部位转运Pb的能力减弱.各单一时期Pb胁迫处理糙米Pb累积的贡献率大小为:S孕穗>S拔节>S分蘖>S灌浆>S成熟>S蜡熟.S孕穗处理Pb累积对糙米Pb累积相对贡献率最大, 为43.3%, 其次是S拔节和S分蘖处理, 相对贡献率分别是24.3%和21.3%.
|
|
表 4 不同生育期Pb胁迫下水稻成熟期植株、地上部位和糙米Pb累积的相对贡献率 Table 4 Effects of exogenous Pb stress at different growth stages on relative contributions of Pb accumulation in whole rice plants, aboveground parts, and brown rice at the maturing stage |
3 讨论 3.1 水稻不同生育期Pb累积的差异
本研究中水稻不同生育期吸收的Pb对成熟后各部位Pb累积差异明显.在分蘖期(S分蘖)、拔节期(S拔节)和孕穗期(S孕穗)Pb胁迫下, 水稻糙米、茎、叶和穗轴Pb含量和累积量均较高, 而灌浆期(S灌浆)、蜡熟期(S蜡熟)和成熟期(S成熟)处理糙米、茎、叶和穗轴Pb含量和累积量均较低(表 3和图 2).其原因在于水稻在分蘖期和拔节期是水稻的营养生长期, 生长迅速且新陈代谢作用旺盛, 需要从营养液中吸收大量营养元素, 大量的Pb随着营养物质进入到水稻植株内[18].孕穗期是水稻拔节进一步完成的时期, 水稻长出穗轴, 在该时期吸收的营养物质对水稻后期形成谷壳和糙米贡献较大.水稻不同生育期吸收的Pb对成熟后根系中Pb累积差异也非常明显, 根系Pb含量和累积量大小趋势均为:生殖生长阶段(灌浆期、蜡熟期和成熟期)>营养生长阶段(分蘖期、拔节期和孕穗期).出现这种现象的原因可能在于:①相比于水稻营养生长阶段, 水稻生殖生长阶段根系生物量大, 吸收和附着Pb的位置充足[19], 但水稻叶片光合作用能力和根系生理活性下降[20], 根系向地上部位转运Pb的能力减弱, 大量的Pb被根系阻隔[7, 21]; ②水稻营养生长阶段, 根系根表铁膜处于逐步形成期, 新生根系前端还没有完全覆盖根表铁膜, 阻控Pb向地上部迁移能力较弱; 而生殖生长阶段, 水稻对重金属的解毒能力和耐受性增强[22], 根表铁膜已经形成且逐渐老化变厚, 对Pb具有较强的截留能力[23], 因此后期Pb胁迫的Pb大部分被根表铁膜阻隔并累积在根系中.在全生育期Pb胁迫下, 水稻在前期吸收的Pb主要向地上部位迁移, 而后期主要将Pb滞留在根部, 整个生育期(103 d)水稻受Pb胁迫时间长, 导致水稻植株Pb累积量高达5 785.6 μg·株-1.
本研究中水稻孕穗期(S孕穗)吸收累积的Pb对糙米中Pb的相对贡献率最高, 为43.3%, 其次为拔节期和分蘖期, 相对贡献率分别为24.3%和21.3%(表 4).这与Liu等[24]的研究结果相近, 水稻幼苗时期吸收累积的Pb在后期生长过程中转运至糙米.缪小建等[25]的研究发现, 孕穗期前水稻茎鞘贮存的非结构性碳水化合物和孕穗期后的光合产物, 对启动灌浆初期籽粒库的活性和促进籽粒灌浆起重要作用, 并在孕穗期后转运至穗部籽粒中[26], 同时将累积在茎鞘中的Pb转运至糙米.Xie等[27]的盆栽试验表明, 糙米中Pb大部分来源于根系吸收土壤中的有效Pb通过茎鞘转运到穗轴再传输到糙米, 处于分蘖期时的水稻受主动运输和被动运输双重影响[13].在生育后期水稻根系Pb不易向地上部位转运, 茎和叶中贮藏的Pb仍然会向糙米转运.地上部位Pb累积的相对贡献率在分蘖期、拔节期和孕穗期较高, 分别为22.1%、34.5%和23.6%.在营养生长阶段吸收的Pb较多地累积在地上部位, 而不在根系中累积, 这也可能是Pb向糙米转移的重要来源[28].这些结果说明, 糙米中Pb累积主要来源于孕穗期根系吸收及孕穗期其他部位中Pb向糙米的转运, 因此孕穗期是糙米Pb累积的关键生育期.这为在水稻孕穗期采用淹水灌溉、施加改良剂或叶面阻控等技术方法降低糙米Pb累积提供理论依据.此外, 孕穗末期糙米中Pb含量还可作为评估水稻成熟后糙米Pb含量的一个重要预测指标[24].
