 2020, Vol. 41
2020, Vol. 41



2. 中国科学院大学, 北京 100049;
3. 江南大学环境与土木工程学院, 无锡 214122
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. School of Environment and Civil Engineering, Jiangnan University, Wuxi 214122, China
在浅水湖泊中, 湖水与底泥间物质交换强烈, 对污染响应比较敏感, 因此浅水湖泊极易发生富营养化.而沉水植物作为浅水湖泊中的重要生产者, 在湖泊生态系统的物质循环和能量流动方面发挥着重要作用, 不仅可与浮游藻类竞争水中的光、能源和营养物质[1], 对藻类生长起到抑制作用[2, 3]; 还可以有效抑制沉积物再悬浮过程[1, 2], 并为浮游动物、底栖动物以及鱼类等提供适宜的栖息地.因此, 沉水植物丰富的区域往往具有更高的水体透明度[3]和生物多样性[4].丰富的沉水植物也被认为是健康水生生态系统的重要表征[5], 恢复水生植被成为浅水湖泊富营养化生态修复的重要措施[6].
然而, 沉水植物对湖泊生态系统的影响并不一定都是积极的.沉水植物的生长具有一定的周期性, 如果没有合适的群落演替或是不能在其死亡前做到及时有效的收割, 其腐烂分解会向水体中大量释放氮、磷等生源要素[7~11]、改变水体光环境[12]并可能为藻类水华暴发或湖泛的发生提供物质和环境条件[13], 进一步增加湖泊的富营养化程度.尤其是对于死亡分解过程发生在夏季的水生植物, 高温环境可能会促使蓝藻门的部分种属(如微囊藻)占据优势和大量繁殖[12], 进而威胁饮用水安全和水生生态系统的服务功能.
菹草(Potamogeton crispus)隶属于眼子菜科, 就是一种夏季死亡的沉水植物.其在秋冬发芽, 4月和5月达到最大现存量, 到6月开始大量死亡并腐烂分解[14].菹草在我国的浅水富营养化湖泊中广泛分布[15].此外, 该物种对弱光具有较强的耐受性[16], 常被用作富营养化湖泊生态恢复的先锋种.已有研究表明, 生长期的菹草能有效吸附水体中的总磷等营养元素[17]及金属元素[18], 明显改善水质; 相对地, 菹草衰亡时, 茎叶中吸附的营养物质会重新进入水体, 造成水体溶解氧下降、氮磷浓度升高的现象, 引发二次污染.然而, 已有的研究多集中于室内控制实验, 控制体系较为单一, 对湖泊自然环境下菹草的环境效益研究较少[19], 尤其是菹草对浮游植物群落结构的影响明显缺乏, 而后者对于浅水湖泊, 尤其是富营养化的湖泊研究是极为重要的.
因此, 本文以菹草为例, 选取了我国江淮平原湖群14个水文背景相似的浅水湖泊, 在5月菹草生长季节和7月死亡分解阶段分别对湖泊水体的理化指标和浮游植物群落结构进行调查, 通过探讨菹草的生长与衰亡对水环境和浮游植物生物量与群落结构的影响, 尤其是对蓝藻种属的影响, 以期为菹草这一常见沉水植物的生态管控提供科学建议.
1 材料与方法 1.1 研究区域概况本文以我国江淮中下游平原地区的14个浅水湖泊为研究对象(图 1).江淮中下游平原湖区包括我国长江及淮河中下游、黄河下游等地区的自然湖泊, 是我国五大湖区之一.据统计, 该区域面积大于1 km2的自然湖泊面积约占全国湖泊总面积为23.3%, 是我国湖泊分布密度最大的地区之一[20].该区气候以亚热带季风与季风性湿润气候为主, 夏季高温多雨.湖泊多由河流水系演变或者海岸线变迁形成, 具有相似的水文地质背景, 湖泊较浅, 换水周期较长, 降水和河川径流是其主要的补给水源.同时该区是我国人口最密集的地区之一, 湖泊具有重要的经济和生态价值, 是周围城市和地区的工业用水、调蓄灌溉、航运水产和生活取水的重要来源; 但近年来受人类活动影响强烈, 该区大部分湖泊都处于不同程度的富营养化程度.

|
图 1 采样湖泊空间分布示意 Fig. 1 Spatial distribution of the study lakes |
在菹草的生长阶段(春季, 2018年4月27日~5月11日)和死亡阶段(夏季, 2018年7月5~20日), 调查了我国江淮平原生态区的14个浅水湖泊(图 1)的64个点位.包括在春季有菹草生长的26个有草点和没有水生植被覆盖的38个无草点.其中东平湖和梁子湖所有采样点在春季均被菹草所覆盖, 武昌湖、黄大湖、龙感湖、武山湖、珠湖、军山湖、武昌东湖和岳阳南湖这8个湖的采样点未出现任何水生植被生长现象; 而南四湖、洪湖、阳澄湖和白马湖, 既包括有菹草生长的生境也包括无菹草生境.所有的湖中采样点均匀分布, 在有菹草点, 菹草是春季水生植物群落的绝对优势种, 目测估计菹草覆盖度达95%以上; 除了阳澄湖在夏季出现了演替种荷花和菱角, 其余湖泊在夏季均未出现其它水生植物.
