 2020, Vol. 41
2020, Vol. 41



2. 中国海洋大学海洋环境与生态教育部重点实验室, 青岛 266100;
3. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室, 青岛 266071;
4. 海南热带海洋学院生态环境学院, 三亚 572022;
5. 中国海洋大学海洋生命学院, 青岛 266003
2. Key Laboratory of Marine Environment and Ecology, Ministry of Education, Ocean University of China, Qingdao 266100, China;
3. Laboratory for Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China;
4. College of Ecology and Environment, Hainan Tropical Ocean University, Sanya 572022, China;
5. College of Marine Life Science, Ocean University of China, Qingdao 266003, China
红树林是分布于热带及亚热带潮间带的特殊木本植物群落, 能够抵御海浪和台风的袭击, 促进土壤形成, 保护海滩生态环境, 被誉为“海岸卫士”[1].红树林生态系统生物资源丰富, 具有高生产力、高归还率和高分解率的特点[2], 对海湾和河口等生态系统的平衡和稳定起到重要作用.硝化作用是氮循环过程中的重要环节, 将氨氮(NH4+-N)氧化为硝态氮(NO3--N), 其中氨氧化过程作为硝化过程的第一步, 是硝化作用的限速步骤[3], 在海洋、沉积物、湿地、农田和湖泊等环境中广泛存在, 在氮素的生物地球化学循环中发挥着重要作用.
氨单加氧酶是好氧氨氧化微生物专有的胞内酶, 由AmoA、AmoB和AmoC这3个亚基组成[4], 其中编码AmoA亚基的基因具有高度的保守性, 同时也能够编码该酶的活性位点, 故目前常用amoA基因作为分子标记基因来研究氨氧化细菌(ammonia-oxidizing bacteria, AOB)和氨氧化古菌(ammonia-oxidizing archaea, AOA)的群落特征.AOB和AOA共同推动硝化作用的进行, 不同环境条件下二者对硝化作用的贡献也不同, 因此AOB和AOA对硝化作用的相对贡献也是氮素循环研究的重要问题.潜在硝化速率(potential nitrification rate, PNR)是指示好氧氨氧化微生物硝化活性的重要指标, 常通过培养的方法[5]得到好氧氨氧化微生物的最大硝化能力, 可通过添加抑制剂如氨苄青霉素[6]区分AOB和AOA对硝化作用的贡献.
已有一系列以红树林沉积物中好氧氨氧化微生物为对象的研究报道.研究者曾在佛罗里达海岸[7]、中国香港米埔湿地自然保护区[8]、海南东寨港红树林湿地[9]和九龙江河口[10]等区域探究了AOB和AOA的空间分布特征; 红树林沉积物中好氧氨氧化微生物的丰度也呈现出明显的季节性差异[11, 12].在九龙江河口区域[10]红树林沉积物中探究了潜在硝化速率与AOB和AOA amoA基因拷贝数之间的相关性关系及影响PNR的关键环境因子.此外, 还有AOB和AOA的群落结构及对环境因子的响应[13]等方面的内容.
三亚河红树林呈窄条带状沿三亚河分布, 是我国最南端、最具典型热带特征[14, 15]且唯一位于城市中心区水道沿岸[16]的红树林, 相较海岸红树林沉积物盐度较低, 受人类活动影响较大.目前缺少三亚河红树林沉积物好氧氨氧化微生物的报道, 故本研究首次对这部分内容进行了补充, 分析了三亚河红树林表层沉积物中好氧氨氧化微生物的分布特征, 并通过模拟培养实验得到AOB和AOA的潜在硝化速率及其对硝化作用的相对贡献率, 对其数量分布和硝化活性进行比较分析, 通过对氨氧化过程的研究提供基础数据, 以期为氮素生物地球化学循环过程中微生物调控机制的研究提供科学依据, 并为红树林生态系统中氮污染控制和生态修复及管理提供参考.