3.2 水稻各部位Pb累积转运的过程本研究表明, 不同生育期Pb胁迫水稻根系Pb累积量占植株累积量的48.3%~98.2%, 尤其是灌浆期、蜡熟期和成熟期Pb胁迫处理下根Pb累积量分别为762.9、1 969.1和1 448.6 μg·株-1 [图 2(a)].Pb可通过Ca2+的阳离子转运体进入水稻根系[29], 根部成为外源Pb从外界环境进入水稻植株内最关键的部位.有研究表明, 水稻植株所吸收的Pb大部分被局限于根系组织[30, 31], 根系累积量占水稻植株的累积量甚至超过95%[32].这与本研究中S成熟和S蜡熟处理的研究结果一致, 其根系中Pb累积量均超过95%.穗轴、茎和叶的Pb累积量均在拔节期Pb胁迫下最高, 分别为7.5、115.1和76.3 μg·株-1[图 2(a)].水稻茎鞘是连接穗轴与叶片传输物质的主要通道, 通过蒸腾作用将根系吸收的营养物质和其他物质运输到穗轴和叶片[27, 33].在水稻地上部位中, 茎和叶的生物量平均值远大于其他部位, 导致茎和叶中Pb累积量相对较高.在分蘖期和拔节期水稻茎和叶迅速生长, 茎节也不断增多, 而茎和叶作为水稻营养生长发育最关键的两个部位, 在生物量快速增长的同时, 吸收累积了大量的Pb[8], 这成为水稻进入生殖生长阶段以后糙米中Pb积累的重要来源[28].这与Zhou等[10]的研究结果相类似, 在Cd低污染土壤和高污染土壤种植的水稻, 叶中的Cd含量在乳熟期开始逐渐降低, 并通过茎逐渐向糙米中转运, 对糙米Cd积累的贡献率分别达到30.0%和22.5%.
在分蘖期、拔节期和孕穗期下的茎节中Pb含量皆高于其他地上部位, 其中茎节1的Pb含量平均值高于其他茎节.茎节内扩大维管束的木质部区域比茎和叶的维管束大10倍以上, 这种差异会降低木质部中的质量流速度[34], 从而使Pb在茎节中保留时间更长, 因此茎节中Pb含量较高.Yamaji等[15]的研究认为, 茎节是矿物元素优先分布的枢纽, 茎节1中Cd含量高于其他茎节, 与本研究关于Pb的结果相似.因此, Pb在水稻植株中的转运过程可能与Cd的转运相似.水稻植株中Cd向籽粒转运过程是根系中的Cd通过木质部向茎运输, 然后经过木质部向韧皮部转移, 由韧皮部向籽粒运输, 最终在籽粒中累积[35].本研究通过相关性分析发现(表 5), 单一生育期Pb胁迫处理下水稻根系Pb含量与茎节1(r=-0.933)和其他茎节(r=-0.965)中Pb含量均呈极显著负相关关系(n=6, r0.01=0.917), 而茎中Pb含量与茎节1(r=0.923)和根基茎(r=0.965)中Pb含量均呈极显著正相关关系(n=6, r0.01=0.917), 与其他茎节中Pb含量呈显著正相关关系(r=0.914; n=6, r0.05=0.811), 叶中Pb含量与茎节1中Pb含量呈显著正相关关系(r=0.898; n=6, r0.05=0.811), 与根基茎中Pb含量呈极显著正相关关系(r=0.954; n=6, r0.01=0.917), 这说明Pb在水稻各部位之间的迁移转运与茎节紧密相关.有研究发现, 茎节正是物质从木质部向韧皮部转移的中心器官, 在水稻的分蘖期、拔节期和孕穗期长出.茎节是阻隔重金属进入糙米的一道屏障, Pb由根部吸收后转运到地上部分, 首先通过茎节被重新分布, 一部分进入茎鞘、谷壳和糙米, 另一部分进入叶片[36].
|
|
表 5 不同生育期水稻各部位Pb含量相关性1) Table 5 Correlations of Pb contents in different rice tissues at different growth stages |
4 结论
(1) 各单一时期Pb胁迫处理较CK水稻株高和分蘖数不存在显著影响, 对生物量有一定的抑制作用, 而全生育期Pb胁迫处理下水稻株高和生物量均受到明显抑制.