利用便携式多参数水质分析仪(YSI, 6600V2)原位测定了水面表层的水温(WT)、溶解氧(DO)和pH.使用Secchi盘在现场测量各点位的透明度(SD).在每个点位收集表层50 cm水样300 mL, 并立即冷藏带回实验室用以分析总氮(TN)、总磷(TP)和高锰酸盐指数.其中, TN采用过硫酸钾紫外分光光度法测定, TP采用钼酸铵分光光度法测定; 高锰酸盐指数采用酸性高锰酸盐滴定法测定.叶绿素a(Chl-a)采用热乙醇萃取分光光度法测量, 在每个点采集定量水通过0.45 μm孔径的玻璃纤维滤膜(Whatman GF/F), 并用铝箔纸收集滤膜, 包好冷藏送回实验室, 用于提取Chl-a; 同时收集滤膜后的水, 用于测定溶解性总氮(DTN)、溶解性总磷(DTP)和氨氮(NH4+-N).此外, 在每个点位另外采集1 L水, 现场加入1%的酸性鲁哥溶液(即10 mL)用于浮游植物的固定, 然后带回实验室后沉淀48 h并浓缩后, 再在显微镜下参照文献[21]对样品浮游植物的种类进行镜检鉴定和计数.各种属藻类生物量是由该种属藻细胞数量和其对应的细胞重量乘积计算所得.
1.3 蓝藻对营养盐利用性氮磷元素是限制湖泊中浮游植物生长的重要因子之一[22], 水体的TN、TP浓度与浮游植物的生物量或叶绿素a浓度常呈现显著的正相关关系[23].但环境因子, 如温度、光照等气象因子和地理位置的变化会强烈影响营养盐与藻类生产力的关系[24]. Chl-a/TP的比值常常被用来衡量和评估指定水体环境中营养盐被蓝藻的利用效率[23, 25].因此, 本文分别计算了有草区和无草区的蓝藻生物量与TP和TN的比值(Cyano/TP和Cyano/TN), 用以比较两种环境下蓝藻生长的适宜性大小.即利用不同环境下蓝藻生物量与所利用的营养盐浓度比值来评估该环境下蓝藻生长的潜力, 对于越适宜蓝藻生长的环境, 这一比值越大.
1.4 统计分析利用Kolmogorov-Smirnov和Levene检验对所获取的数据进行正态和方差齐性检验, 对通过检验的数据采用单因素方差分析(one-way ANOVA)的方法对有无菹草生长区之间的SD、DO、TN、TP和高锰酸盐指数进行比较; 采用配对样本检验(Paired-samples T test)的方法对春夏季节间同区位的水质参数差异进行分析.所有样本的相关性通过Pearson相关分析进行检验.对于不符合正态分布和方差齐性的数据, 使用Wilcoxon秩和检验[26]的方法进行非参数比较.选用浮游植物和蓝藻生物量的质量数据进行分析.
利用相似性分析(analysis of similarities, ANOSIM)对浮游植物和蓝藻门各属的细胞密度数据进行分析, 以量化无草区和有草区两者间浮游植物群落结构和蓝藻种属的差异.并利用非度量多维标度(nonmetric multidimensional scaling, NMDS)对差异性并进行比较; NMDS的特点是根据样品中包含的物种信息, 以点的形式反映在多维空间上; 通过点与点间的距离(Bray-curtis相异性矩阵距离)体现不同样品间的差异程度, 以减少优势类群的影响[27].同时利用公式(1)对所获浮游植物样本的优势种进行计算.
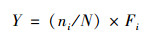
|
(1) |
式中, ni为第i种的个体数, N为样品中的个体总数, Fi为样品中第i种个体在总样本中出现的频率, 当优势度Y>0.02时, 判定该种为优势种[28].
2 结果与分析 2.1 水质物理化学参数有菹草生长区(有草区)和无草区的水质理化参数如表 1所示.结果表明, 菹草的生长提升了水体的SD、DO和pH.具体表现为春季有草区SD达(114±43)cm, 显著高于无草区(74±53)cm(P=0.001).有草区的DO为(12.24±2.82)mg·L-1也高于无草区(9.88±2.26)mg·L-1(P < 0.001); pH也表现为有草区显著较高(P < 0.001).而在夏季, 有菹草衰亡区的SD为(66±35)cm, 在均值上低于无草区的(86±75)cm(P=0.34), DO、pH在有草区和无草区间也不存在统计上的差异.
|
|
表 1 有无菹草对水环境参数值的影响1) Table 1 Influence of the presence or absence of P. crispus on water environment parameters |
配对样本检验的结果表明, 从春季菹草生长期到夏季死亡期的过程中, 有草区的SD显著下降(P < 0.001, n1=n2=26), 而无草区的SD在季节间不存在显著差异(P=0.34, n1=n2=38).对于DO, 有草区的DO从春季的(12.24±2.82)mg·L-1降至夏季的(9.27±3.36)mg·L-1, 表明菹草在春季的生长, 光合作用大大提升了水体的溶解氧水平(P < 0.001, n1=n2=26);有草区春夏季的高锰酸盐指数分别为(5.26±2.05)mg·L-1和(6.20±1.64)mg·L-1, 表现出显著的上升(P=0.028, n1=n2=26), 说明在菹草分解阶段, 水中的有机质含量增加.而无草区的DO、pH和高锰酸盐指数在季节间均未表现出较大差异.