1 材料与方法 1.1 样品采集分别在三亚河上游、中游和下游沿河选择采样区域SY1(18°16′20″N, 109°30′27″E)、SY2(18°15′31″N, 109°30′13″E)和SY3(18°14′50″N, 109°30′55″E), 并在各采样区域分别垂直岸线选取向河侧、红树林中心和向陆侧设置采样点, 各采样点位置分布如图 1所示(因三亚河中游红树林区域十分狭窄, 故只设置了向河侧和向陆侧2个采样点). 2018年冬季(12月)和2019年夏季(6月)采集红树林表层沉积物样品(1~2 cm), 在各采样点现场测定沉积物上覆水中环境因子后, 采用梅花布点法用园艺铲采集样品并放入新的自封袋中, 每个采样点采集3个平行样, 4 h内带回实验室, 一部分样品置于4℃保存, 用于环境因子分析和测定PNR的模拟培养实验; 一部分样品置于-20℃保存, 用于DNA提取及后续实验.使用洁净的塑料水舀和蒸馏水桶现场采集水样, 采集3个平行样.

|
图 1 三亚河红树林表层沉积物采样点分布示意 Fig. 1 Distribution of sampling sites in mangrove surface sediments in the Sanya River |
部分环境因子如温度、溶解氧(dissolved oxygen, DO)、氧化还原电位(Eh)、pH和盐度由便携式测定仪HQ40d(Hach, USA)现场测定; 其余环境因子如总氮(total nitrogen, TN)、总磷(total phosphorus, TP)、总有机碳(total organic carbon, TOC)、氨氮(NH4+-N)、亚硝态氮(NO2--N)和硝态氮(NO3--N)在实验室测定.其中TOC参考环境标准HJ 615-2011采用重铬酸钾氧化-分光光度法进行测定, 其余指标由全自动间断化学分析仪SmartChem 140(AMS Alliance, Italy)测定.表 1列出了各环境因子的测定结果.
|
|
表 1 各采样点的环境因子测定结果 Table 1 Environmental factors at the sampling sites |
1.3 潜在硝化速率测定
潜在硝化速率模拟培养实验参照Bernhard等[17]的方法.使用现场采集的水样通过0.22 μm滤膜过滤, 灭菌, 添加浓度分别为300 μmol ·L-1和60 μmol ·L-1的NH4Cl和KH2PO4.称取2.0 g沉积物样品, 加入60 mL处理后的水样, 以各采样点原位测定的温度为准选择合适温度(冬季样品27℃, 夏季样品32℃), 黑暗条件下连续振荡培养(120 r ·min-1)3 d, 每个样品设置3个平行样.实验中设置培养组1和培养组2, 其中培养组1加入1.0 g ·L-1氨苄青霉素(经验证可完全抑制AOB活性)[18], 计算AOA的PNR; 培养组2不加入氨苄青霉素, 计算AOB和AOA的总PNR; 二者相减即得到AOB的PNR.培养过程中每天分别采集水样并过滤, 置于-20℃保存, 用于测定NH4+-N、NO2--N和NO3--N.通过测得的NO2--N和NO3--N的增加量并考虑溶解无机氮(dissolved inorganic nitrogen, DIN)的损失, 计算PNR[19] [μmol ·(g ·d)-1, 以N计]:
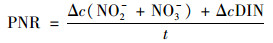
|
(1) |
式中, Δc(NO2-+NO3-)表示培养过程中NO2--N和NO3--N的变化量(μmol ·g-1, 以N计); ΔcDIN表示培养过程中DIN变化量的绝对值(μmol ·g-1, 以N计), 若培养体系中没有DIN损失则ΔcDIN为零; t表示培养时间(d).AOB对硝化作用的相对贡献率可由公式(2)计算得到, AOA对硝化作用的相对贡献率可由公式(3)计算得到[20]:
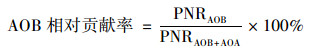
|
(2) |
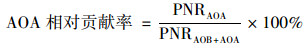
|
(3) |
式中, PNRAOB表示AOB的潜在硝化速率, PNRAOA表示AOA的潜在硝化速率, PNRAOB+AOA表示总潜在硝化速率即AOB和AOA的潜在硝化速率之和.