(2) 水稻成熟后不同部位的Pb含量差异表现为:根系>茎节1>其他茎节>根基茎>茎>叶>穗轴>谷壳>糙米.不同生育期Pb胁迫处理糙米Pb含量变化范围为0.1~1.2 mg·kg-1, 其顺序为:孕穗期Pb胁迫>拔节期Pb胁迫>分蘖期Pb胁迫>灌浆期Pb胁迫>成熟期Pb胁迫>蜡熟期Pb胁迫.
(3) 水稻植株地上部位和糙米中Pb累积量相对贡献率均为:水稻营养生长阶段(分蘖期、拔节期和孕穗期)>生殖生长阶段(灌浆期、蜡熟期和成熟期), 其中孕穗期为糙米Pb积累的关键生育期, 对糙米Pb累积的相对贡献率为43.3%, 其次是拔节期和分蘖期处理, 相对贡献率分别为24.4%和21.3%.
(4) 糙米中Pb累积主要来源于孕穗期根系吸收及孕穗期其他部位中Pb向糙米的转运, 建议在水稻孕穗期适度采用淹水灌溉、施加改良剂或叶面阻控等技术措施, 以降低糙米Pb的累积, 实现Pb污染稻田安全利用.
| [1] | Liu J, Wang D Q, Song B, et al. Source apportionment of Pb in a rice-soil system using field monitoring and isotope composition analysis[J]. Journal of Geochemical Exploration, 2019, 204: 83-89. DOI:10.1016/j.gexplo.2019.05.009 |
| [2] | Yang Y J, Chen J M, Huang Q N, et al. Can liming reduce cadmium (Cd) accumulation in rice (Oryza sativa) in slightly acidic soils? A contradictory dynamic equilibrium between Cd uptake capacity of roots and Cd immobilisation in soils[J]. Chemosphere, 2018, 193: 547-556. DOI:10.1016/j.chemosphere.2017.11.061 |
| [3] | Verbruggen N, Hermans C, Schat H. Mechanisms to cope with arsenic or cadmium excess in plants[J]. Current Opinion in Plant Biology, 2009, 12(3): 364-372. DOI:10.1016/j.pbi.2009.05.001 |
| [4] | Fang Y, Sun X Y, Yang W J, et al. Concentrations and health risks of lead, cadmium, arsenic, and mercury in rice and edible mushrooms in China[J]. Food Chemistry, 2014, 147: 147-151. DOI:10.1016/j.foodchem.2013.09.116 |
| [5] | Yang Q W, Shu W S, Qiu J W, et al. Lead in paddy soils and rice plants and its potential health risk around Lechang Lead/Zinc Mine, Guangdong, China[J]. Environment International, 2004, 30(7): 883-889. DOI:10.1016/j.envint.2004.02.002 |
| [6] | Wan Y N, Huang Q Q, Camara A Y, et al. Water management impacts on the solubility of Cd, Pb, As, and Cr and their uptake by rice in two contaminated paddy soils[J]. Chemosphere, 2019, 228: 360-369. DOI:10.1016/j.chemosphere.2019.04.133 |
| [7] |
王倩倩, 贾润语, 李虹呈, 等. Cd胁迫水培试验下水稻糙米Cd累积的关键生育时期[J]. 中国农业科学, 2018, 51(23): 4424-4433. Wang Q Q, Jia R Y, Li H C, et al. Key growth stage of Cd accumulation in brown rice through a hydroponic experiment with Cd stress[J]. Scientia Agricultura Sinica, 2018, 51(23): 4424-4433. DOI:10.3864/j.issn.0578-1752.2018.23.003 |
| [8] |
叶长城, 陈喆, 彭鸥, 等. 不同生育期Cd胁迫对水稻生长及镉累积的影响[J]. 环境科学学报, 2017, 37(8): 3201-3206. Ye C C, Chen Z, Peng O, et al. Effects of cadmium stress on growth and cadmium accumulation in rice at different growth stages[J]. Acta Scientiae Circumstantiae, 2017, 37(8): 3201-3206. |
| [9] |
彭华, 田发祥, 魏维, 等. 不同生育期施用硅肥对水稻吸收积累镉硅的影响[J]. 农业环境科学学报, 2017, 36(6): 1027-1033. Peng H, Tian F X, Wei W, et al. Effects of silicon fertilizer application on the cadmium and silicon content of rice at different growth stages[J]. Journal of Agro-Environment Science, 2017, 36(6): 1027-1033. |
| [10] | Zhou H, Zhu W, Yang W T, et al. Cadmium uptake, accumulation, and remobilization in iron plaque and rice tissues at different growth stages[J]. Ecotoxicology and Environmental Safety, 2018, 152: 91-97. DOI:10.1016/j.ecoenv.2018.01.031 |
| [11] |