考虑到不同湖泊本身的水文背景的不同, 以及受人类活动的影响, 不同点位营养负荷的差异, 各样本的营养数据间的差异较大.但对于TP, 不管是春季还是夏季, 均表现为有草区的TP较低.而TN在区位间和季节间均没有表现出显著的差异; 但在夏季有草区的NH4+-N显著高于无草区(P < 0.001).
2.2 浮游植物 2.2.1 Chl-a和蓝藻生物量在春季, 无草区和有草区的Chl-a浓度分别为(25.83±39.39)μg·L-1和(11.86±16.09)μg·L-1(表 1), 有草区的Chl-a浓度略低于无草区(P=0.06).浮游植物和蓝藻生物量的结果表明(图 2), 春季的浮游植物种类鉴定出7门31属, 其中无草区浮游植物的总生物量为0.60 mg·L-1, 优势种包括鱼腥藻、束丝藻、颤藻、直链藻和栅藻, 其中蓝藻生物量0.10 mg·L-1, 占总生物量的17.02%.有草区的浮游植物的总生物量为0.43 mg·L-1, 与无草区间没有显著差异(P=0.49);主要优势种为席藻和直链藻, 其中蓝藻生物量分别为0.06 mg·L-1, 占总生物量的比例为13.93%, 显著低于无草区(P=0.05).

|
图 2 有无菹草区浮游植物总生物量和蓝藻生物量(Mean±SE) Fig. 2 Histogram of phytoplankton and cyanobacteria biomass under P. crispus presence or absence (mean biomass±SE) |
夏季, 两点位的Chl-a浓度间没有明显差异(P=0.27), 分别为(45.66±25.96)μg·L-1和(46.99±39.55)μg·L-1.但浮游植物的总生物量在统计上表现出显著差异(P=0.014), 有草区的浮游植物总生物量为2.68 mg·L-1, 高于无草区的生物量2.29 mg·L-1; 相对应的有草区蓝藻生物量高达0.91 mg·L-1, 与无草区蓝藻生物量0.44 mg·L-1间差异极显著(P < 0.001), 两者各占总生物量的34.05%和18.96%, 有草区的浮游植物优势种为鱼腥藻、平裂藻、束丝藻和颤藻, 均属于蓝藻门, 无草区的优势种包括束丝藻、颤藻、席藻、鱼腥藻、微囊藻和直链藻多种.可以看出, 尽管浮游藻类的总生物量和蓝藻的生物量在季节间存在较大的增加, 但对于无草区, 蓝藻占总生物量的比例并没有增长的趋势, 而有草区, 夏季蓝藻所占的比例有较大幅度的增长.相似性分析的结果显示, 造成夏季藻类种群明显不同的是蓝藻中的颤藻, 颤藻在夏季有草区的生物量显著增加.
2.2.2 蓝藻对营养盐的利用效率利用蓝藻生物量和营养盐(TN和TP)的比值来量化指定环境下蓝藻对营养盐的利用效率(表 2).在春季, 有草区和无草区的Cyano/TP分别为(1.05±2.71)和(0.97±2.21)(P=0.07), Cyano/TN分别为(0.05±0.15)和(0.06±0.12)(P=0.38), 均没有表现出显著差异.而夏季两者的Cyano/TP比值分别为(16.20±15.55)和(7.46±9.59)(P < 0.001), 表明有草区蓝藻对TP的利用效率极显著高于无草区蓝藻对TP的利用效率; 类似的结果也出现在蓝藻对TN的利用上, 夏季Cyano/TN在无草区为(0.44±0.50), 而在有草区高达(0.78±0.60), 显著高于无草区蓝藻对TN的利用率(P=0.002).表明在菹草死亡的环境下更适宜蓝藻的大量增殖, 此时的蓝藻能够对TN具更高的吸收转化率.
|
|
表 2 有无菹草区蓝藻生物量和营养盐(TN和TP)的比值 Table 2 Ratio of Cyanophyta biomass and nutrients (total phosphorus and total nitrogen) under P. crispus presence or absence |
3 讨论 3.1 菹草的生长与衰亡过程对水环境的影响
已有研究发现, 菹草在生长阶段有改善水体光环境、降低水相中营养盐水平的作用[29].本研究通过较大空间尺度的现场调研, 结果表明, 菹草对水体透明度的改善主要因为两方面的原因.第一, 菹草特殊的波浪式叶片, 能加快水体中悬浮颗粒物和泥沙的絮凝作用, 可快速降低外源非藻类浊度.室内实验表明, 在24 h的水力停留时间下, 菹草可使输入水体的悬浮泥沙质量浓度降低约70%[30].第二, 沉水植物的大量生长可以很好地抑制沉积物再悬浮的过程, 从而降低内源无机颗粒物对透明度的影响[4].此外, 植物的光合作用提升了水体的溶解氧, 植株的茎叶能为附生微生物的生存提供环境, 增加细菌的多样性[31], 促进水中有机质的降解.