1.4 DNA提取与PCR扩增称取0.5 g沉积物样品, 使用DNeasy PowerSoilⓇ Kit(QIAGEN, Germany)提取微生物DNA, 提取的DNA样品用超微量紫外分光光度计DS-11(DeNovix, USA)检测浓度和纯度, 置于-20℃保存.分别采用引物amoA-1F/amoA-2R[21]和Arch-amoA26F/Arch-amoA417R[22](表 2)对AOB和AOA的amoA基因进行PCR扩增, 扩增产物通过1%琼脂糖凝胶电泳检测, 确定所得条带与amoA基因条带相符.
|
|
表 2 amoA基因qPCR扩增引物及扩增程序 Table 2 Primers and procedures for qPCR amplification of amoA genes |
1.5 amoA基因的定量PCR检测
分别对AOB和AOA的amoA基因进行实时荧光定量PCR(real-time quantitative polymerase chain reaction, qPCR)检测, 使用FastStart Universal SYBRⓇ Green Master (ROX)试剂盒(Roche, Germany)和ABI 7500实时荧光定量PCR仪(Applied Biosystems, USA)进行分析测定, 扩增体系内添加0.20 μg ·μL-1牛血清蛋白[23](bovine serum albumin, BSA), 扩增程序如表 2所示.每组实验添加阴性对照, 扩增完成后增加熔解曲线分析, 确定样品扩增特异性.通过检测梯度稀释的含有AOB和AOA amoA基因的质粒标准品, 以amoA基因拷贝数的对数为横坐标, 以Ct值为纵坐标, 构建标准曲线.通过标准曲线换算得到样品中amoA基因拷贝数, 每个样品测定3个平行样.
2 结果与分析 2.1 AOB和AOA的amoA基因定量分析如1.4节所述, 通过琼脂糖凝胶电泳检测已确定扩增产物为amoA基因片段.采用qPCR技术对冬夏两季三亚河红树林表层沉积物中AOB和AOA的amoA基因进行定量检测, 结果如图 2所示.

|
图 2 冬夏两季AOB和AOA amoA基因丰度分布 Fig. 2 Abundance distribution of amoA genes of AOB and AOA during the winter and summer |
冬季AOB amoA基因丰度在0.092×104~7.115×104 copies ·g-1之间, 夏季则在0.303×104~6.677×104 copies ·g-1之间[图 2 (a)].在水平分布上, AOB amoA基因丰度沿河分布呈现出三亚河上游较高、中游较低和下游较高的特征, 由河向陆分布呈现出向河侧较高、红树林中心较低和向陆侧较高的特征; 不同季节中, AOB amoA基因丰度在多数采样点呈现出冬季高于夏季的特征.
冬季AOA amoA基因丰度在0.067×104~1.293×105 copies ·g-1之间, 夏季则在0.876×104~1.233×105 copies ·g-1之间[图 2 (b)].在水平分布上, AOA amoA基因丰度沿河分布特征与AOB一致, 由河向陆多数采样点呈现出与AOB相反的分布特征; 不同季节中, AOA amoA基因丰度在多数采样点呈现出夏季高于冬季的特征.
冬季AOA和AOB amoA基因丰度的比值在0.037~19.347之间, 夏季则在0.687~40.205之间, 多数采样点AOA amoA基因丰度高于AOB[图 2 (c)].在水平分布上, 红树林中心二者比值较高, 且AOA amoA基因丰度是AOB amoA基因丰度10倍以上的采样点均出现在红树林中心; 不同季节中, 二者比值在多数采样点呈现出夏季比冬季升高的特征, 其中SY3-2升高趋势最为明显.
2.2 环境因子对AOB和AOA分布的影响分析使用Canoco 5进行统计分析, 在各采样点AOB和AOA丰度的去趋势对应分析(detrended correspondence analysis, DCA)结果中第一轴的长度小于3, 故选择冗余分析(redundancy analysis, RDA)方法分别对冬夏两季各采样点好氧氨氧化微生物丰度和环境因子进行分析, 得到图 3、表 3和表 4所示的结果.