刘昭兵, 纪雄辉, 彭华, 等. 不同生育期水稻对Cd、Pb的吸收累积特征及品种差异[J]. 土壤通报, 2011, 42(5): 1125-1130. Liu Z B, Ji X H, Peng H, et al. Characteristics of Cd and Pb absorption and accumulation by rice at different growth stages and the differences between varieties[J]. Chinese Journal of Soil Science, 2011, 42(5): 1125-1130. |
| [12] | Cheng W D, Zhang G P, Yao H G, et al. Effect of grain position within a panicle and variety on As, Cd, Cr, Ni, Pb concentrations in japonica rice[J]. Rice Science, 2005, 12(1): 48-56. |
| [13] | Zhou H, Zeng M, Zhou X, et al. Heavy metal translocation and accumulation in iron plaques and plant tissues for 32 hybrid rice (Oryza sativa L.) cultivars[J]. Plant and Soil, 2015, 386(1-2): 317-329. DOI:10.1007/s11104-014-2268-5 |
| [14] | 薛应龙. 植物生理学实验手册[M]. 上海: 上海科学技术出版社, 1985. |
| [15] | Yamaji N, Xia J X, Mitani-Ueno N, et al. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2[J]. Plant Physiology, 2013, 162(2): 927-939. DOI:10.1104/pp.113.216564 |
| [16] |
莫争, 王春霞, 陈琴, 等. 重金属Cu, Pb, Zn, Cr, Cd在水稻植株中的富集和分布[J]. 环境化学, 2002, 21(2): 110-116. Mo Z, Wang C X, Chen Q, et al. Distribution and enrichment of heavy metals of Cu, Pb, Zn, Cr and Cd in paddy plant[J]. Environmental Chemistry, 2002, 21(2): 110-116. DOI:10.3321/j.issn:0254-6108.2002.02.002 |
| [17] |
胡莹, 黄益宗, 段桂兰, 等. 镉对不同生态型水稻的毒性及其在水稻体内迁移转运[J]. 生态毒理学报, 2012, 7(6): 664-670. Hu Y, Huang Y Z, Duan G L, et al. Cadmium toxicity and its translocation in two ecotype rice cultivar[J]. Asian Journal of Ecotoxicology, 2012, 7(6): 664-670. |
| [18] |
许嘉琳, 鲍子平, 杨居荣, 等. 农作物体内铅、镉、铜的化学形态研究[J]. 应用生态学报, 1991, 2(3): 244-248. Xu J L, Bao Z P, Yang J R, et al. Chemical forms of Pb, Cd and Cu in crops[J]. Chinese Journal of Applied Ecology, 1991, 2(3): 244-248. DOI:10.3321/j.issn:1001-9332.1991.03.010 |
| [19] |
史锟, 张福锁, 刘学军, 等. 不同时期施铁对水稻根表铁胶膜中铁镉含量及根系含镉量的影响[J]. 农业环境科学学报, 2004, 23(1): 6-12. Shi K, Zhang F S, Liu X J, et al. Effects of different periods applied Fe2+ and concentrations on Fe and Cd contents in iron plaque on rice root[J]. Journal of Agro-Environment Science, 2004, 23(1): 6-12. DOI:10.3321/j.issn:1672-2043.2004.01.002 |
| [20] |
李香玲, 冯跃华. 水稻根系生长特性及其与地上部分关系的研究进展[J]. 中国农学通报, 2015, 31(6): 1-6. Li X L, Feng Y H. Research advance on relation of aerial part and root traits of rice[J]. Chinese Agricultural Science Bulletin, 2015, 31(6): 1-6. |
| [21] | Guo J X, Li Y Y, Hu C, et al. Ca-containing amendments to reduce the absorption and translocation of Pb in rice plants[J]. Science of the Total Environment, 2018, 637-638: 971-979. DOI:10.1016/j.scitotenv.2018.05.100 |
| [22] |
彭鸥, 铁柏清, 叶长城, 等. 稻米镉关键积累时期研究[J]. 农业资源与环境学报, 2017, 34(3): 272-279. Peng O, Tie B Q, Ye C C, et al. The key period of cadmium accumulation in rice[J]. Journal of Agricultural Resources and Environment, 2017, 34(3): 272-279. |
| [23] |
胡莹, 黄益宗, 黄艳超, 等. 不同生育期水稻根表铁膜的形成及其对水稻吸收和转运Cd的影响[J]. 农业环境科学学报, 2013, 32(3): 432-437. Hu Y, Huang Y Z, Huang C Y, et al. Formation of iron plaque on root surface and its effect on Cd uptake and translocation by rice (Oryza sativa L.) at different growth stages[J]. Journal of Agro-Environment Science, 2013, 32(3): 432-437. |