而在夏季, 菹草的死亡分解造成了水体SD、pH值和DO的显著下降, 有机质含量显著升高.其主要原因是菹草死亡后, 沉积物的再悬浮过程加剧; 并且失去菹草对光的竞争作用, 浮游植物获得了更好地光照条件并得以快速生长[24], 使水体浊度上升.此外, 菹草植株的分解也会在一定程度上对水体的光照条件造成影响, 使水体浊度快速下降, 并破坏底栖生物的生境.因为菹草植株由大量的有机物组成, 其中干植株的有机物含量可占84%[32]; 在菹草死亡时, 植株快速腐烂, 研究表明半数生物量的分解完成的时间约25 d[14], 80%左右的初始干重在40 d后消失[18]; 且开始分解速率较高, 后逐渐降低.根叶的凋零和分解生成有机物溶入水体中, 会释放出大量的有机物.高锰酸盐指数作为水中还原性有机物的指示量, 与水中碳源含量密切正相关[33], 本研究中也发现, 有草区的高锰酸盐指数值在夏季明显升高, 显著高于无草区的这一数值.表明菹草植株的分解, 在一定程度上使水体中的有机质含量增加, 从而能为浮游植物的生长提供了更多的能量.
此外, 植株有机碎屑的分解过程中, 会大量消耗水中的DO, 使水体底部处于短时的缺氧状态[34], 影响水下底栖生物和微生物的群落结构, 甚至诱发湖泛的出现[13], 在本研究中也观察到水体DO的降低, 但未出现明显的缺氧环境, 可能是因为浅水湖泊受到风浪的扰动较大.水体的扰动不仅会影响营养盐和有机物的释放的过程与速率[32], 还会增加水体的复氧情况.微生物在分解菹草的过程中, 同时也会代谢产生CO2释放回水体中, 降低水体pH值[34].
营养盐方面, 菹草的生长显著降低了水相的磷水平.不论是春季还是夏季, TP的浓度均表现为有草区低于无草区.茂盛生长期的菹草可以从水体和沉积物中大量吸收磷等营养物质[17], 并将其储存到植株体或者转移至沉积物中[35, 36], 从而显著降低水相的TP水平.其次, 对于浅水湖泊来说, 沉积物再悬浮过程一直是内源磷释放的重要途径之一, 沉水植被生长改变底泥氧化还原条件, 抑制底泥磷的释放, 进一步降低了水相TP, 从而限制藻类的生长.而在夏季时, 沉水植物的分解使大量磷得以重新释放[9, 10], 促进磷酸盐酶的活性, 常常使上覆水中TP浓度出现较大的增加趋势[7, 37].
理论上来说, 菹草的生长也会显著降低水相氮的水平.比如, 有研究表明太湖表层沉积物中TN和总有机碳含量都与水生植物的现存量间存在显著的正相关关系[38], 说明湖水中的氮和有机碳很可能是经过水生植物的沉积作用, 被传输到了底泥之中.此外, 除了植物吸收, 沉积物的吸附和微生物的反硝化作用都是水体氮素去除的重要途径[7], 直接影响水相中的TN浓度.因此, 在水生植物生物量较低时, 沉水植物分解过程中也会出现上覆水的TN和TP比初始浓度低的情况[7].本研究中菹草对TN和TP的浓度影响在统计上就并不显著, 但伴随夏季蓝藻的产生, NH4+-N浓度明显升高, 与前人的研究结果一致[39].基于野外实地观测, 湖泊面积广阔和蓄水量大, 不同于室内实验所设定的水体体积, 营养盐有足够的空间进行转化和吸附是重要的原因.另一方面, 野外环境中, 营养盐浓度与水文等多因素密切相关.比如水位, 水位对营养盐调节具有复杂性, 既可以通过增加径流带来流域上的营养物质的输入, 又可以对湖水有一定的稀释作用, 改变氮磷比[40], 在一定程度上能够缓冲菹草分解时释放的氮磷营养盐浓度.
3.2 菹草的死亡分解对藻类生物量和群落结构的影响对于一个稳定生态系统来说, 藻类的群落结构会随着温度、水文因子在季节间的改变而发生规律性的演替, 比如, 在亚热带的水库中, 浮游植物在春季常以硅藻、甲藻和绿藻为优势类群, 夏季转为以蓝藻、绿藻和硅藻为主; 秋季随着温度降低, 蓝藻和绿藻减少, 硅藻和甲藻再次占优势[41].水文和营养盐等环境条件的快速变化同样会使浮游植物的群落结构和优势种发生改变, 使个别门类的细胞数量占比大幅增加[42], 如许多富营养化的浅水湖泊中, 蓝藻成为了夏季的常见优势种, 大量繁殖, 极易引发水华, 产生神经毒素、细胞毒素和肝毒素等多种毒素和异味, 引起水生生物的死亡和食物网结构的改变, 严重损害人体的健康和生态系统的服务功能[43].因此, 蓝藻的大量生长需要引起足够的重视.
本研究中, 春季有草区的叶绿素含量和浮游植物总生物量都明显地低于无草区.这是由于春季沉水植物在与浮游植物竞争中占优势, 占据了水体中的光和营养物质等条件, 在一定程度上抑制了藻类的生物量[2]; 研究已证实, 沉水植物及其附着的生物膜会产生化感物质, 抑制藻类和细菌的生物量[44, 45].此外, 沉水植物还会为底栖生物提供栖息的环境, 对藻类产生一定的牧食作用.此时蓝藻的生物量均较低, 春季不是蓝藻的最佳生长环境.在夏季, 虽然有草和无草区的Chl-a含量和浮游植物总生物量上未表现出明显差别, 但由春夏两季有草区和无草区间蓝藻和总生物量的占比可知, 尽管浮游藻类的总生物量和蓝藻的生物量在季节间存在较大的增加, 但是在无草区, 蓝藻占总生物量的比例并没有增长的趋势(分别为17.02%和18.96%); 而有草区蓝藻占总生物量的比例有较大幅度的增长(分别为13.63%和34.05%).在有草区, 夏季蓝藻所占的比例不仅高于春季的比例, 同时也高于无草区夏季的蓝藻所占比例, 大幅增加.