|
图 3 冬夏两季环境因子与AOB和AOA丰度的RDA分析 Fig. 3 RDA ordination of the relationships between environmental factors and the abundances of AOB and AOA during the winter and summer |
|
|
表 3 冬季环境因子与前两个排序轴间的相关系数 Table 3 Correlation coefficients of environmental factors with the first two axes of RDA during the winter |
|
|
表 4 夏季环境因子与前两个排序轴间的相关系数 Table 4 Correlation coefficients of environmental factors with the first two axes of RDA during the summer |
冬季AOB和AOA丰度与环境因子的冗余分析[图 3 (a)]结果显示, 排序轴1和排序轴2的解释度分别为61.08%和38.71%, 表明这两个排序轴基本能反映环境因子对AOB和AOA丰度的影响.结合环境因子与两个排序轴间的相关系数(表 3), DO与排序轴1的相关系数较高, pH和TOC与排序轴2的相关系数较高, 其中pH与AOB和AOA总体分布间达到了显著相关(P < 0.05), 总体而言, pH、TOC和DO是影响冬季三亚河红树林表层沉积物中AOB和AOA丰度的关键环境因子, 其中pH与AOB丰度具有较高的显著正相关性(P < 0.05), TOC与AOA丰度具有较高的正相关性.
夏季AOB和AOA丰度与环境因子的冗余分析结果显示[图 3 (b)], 排序轴1和排序轴2的解释度分别为60.45%和37.72%, 表明这两个排序轴基本能反映环境因子对AOB和AOA丰度的影响.结合环境因子与两个排序轴间的相关系数(表 4), 硝态氮和DO与排序轴1的相关系数较高, pH与排序轴2的相关系数较高, 其中硝态氮与AOB和AOA分布间达到了显著相关(P < 0.05), 总体而言, 硝态氮、DO和pH是影响夏季三亚河红树林表层沉积物中AOB和AOA丰度的关键环境因子, 其中pH与AOB丰度具有较高的正相关性, DO与AOA丰度具有较高的负相关性(图 3).
2.3 AOB和AOA潜在硝化速率及相对贡献分析通过模拟培养实验, 计算得到冬夏两季AOB和AOA的PNR(图 4)及二者对硝化作用的相对贡献率(图 5).

|
图 4 冬夏两季AOB和AOA的潜在硝化速率 Fig. 4 Potential nitrification rates of AOB and AOA during the winter and summer |

|
图 5 冬夏两季AOB和AOA对硝化作用的相对贡献率 Fig. 5 Relative contribution rates of AOB and AOA to nitrification during the winter and summer |
冬季AOB的PNR(以N计, 下同)在0.021~0.192 μmol ·(g ·d)-1之间, 夏季则在0.038~0.345 μmol ·(g ·d)-1之间[图 4 (a)].AOB的PNR在多数采样点沿三亚河呈现上游和下游较高、中游较低的特征, 由河向陆呈现向河侧较低、红树林中心和向陆侧较高的特征; 夏季AOB的PNR均高于冬季, 与乳山湾近海沉积物中[24]的季节变化趋势一致但数值较低.
冬季AOA的PNR在0.080~0.137 μmol ·(g ·d)-1之间, 夏季则在0.118~0.438 μmol ·(g ·d)-1之间[图 4 (b)].AOA的PNR沿三亚河多数采样点冬季的变化并不明显, 但夏季呈现与AOB相同的特征, 由河向陆冬季呈现与AOB相反的特征而夏季则表现出上游由河向陆逐渐增高且下游由河向陆逐渐降低的特征; 夏季AOA的PNR也均高于冬季, 与长江口潮滩湿地沉积物中[25]的季节变化趋势一致但数值也较低.
冬季AOB和AOA总的PNR在0.143~0.272 μmol ·(g ·d)-1之间, 夏季则在0.317~0.495 μmol ·(g ·d)-1之间[图 4 (c)].总PNR沿三亚河呈现冬季中游较低、上游和下游较高, 夏季下游较低、上游和中游较高的特征, 由河向陆冬季呈现上游红树林中心较高而下游红树林中心较低的特征, 夏季则均由河向陆增高; 夏季总的PNR明显高于冬季, 与长江口潮滩湿地沉积物中[25]的季节变化趋势一致且数值较低, 但与九龙江河口红树林未处理沉积物[10]中的数值近似一致.
冬季AOB对硝化作用的相对贡献率在14.81% ~70.73%之间, AOA则在29.27% ~85.19%之间[图 5 (a)]; 夏季AOB对硝化作用的相对贡献率在8.06% ~67.46%之间, AOA则在28.80% ~91.94%之间[图 5 (b)].在多数采样点冬季AOA对硝化作用的相对贡献率高于AOB而夏季AOB对硝化作用的相对贡献率高于AOA, 其中SY2-1冬夏两季AOA的相对贡献率均最高且高于AOB.