| [24] | Liu J G, Ma X M, Wang M X, et al. Genotypic differences among rice cultivars in lead accumulation and translocation and the relation with grain Pb levels[J]. Ecotoxicology and Environmental Safety, 2013, 90: 35-40. DOI:10.1016/j.ecoenv.2012.12.007 |
| [25] |
缪小建, 王绍华, 李刚华, 等. 大中穗型水稻品种灌浆期非结构性碳水化合物运转特征[J]. 南京农业大学学报, 2009, 32(2): 1-4. Miao X J, Wang S H, Li G H, et al. Translocation of non-structural carbohydrates in large and medium panicle rice cultivars during grain filling stage[J]. Journal of Nanjing Agricultural University, 2009, 32(2): 1-4. DOI:10.3969/j.issn.1671-7465.2009.02.001 |
| [26] |
刘慧娟, 饶玉春, 杨窑龙, 等. 水稻叶鞘相关性状的遗传分析[J]. 分子植物育种, 2011, 9(3): 278-287. Liu H J, Rao Y C, Yang Y L, et al. QTL analysis of leaf sheath traits in rice(Oryza sativa L.)[J]. Molecular Plant Breeding, 2011, 9(3): 278-287. DOI:10.3969/mpb.009.000278 |
| [27] | Xie T H, Li Y Y, Dong H X, et al. Effects and mechanisms on the reduction of lead accumulation in rice grains through lime amendment[J]. Ecotoxicology and Environmental Safety, 2019, 173: 266-272. DOI:10.1016/j.ecoenv.2019.02.010 |
| [28] | Ashraf U, Hussain S, Akbar N, et al. Water management regimes alter Pb uptake and translocation in fragrant rice[J]. Ecotoxicology and Environmental Safety, 2018, 149: 128-134. DOI:10.1016/j.ecoenv.2017.11.033 |
| [29] | Kim Y Y, Yang Y Y, Lee Y. Pb and Cd uptake in rice roots[J]. Physiologia Plantarum, 2002, 116(3): 368-372. |
| [30] |
段德超, 于明革, 施积炎. 植物对铅的吸收、转运、累积和解毒机制研究进展[J]. 应用生态学报, 2014, 25(1): 287-296. Duan D C, Yu M G, Shi J Y. Research advances in uptake, translocation, accumulation and detoxification of Pb in plants[J]. Chinese Journal of Applied Ecology, 2014, 25(1): 287-296. |
| [31] |
林舜华, 黄银晓, 姚依群, 等. 铅在植物-土壤系统中的分配规律及其生态效应——以大豆、水稻为例[J]. 植物生态学与地植物学丛刊, 1985, 9(2): 85-91. Lin S H, Huang Y X, Yao Y Q, et al. The distribution of Pb in the plant-soil system and its ecological reaction[J]. Acta Phytoecologica et Geobotanica Sinica, 1985, 9(2): 85-91. |
| [32] |
朱珺. NRT1.1在植物响应铅胁迫中的作用及机制[D].杭州: 浙江大学, 2019. Zhu J. The role of nitrate transporter 1.1 in plant response to Pb stress and its underling mechanism[D]. Hangzhou: Zhejiang University, 2019. http://cdmd.cnki.com.cn/Article/CDMD-10335-1019038891.htm |
| [33] |
周鸿, 曲仲湘, 王焕校. 铅对几种农作物的影响及迁移积累规律初探[J]. 环境科学学报, 1983, 3(3): 222-234. Zhou H, Qu Z X, Wang H X. A study on the effects of lead on several kinds of crops and its translocation and accumulation therein[J]. Acta Scientiae Circumstantiae, 1983, 3(3): 222-234. |
| [34] | Yamaji N, Ma J F. The node, a hub for mineral nutrient distribution in graminaceous plants[J]. Trends in Plant Science, 2014, 19(9): 556-563. DOI:10.1016/j.tplants.2014.05.007 |
| [35] | Fujimaki S, Suzui N, Ishioka N S, et al. Tracing cadmium from culture to spikelet:noninvasive imaging and quantitative characterization of absorption, transport, and accumulation of cadmium in an intact rice plant[J]. Plant Physiology, 2010, 152(4): 1796-1806. DOI:10.1104/pp.109.151035 |
| [36] | Feng X M, Han L, Chao D Y, et al. Ionomic and transcriptomic analysis provides new insight into the distribution and transport of cadmium and arsenic in rice[J]. Journal of Hazardous Materials, 2017, 331: 246-256. DOI:10.1016/j.jhazmat.2017.02.041 |