这是因为在菹草衰亡的生境中, 蓝藻对营养盐的利用效率大大提升, 即相同的水相磷浓度, 蓝藻获得了更大的生长量.首先, 夏季蓝藻与其它藻类的营养竞争中常常处于优势地位.蓝藻的最佳生长温度高于其它藻类(通常高于25℃), 在夏季表现出更好的生长速率和水华潜力[46].其次, 蓝藻对强光和弱光的耐受性都较好, 如颤藻对光的耐受性较强, 微囊藻会通过增加细胞内类胡萝卜素的含量来免受强光的抑制; 在较低的光照条件下蓝藻可用较少的能量维持细胞的结构和功能, 比硅藻或绿藻[12]等具有更高的生长速率[47]和生物量.在菹草死亡时, 水体浊度上升, 其他种群的生长受到抑制, 是利于蓝藻生长的正反馈环境.此外某些蓝藻凭借伪空泡调节自身浮力, 进而迁移到光照充足和营养丰富的表层水体区域, 获得生长[42].在本研究中, 夏季时候, 水温在有草区和无草区分别达到(29.9±1.5)℃和(31.0±1.4)℃, 较春季上升.适合蓝藻生长, 加上菹草死亡为藻类的生长提供了较大生态位空缺, 包括营养盐生态位和光的生态位等, 蓝藻更有利于成为优势属.此外, 植株残体的分解, 还会为水体提供大量的有机物[48], 为藻类生长提供碳源.还有在植株死亡阶段, 对藻类的抑制效应或释放的化感物质会突然消失, 这种抑制效应突然消失对藻类生物群落的影响还不甚清楚和难以量化.因此, 有草区的蓝藻在营养盐的利用效率上远高于无草区.
3.3 菹草在自然湖泊中的恢复与管理菹草现已经成为江淮平原地区浅水湖泊的主要优势类群之一, 有研究发现, 该区湖泊水生植物多样性降低.例如, 通过植被指数对东平湖的菹草识别发现, 三十多年间菹草群落不断扩张, 由湖岸边零星分布逐渐蔓延至全湖且连片式分布, 且菹草覆盖的面积与湖水营养化的水平显著正相关[15].夏季菹草整株死亡, 分解和释放大量营养盐, 同时水底沉积物在悬浮作用变强, 水体急剧变浑浊, 同时腐烂的植株为水体提供了充足的有机质, 为蓝藻的生长提供适宜的环境, 使蓝藻对营养盐有更强的吸收和转化效率, 获得较大的生物量[44], 并进一步引发蓝藻水华的暴发.
单一沉水植物的生长对水体环境并不是完全积极的.当菹草在湖泊中生物量过大时, 还会较大地抑制了其它水生植物的生存空间, 阻碍其它物种的定居与扩展[49], 造成了湖泊水体高等植物的单一化, 不利于水生植物种群季节性的正常演替[44]和湖泊生态系统的长期稳定.此外, 菹草死亡时还会大量消耗水中的溶解氧, 甚至造成水底缺氧, 影响底栖生物的生存, 破环水生环境的稳定性.
因此, 沉水植物的控制和管理非常重要, 特别是对于富营养化浅水湖泊的恢复[7]; 要充分考虑沉水植物带来的二次污染[18].在生态恢复的过程中, 应该对大型植物的选择与引种进行筛选[3], 尽量选择沉水植物、漂浮植物等多种物种的镶嵌组合形式, 注重多种植物的演替行为, 增强对群落中菹草或其它水生植物的生物量控制[49], 构建更具异质性和多样性的水体环境.
4 结论菹草是浅水湖泊中的常见的沉水植物类群, 在江淮平原的一些浅水湖泊中已逐渐成为优势种, 并常作为沉水植物先锋种用于水体的生态修复.菹草在生长过程中可以促进水中悬浮颗粒物的沉降, 改善水体光环境, 吸附水中营养物质并将其输送到植株体内或底泥中, 达到净化水质的效果.但是, 在菹草大量腐烂的时候, 植株的分解和底泥再悬浮过程, 会使植物体和沉积物中的有机质和营养素大量释放, 快速改变水环境, 引起藻类增殖, 尤其是富营养化湖泊中常见的蓝藻.浑浊的水体环境和夏季的高温为蓝藻的生长提供了竞争优势, 加上充足的有机质和营养素, 极大加重了富营养化湖泊中蓝藻水华暴发的风险, 对水体生态环境产生极其不利的影响.因此, 亟需要加强对浅水湖泊中沉水植物的控制和管理, 在水生植物修复时, 注重水生植物的演替行为, 维持其在蓝藻易暴发的夏秋季的生长量, 构建更健康安全的水生生态系统群落结构.