2.4 AOB和AOA amoA基因丰度与潜在硝化速率的相关性分析分别对冬夏两季好氧氨氧化微生物的潜在硝化速率与amoA基因丰度进行Pearson相关性分析(表 5), 结果表明AOB和AOA的潜在硝化速率与amoA基因丰度之间均不存在显著相关性(P>0.05).
|
|
表 5 冬夏两季潜在硝化速率与amoA基因丰度的相关性系数 Table 5 Correlation coefficients of potential nitrification rates and abundance of amoA genes during the winter and summer |
3 讨论 3.1 AOB和AOA的分布特征及影响其分布的关键环境因子
本研究中, 多数采样点AOA amoA基因丰度高于AOB, 这与Li等[8]在中国香港米埔红树林和Wang等[10]在九龙江河口红树林的研究结果相反, 而与Marcos等[7]在佛罗里达海岸红树林、Cao等[12]在受污染的红树林和张禹[9]在海南东寨港红树林的研究结果一致.AOA amoA基因丰度在红树林中心区域更高, 推测可能受植物根际影响更大, 这与Li等[8]在中国香港米埔红树林的研究结果相反而与Chen等[26]在水稻根际土壤中的研究结果一致, 已有研究表明, 植物能够通过根系将氧气输送至根区, 并在根际释放能够为微生物提供养分的分泌物, 使得植物根际的微生物总量和多样性都较高[27], 但植物也可能与氨氧化微生物竞争氧和氨氮而不能促进微生物生长, 考虑到本研究区域位于河流沿岸且红树林中还生长有半红树植物如黄槿等, 推测植物的生理生态特性与陆地中植物更为相似, 故根际效应也更为相似.多数采样点AOB amoA基因丰度冬季高于夏季, 与Wang等[11]在中国香港米埔红树林沉积物、陆诗敏等[28]在养殖池塘沉积物和Bernhard等[17]在河口沉积物中的研究结果一致, AOA amoA基因丰度则夏季高于冬季, AOA存在与AOB相反的时空分布特征, 这与苏瑜等[29]在我国北方土壤中的研究结果类似, 原因可能是受到环境因子变化的影响, AOA比AOB更适于在低氧、低pH和低氨环境中生长, 而AOB比AOA更能适应高氨环境, 当溶解氧、pH和氨氮这些环境因子变化时, AOB和AOA的数量随之出现相反的变化趋势, 此外本研究区域内作为反应底物的氧气质量浓度较低, AOB和AOA之间可能因底物限制存在竞争.有研究表明[30, 31]AOA amoA基因丰度与AOB amoA基因丰度的比值AOA/AOB可作为评价富营养化程度的生物指标, 在贫营养环境中AOA/AOB较高, 本研究中多数采样点冬季AOA/AOB更低, 即冬季三亚河红树林沉积物中富营养程度更高, 与冬季三亚河水质降低[32]的结果一致.三亚河红树林区域位于城市中心, 受人类活动影响较为明显, 12月属于三亚市旅游旺季, 旅游人口数量的大量增加导致住宿和餐饮排放的生活污水大量增加[32], 三亚河中大量的营养盐陆源输入使得红树林沉积物中营养盐含量升高.