| [1] | Gao J Q, Xiong Z T, Zhang J D, et al. Phosphorus removal from water of eutrophic Lake Donghu by five submerged macrophytes[J]. Desalination, 2009, 242(1-3): 193-204. |
| [2] |
鲜啟鸣, 陈海东, 邹惠仙, 等. 四种沉水植物的克藻效应[J]. 湖泊科学, 2005, 17(1): 75-80. Xian Q M, Chen H D, Zou H X, et al. Allelopathic effects of four submerged macrophytes on Microcystis aeruginosa[J]. Journal of Lake Sciences, 2005, 17(1): 75-80. DOI:10.3321/j.issn:1003-5427.2005.01.012 |
| [3] | Wang G X, Zhang L M, Chua H, et al. A mosaic community of macrophytes for the ecological remediation of eutrophic shallow lakes[J]. Ecological Engineering, 2009, 35(4): 582-590. |
| [4] | Lv T, He Q K, Hong Y P, et al. Effects of water quality adjusted by submerged macrophytes on the richness of the epiphytic Algal Community[J]. Frontiers in Plant Science, 2019, 9: 1980. |
| [5] | Søndergaard M, Johansson L S, Lauridsen T L, et al. Submerged macrophytes as indicators of the ecological quality of lakes[J]. Freshwater Biology, 2010, 55(4): 893-908. DOI:10.1111/j.1365-2427.2009.02331.x |
| [6] | Gulati R D, Van Donk E. Lakes in the Netherlands, their origin, eutrophication and restoration: state-of-the-art review[A]. In: Nienhuis P H, Gulati R D, (Eds.). Ecological Restoration of Aquatic and Semi-Aquatic Ecosystems in the Netherlands (NW Europe)[M]. Dordrecht: Springer, 2002. 73-106. |
| [7] | Li C H, Wang B, Ye C, et al. The release of nitrogen and phosphorus during the decomposition process of submerged macrophyte (Hydrilla verticillata Royle) with different biomass levels[J]. Ecological Engineering, 2014, 70: 268-274. DOI:10.1016/j.ecoleng.2014.04.011 |
| [8] | Wang L Z, Liu Q J, Hu C W, et al. Phosphorus release during decomposition of the submerged macrophyte Potamogeton crispus[J]. Limnology, 2018, 19(3): 355-366. |
| [9] | Zhou X H, He Z L, Ding F H, et al. Biomass decaying and elemental release of aquatic macrophyte detritus in waterways of the Indian River Lagoon basin, South Florida, USA[J]. Science of the Total Environment, 2018, 635: 878-891. DOI:10.1016/j.scitotenv.2018.04.047 |
| [10] | Carignan R, Kalff J. Phosphorus release by submerged macrophytes:significance to epiphyton and phytoplankton[J]. Limnology and Oceanography, 1982, 27(3): 419-427. DOI:10.4319/lo.1982.27.3.0419 |
| [11] |
魏宏农, 潘建林, 赵凯, 等. 菹草附着物对营养盐浓度的响应及其与菹草衰亡的关系[J]. 生态学报, 2013, 33(24): 7661-7666. Wei H N, Pan J L, Zhao K, et al. Response of periphyton to nutrient level and relationships between periphyton and decay degree of Potamogeton crispus[J]. Acta Ecologica Sinica, 2013, 33(24): 7661-7666. |
| [12] | Paerl H W, Huisman J. Blooms like it hot[J]. Science, 2008, 320(5872): 57-58. DOI:10.1126/science.1155398 |
| [13] | Shen Q S, Zhou Q L, Shang J G, et al. Beyond hypoxia:occurrence and characteristics of black blooms due to the decomposition of the submerged plant Potamogeton crispus in a shallow lake[J]. Journal of Environmental Sciences, 2014, 26(2): 281-288. |
| [14] |
陈志刚, 林志军, 周晓红, 等. 菹草腐烂分解过程中污染物的动态释放[J]. 干旱区资源与环境, 2017, 31(8): 153-159. Chen Z G, Lin Z J, Zhou X H, et al. Pollutants dynamic release of Potamogeton crispus during decomposition process[J]. Journal of Arid Land Resources and Environment, 2017, 31(8): 153-159. |
| [15] |
梁莉莉, 于泉洲, 邓焕广, 等. 基于时序NDVI的东平湖菹草(Potamogeton crispus L.)遥感提取及时空格局[J]. 湖泊科学, 2019, 31(2): 529-538. Liang L L, Yu Q Z, Deng H G, et al. Spatio-temporal pattern of Potamogeton crispus L. in Lake Dongping based on NDVI time series[J]. Journal of Lake Sciences, 2019, 31(2): 529-538. |
| [16] | Song X X, Wang Z, Xiao B D, et al. Growth of Potamogeton crispus L. from turions in darkness:Implications for restoring submerged plants in eutrophic lakes[J]. Ecological Engineering, 2017, 101: 255-260. DOI:10.1016/j.ecoleng.2017.01.035 |
| [17] |
任文君, 胡晓波, 刘霞, 等. 白洋淀菹草对富营养化水体总磷的净化[J]. 应用生态学报, 2011, 22(4): 1053-1058. Ren W J, Hu X B, Liu X, et al. Total phosphorus removal from eutrophic water in Baiyangdian Lake by Potamogeton crispus[J]. Chinese Journal of Applied Ecology, 2011, 22(4): 1053-1058. |
| [18] | Deng H G, Zhang J, Chen S Y, et al. Metal release/accumulation during the decomposition of Potamogeton crispus in a shallow macrophytic lake[J]. Journal of Environmental Sciences, 2016, 42: 71-78. DOI:10.1016/j.jes.2015.07.004 |
| [19] | Cao X Y, Wan L L, Xiao J, et al. Environmental effects by introducing Potamogeton crispus to recover a eutrophic lake[J]. Science of the Total Environment, 2018, 621: 360-367. DOI:10.1016/j.scitotenv.2017.11.267 |
| [20] | 王苏民, 窦鸿身. 中国湖泊志[M]. 北京: 科学出版社, 1998. |
| [21] |
胡鸿钧, 魏印心. 中国淡水藻类[M]. 北京: 科学出版社, 2006. Hu H J, Wei Y X. The freshwater algae of China[M]. Beijing: Science Press, 2006. |
| [22] | Rangel L M, Silva L H S, Rosa P, et al. Phytoplankton biomass is mainly controlled by hydrology and phosphorus concentrations in tropical hydroelectric reservoirs[J]. Hydrobiologia, 2012, 693(1): 13-28. DOI:10.1007/s10750-012-1083-3 |
| [23] | Mazumder A. Phosphorus-chlorophyll relationships under contrasting herbivory and thermal stratification:predictions and patterns[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1994, 51(2): 390-400. DOI:10.1139/f94-040 |
| [24] | Huo S L, He Z S, Ma C Z, et al. Spatio-temporal impacts of meteorological and geographic factors on the availability of nitrogen and phosphorus to algae in Chinese lakes[J]. Journal of Hydrology, 2019, 572: 380-387. DOI:10.1016/j.jhydrol.2019.03.010 |
| [25] | Havens K E, Nürnberg G K. The phosphorus-chlorophyll relationship in lakes:potential influences of color and mixing regime[J]. Lake and Reservoir Management, 2004, 20(3): 188-196. DOI:10.1080/07438140409354243 |
| [26] | Wilcoxon F. Individual comparisons by ranking methods[A]. In: Kotz S, Johnson N L, (Eds). Breakthroughs in Statistics[M]. New York, NY: Springer, 1992. 196-202. |
| [27] | Vilas M P, Marti C L, Oldham C E, et al. Macrophyte-induced thermal stratification in a shallow urban lake promotes conditions suitable for nitrogen-fixing cyanobacteria[J]. Hydrobiologia, 2018, 806(1): 411-426. DOI:10.1007/s10750-017-3376-z |
| [28] |
刘金殿, 顾志敏, 杨元杰, 等. 长诏水库浮游植物群落结构及水质评价[J]. 生态学杂志, 2012, 31(11): 2865-2871. Liu J D, Gu Z M, Yang Y J, et al. Phytoplankton community structure and water quality assessment of Changzhao Reservoir, Zhejiang of East China[J]. Chinese Journal of Ecology, 2012, 31(11): 2865-2871. |
| [29] | Qiu D R, Wu Z B, Liu B Y, et al. The restoration of aquatic macrophytes for improving water quality in a hypertrophic shallow lake in Hubei Province, China[J]. Ecological Engineering, 2001, 18(2): 147-156. |
| [30] |
曹昀, 王国祥. 冬季菹草对悬浮泥沙的影响[J]. 生态与农村环境学报, 2007, 23(1): 54-56. Cao Y, Wang G X. Effects of submersed plant Potamogeton crispus on suspended mud and sand[J]. Journal of Ecology and Rural Environment, 2007, 23(1): 54-56. DOI:10.3969/j.issn.1673-4831.2007.01.012 |
| [31] |
李琳琳, 汤祥明, 高光, 等. 沉水植物生态修复对西湖细菌多样性及群落结构的影响[J]. 湖泊科学, 2013, 25(2): 188-198. Li L L, Tang X M, Gao G, et al. Influence of submerged vegetation restoration on bacterial diversity and community composition in West Lake[J]. Journal of Lake Sciences, 2013, 25(2): 188-198. DOI:10.3969/j.issn.1003-5427.2013.02.003 |
| [32] |
卢少勇, 陈建军, 覃进, 等. 扰动强度对菹草浸泡过程中氮磷碳释放的影响[J]. 环境科学, 2011, 32(7): 1940-1944. Lu S Y, Chen J J, Qin J, et al. Influence of disturbance intensity on nitrogen, phosphorus and permanganate index release of Potamogeton crispus during soaking in water[J]. Environmental Science, 2011, 32(7): 1940-1944. |
| [33] | 马永才, 李英, 韩永生, 等. 总有机碳与高锰酸盐指数及化学需氧量的相关性[J]. 环境监测管理与技术, 2001, 3(13): 40-41. |
| [34] |
朱庆川, 杨卫东, 程红. 菹草在水体中腐烂分解试验研究[J]. 现代农业科技, 2013(23): 224-226, 229. Zhu Q C, Yang W D, Cheng H. Test of Potamogeton crispus decomposing in water[J]. Modern Agricultural Science and Technology, 2013(23): 224-226, 229. DOI:10.3969/j.issn.1007-5739.2013.23.144 |