DO和pH是影响AOB和AOA丰度的关键环境因子.此外, 冬季AOB和AOA的丰度受TOC的影响较大, 而夏季AOB和AOA的丰度受硝态氮的影响较大.氧气作为氨氧化反应的底物, 是影响好氧氨氧化微生物生长的重要环境因子, 分析图 3可以发现, DO与AOA丰度之间存在更高的负相关性, 即随DO降低AOA丰度升高的程度更高, 说明AOA更能适应低氧环境, 这与Pettridge等[33]在土壤中、Auguet等[34]在湖泊中和Caffrey等[35]在河口沉积物中的结果一致.已有研究表明, AOA比AOB对氧气有更高的亲和力, 故AOA比AOB更适合在低氧环境中生长[36], 本研究区域尤其在夏季DO普遍偏低, 这种环境条件更适于AOA生长.pH与AOB丰度之间存在较高的显著正相关性(P < 0.05), 即AOB丰度随pH降低而降低, 这与Cao等[12]在红树林沉积物和王超等[37]在子牙河水系的研究结果类似.已有研究表明, 环境中pH降低会导致作为反应底物的氨对氨氧化微生物的生物可利用性降低[38], 而AOB生长最适pH为7.0~8.5[39], 本研究区域较低的pH值可能使pH成为AOB生长的限制因子, 但AOA适宜生长的pH范围较宽(3.7~8.7)[40], 故受影响较小.冬季TOC与AOA丰度之间存在较高的正相关性, 即AOA丰度随TOC升高而升高, 这与苏瑜等[29]的研究结果一致.已有研究发现, AOA能够利用环境中的有机质进行生长, Wang等[41]的研究发现中国香港米埔红树林沉积物中的AOA能够利用烯丙基硫脲进行生长, Hallam等[42]则提出有机质能够促进AOA生长可能是因为AOA中存在完整的三羧酸循环途径关键基因, 具备异养生长的遗传基础.硝态氮作为硝化反应的产物, 在一定程度上会对氨氧化微生物的分布产生影响, 分析图 3可以发现夏季硝态氮与AOB和AOA的丰度之间存在较高的显著负相关性(P < 0.05), 即AOB和AOA丰度都随硝态氮升高而降低, 可能是随反应产物的增加而产生了抑制效应, 这与Li等[8]、Wang等[10]在红树林沉积物中和鲍林林等[43]在北运河沉积物中的研究结果类似.总的来说, 在本研究区域中AOA对环境的适应性比AOB更强, 二者在环境中的分布受到多个环境因子的共同作用.
3.2 AOB和AOA在不同季节的PNR和相对贡献率综合前述分析, 笔者发现夏季PNR均高于冬季, 这与郑艳玲[25]在长江口潮滩湿地的研究结果一致.对于大多数采样点来说, 冬季AOA对硝化作用的相对贡献率高于AOB, 说明冬季硝化作用以AOA为主导, 夏季AOB对硝化作用的相对贡献率高于AOA, 说明夏季硝化作用以AOB为主导, 这与He等[24]在乳山湾邻近海域沉积物中的研究结果一致.AOB和AOA的PNR及其对硝化作用的相对贡献率与amoA基因丰度的分布特征并不一致, 推测原因可能是基于DNA水平检测到的amoA基因不一定能够执行氨氧化功能, 或者是模拟培养实验的局限性, 如在振荡过程中可能破坏沉积物的结构[44], 添加反应底物的培养过程可能会使有些氨氧化微生物的活性产生滞后效应[45], 从而导致实验结果不够准确.总的来说, 本研究区域中硝化过程并不完全由AOB或者AOA主导进行.在已有研究中, 中国香港米埔红树林沉积物中AOB对硝化作用起主导作用[46]; 镇江古运河河岸沉积物中AOB对硝化过程起主导作用[47]; 俄勒冈州森林土壤中AOA对硝化过程起主导作用[48].这些研究表明不同环境条件下AOB和AOA在硝化作用中分别占据主导地位, 由此说明环境条件改变会影响AOB和AOA对硝化作用的相对贡献率.
进一步对冬夏两季AOB和AOA对硝化作用的相对贡献率与环境因子进行Pearson相关性分析, 结果表明冬季AOA对硝化作用的相对贡献率与NH4+-N存在显著负相关性(P < 0.05), 所以冬季在NH4+-N含量较低的环境中AOA是硝化作用的主要贡献者, 这与已有研究中AOB在NH4+-N含量较高的环境中是硝化作用主要贡献者[49, 50]的结果相对应.而夏季AOB对硝化作用的相对贡献率与环境因子之间不存在显著相关性(P>0.05).AOA对作为反应底物的氨的亲和力比AOB高[51], 但比AOB更易受高浓度氨抑制[52].在低氨环境中AOB因营养不足受到抑制而AOA仍能进行正常生理活动, 故AOA具有较高的硝化作用相对贡献率; 在高氨环境中AOA则因反应底物浓度过高而受到抑制, 此时AOB仍能进行正常生理活动, 故AOB具有较高的硝化作用相对贡献率.
3.3 AOB和AOA潜在硝化速率与amoA基因丰度的关系从表 5中可以发现AOB和AOA的潜在硝化速率与amoA基因丰度之间不存在显著相关性(P>0.05).与本研究结果类似的是, Wang等[10]和Zhang等[53]都在九龙江河口红树林沉积物中发现潜在硝化速率与amoA基因丰度不存在显著相关性, He等[24]在乳山湾邻近海域沉积物中同样发现潜在硝化速率与amoA基因丰度不存在显著相关性, 此外在河口沉积物[44]和农田土壤[54]中也有相一致的研究结果.总的来说, AOB和AOA的硝化活性与amoA基因丰度并不相关, 因为amoA基因的存在和数量并不能决定微生物活性或者基因功能的执行, 故对氨氧化活性应结合多种手段进行分析, 如DNA稳定同位素探测[55]和反转录定量PCR[44]等.
4 结论(1) 多数采样点中AOA amoA基因拷贝数高于AOB amoA基因拷贝数, 最高高出40倍.
(2) AOB和AOA丰度分布表现出明显的季节特征, 多数采样点中, AOB在冬季丰度较高, AOA则在夏季丰度较高, 冬季AOA和AOB amoA基因丰度的比值更低, 说明冬季三亚河红树林沉积物中富营养程度更高.
(3) 多种环境因子均可影响AOB和AOA的丰度分布, 其中DO、pH、TOC和硝态氮对AOB和AOA丰度的影响较大.
(4) AOB和AOA的潜在硝化速率夏季均高于冬季, 且AOB和AOA对硝化作用的相对贡献也存在明显的季节差异, 多数采样点中冬季AOA在硝化作用中占主导地位而夏季则是AOB在硝化作用中占主导地位, 冬季AOA对硝化作用的相对贡献率与NH4+-N存在显著负相关性, AOB和AOA的潜在硝化速率与amoA基因丰度均不存在显著相关性.
致谢: 感谢海南热带海洋学院赵怀宝老师、杜前进老师和王刚老师在样品采集和环境因子分析中给予的帮助.
| [1] |
石莉.中国红树林及其生态环境功能的综合分析与研究[D].青岛: 中国海洋大学, 2003. 1-11. Shi L. Comprehensive analysis of mangrove in China and its eco-environmental functions[D]. Qingdao: Ocean University of China, 2003. 1-11. http://cdmd.cnki.com.cn/Article/CDMD-10423-2003093427.htm |
| [2] |
林鹏. 中国红树林研究进展[J]. 厦门大学学报(自然科学版), 2001, 40(2): 592-603. Lin P. A review on the mangrove research in China[J]. Journal of Xiamen University (Natural Science), 2001, 40(2): 592-603. DOI:10.3321/j.issn:0438-0479.2001.02.050 |
| [3] | Junier P, Molina V, Dorador C, et al. Phylogenetic and functional marker genes to study ammonia-oxidizing microorganisms (AOM) in the environment[J]. Applied Microbiology and Biotechnology, 2010, 85(3): 425-440. |
| [4] | Chain P, Lamerdin J, Larimer F, et al. Complete genome sequence of the ammonia-oxidizing bacterium and obligate chemolithoautotroph Nitrosomonas europaea[J]. Journal of Bacteriology, 2003, 185(9): 2759-2773. DOI:10.1128/JB.185.9.2759-2773.2003 |
| [5] | Li J L, Nedwell D B, Beddow J, et al. AmoA gene abundances and nitrification potential rates suggest that benthic ammonia-oxidizing bacteria and not archaea dominate N cycling in the Colne Estuary, United Kingdom[J]. Applied and Environmental Microbiology, 2015, 81(1): 159-165. |
| [6] | Zhou X H, Zhang J P, Li Y M, et al. Distribution characteristics of ammonia oxidizing microorganisms in rhizosphere sediments of cattail[J]. Ecological Engineering, 2016, 88: 99-111. DOI:10.1016/j.ecoleng.2015.12.023 |
| [7] | Marcos M S, Barboza A D, Keijzer R M, et al. Tide as steering factor in structuring archaeal and bacterial ammonia-oxidizing communities in mangrove forest soils dominated by Avicennia germinans and Rhizophora mangle[J]. Microbial Ecology, 2018, 75(4): 997-1008. DOI:10.1007/s00248-017-1091-y |
| [8] | Li M, Cao H L, Hong Y G, et al. Spatial distribution and abundances of ammonia-oxidizing archaea (AOA) and ammonia-oxidizing bacteria (AOB) in mangrove sediments[J]. Applied Microbiology and Biotechnology, 2011, 89(4): 1243-1254. DOI:10.1007/s00253-010-2929-0 |