| [35] | Zhou X H, Wang M Y, Wen C Z, et al. Nitrogen release and its influence on anammox bacteria during the decay of Potamogeton crispus with different values of initial debris biomass[J]. Science of the Total Environment, 2019, 650: 604-615. DOI:10.1016/j.scitotenv.2018.08.358 |
| [36] |
潘慧云, 徐小花, 高士祥. 沉水植物衰亡过程中营养盐的释放过程及规律[J]. 环境科学研究, 2008, 21(1): 64-68. Pan H Y, Xu X H, Gao S X. Study on process of nutrition release during the decay of submerged macrophytes[J]. Research of Environmental Sciences, 2008, 21(1): 64-68. |
| [37] | Zhou Y W, Zhou X H, Han R M, et al. Reproduction capacity of Potamogeton crispus fragments and its role in water purification and algae inhibition in eutrophic lakes[J]. Science of the Total Environment, 2017, 580: 1421-1428. DOI:10.1016/j.scitotenv.2016.12.108 |
| [38] |
雷泽湘, 徐德兰, 顾继光, 等. 太湖大型水生植物分布特征及其对湖泊营养盐的影响[J]. 农业环境科学学报, 2008, 27(2): 698-704. Lei Z X, Xu D L, Gu J G, et al. Distribution characteristics of aquatic macrophytes and their effects on the nutrients of water and sedi-ment in Taihu Lake[J]. Journal of Agro-Environment Science, 2008, 27(2): 698-704. DOI:10.3321/j.issn:1672-2043.2008.02.054 |
| [39] | Jacoby J M, Collier D C, Welch E B, et al. Environmental factors associated with a toxic bloom of Microcystis aeruginosa[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2000, 57(1): 231-240. DOI:10.1139/f99-234 |
| [40] | Yang J, Lv H, Liu L M, et al. Decline in water level boosts cyanobacteria dominance in subtropical reservoirs[J]. Science of the Total Environment, 2016, 557-558: 445-452. DOI:10.1016/j.scitotenv.2016.03.094 |
| [41] |
胡韧, 雷腊梅, 韩博平. 南亚热带大型贫营养水库浮游植物群落结构与季节变化——以新丰江水库[J]. 生态学报, 2008, 28(10): 4652-4664. Hu R, Lei L M, Han B P. Phytoplankton assemblage and seasonal dynamics in the large oligotrophic Xinfengjiang reservoir in southern China[J]. Acta Ecologica Sinica, 2008, 28(10): 4652-4664. DOI:10.3321/j.issn:1000-0933.2008.10.005 |
| [42] |
卫志宏, 张利仙, 杨四坤, 等. 洱海浮游植物群落结构及季节演替[J]. 水生态学杂志, 2012, 33(4): 21-25. Wei Z H, Zhang L X, Yang S K, et al. Community structure and seasonal succession of phytoplankton in Erhai Lake[J]. Journal of Hydroecology, 2012, 33(4): 21-25. |
| [43] |
董静, 高云霓, 李根保. 淡水湖泊浮游藻类对富营养化和气候变暖的响应[J]. 水生生物学报, 2016, 40(3): 615-623. Dong J, Gao Y N, Li G B. A review:responses of phytoplankton communities to eutrophication and climate warming in freshwater lakes[J]. Acta Hydrobiologica Sinica, 2016, 40(3): 615-623. |
| [44] |
杨文斌, 王国祥. 南京玄武湖菹草种群的环境效应[J]. 湖泊科学, 2007, 19(5): 572-576. Yang W B, Wang G X. Environmental effects of Potamogeton crispus population in Lake Xuanwu, Nanjing[J]. Journal of Lake Sciences, 2007, 19(5): 572-576. DOI:10.3321/j.issn:1003-5427.2007.05.012 |
| [45] | Nakai S, Inoue Y, Hosomi M, et al. Myriophyllum spicatum-released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa[J]. Water Research, 2000, 34(11): 3026-3032. DOI:10.1016/S0043-1354(00)00039-7 |
| [46] | Paerl H W, Otten T G. Harmful cyanobacterial blooms:causes, consequences, and controls[J]. Microbial Ecology, 2013, 65(4): 995-1010. DOI:10.1007/s00248-012-0159-y |
| [47] |
孔繁翔, 高光. 大型浅水富营养化湖泊中蓝藻水华形成机理的思考[J]. 生态学报, 2005, 25(3): 589-595. Kong F X, Gao G. Hypothesis on cyanobacteria bloom-forming mechanism in large shallow eutrophic lakes[J]. Acta Ecologica Sinica, 2005, 25(3): 589-595. DOI:10.3321/j.issn:1000-0933.2005.03.028 |
| [48] |
金送笛, 李永函, 倪彩虹, 等. 菹草(Potamogeton crispus)对水中氮、磷的吸收及若干影响因素[J]. 生态学报, 1994, 14(2): 168-173. Jin S D, Li Y H, Ni C H, et al. Uptake by Potamogeton crispus of nitrogen and phosphorus from water and some affecting factors[J]. Acta Ecologica Sinica, 1994, 14(2): 168-173. DOI:10.3321/j.issn:1000-0933.1994.02.019 |
| [49] |
徐伟伟, 胡维平, 邓建才, 等. 菹草生物量控制对群落中沉水植物生长及水质的影响[J]. 生态环境学报, 2015, 24(7): 1222-1227. Xu W W, Hu W P, Deng J C, et al. Influence of harvesting Potamogeton crispus in a submerged plant community on the growth of submerged aquatic plants and their effects on water quality[J]. Ecology and Environmental Sciences, 2015, 24(7): 1222-1227. |