| [9] |
张禹.海南东寨港红树林湿地氨氧化古菌(AOA)多样性空间分布特征研究[D].海口: 海南大学, 2017. 42-49. Zhang Y. Diversity distribution of ammonia-oxidizing archaea from mangrove wetland system in Dongzhai Harbour, Hainan[D]. Haikou: Hainan University, 2017. 42-49. http://cdmd.cnki.com.cn/Article/CDMD-10589-1017173630.htm |
| [10] | Wang H T, Su J Q, Zheng T L, et al. Insights into the role of plant on ammonia-oxidizing bacteria and archaea in the mangrove ecosystem[J]. Journal of Soils and Sediments, 2015, 15(5): 1212-1223. DOI:10.1007/s11368-015-1074-x |
| [11] | Wang Y F, Feng Y Y, Ma X J, et al. Seasonal dynamics of ammonia/ammonium-oxidizing prokaryotes in oxic and anoxic wetland sediments of subtropical coastal mangrove[J]. Applied Microbiology and Biotechnology, 2013, 97(17): 7919-7934. DOI:10.1007/s00253-012-4510-5 |
| [12] | Cao H L, Li M, Hong Y G, et al. Diversity and abundance of ammonia-oxidizing archaea and bacteria in polluted mangrove sediment[J]. Systematic and Applied Microbiology, 2011, 34(7): 513-523. DOI:10.1016/j.syapm.2010.11.023 |
| [13] | Laanbroek H J, Keijzer R M, Verhoeven J T A, et al. Changes in community composition of ammonia-oxidizing betaproteobacteria from stands of black mangrove (Avicennia germinans) in response to ammonia enrichment and more oxic conditions[J]. Frontiers in Microbiology, 2013, 4: 343. |
| [14] |
丘耀文, 余克服. 海南红树林湿地沉积物中重金属的累积[J]. 热带海洋学报, 2011, 30(2): 102-108. Qiu Y W, Yu K F. Accumulation of heavy metals in sediment of mangrove wetland from Hainan Island[J]. Journal of Tropical Oceanography, 2011, 30(2): 102-108. DOI:10.3969/j.issn.1009-5470.2011.02.015 |
| [15] |
张敏, 赵牧秋, 车志伟, 等. 三亚河红树林沉积物酶活性时空动态研究[J]. 琼州学院学报, 2016, 23(2): 86-91. Zhang M, Zhao M Q, Che Z W, et al. Seasonal and spatial dynamics of sediment enzyme activities under Sanya River mangrove forest[J]. Journal of Qiongzhou University, 2016, 23(2): 86-91. |
| [16] |
王丽荣, 李贞, 蒲杨婕, 等. 海南东寨港、三亚河和青梅港红树林群落健康评价[J]. 热带海洋学报, 2011, 30(2): 81-86. Wang L R, Li Z, Pu Y J, et al. Health evaluation of mangrove community in Dongzhaigang, Sanyahe, and Qingmeigang mangrove nature reserves of Hainan province[J]. Journal of Tropical Oceanography, 2011, 30(2): 81-86. DOI:10.3969/j.issn.1009-5470.2011.02.012 |
| [17] | Bernhard A E, Tucker J, Giblin A E, et al. Functionally distinct communities of ammonia-oxidizing bacteria along an estuarine salinity gradient[J]. Environmental Microbiology, 2007, 9(6): 1439-1447. DOI:10.1111/j.1462-2920.2007.01260.x |
| [18] | Zheng Y L, Hou L J, Newell S, et al. Community dynamics and activity of ammonia-oxidizing prokaryotes in intertidal sediments of the Yangtze estuary[J]. Applied and Environmental Microbiology, 2014, 80(1): 408-419. |
| [19] |
贺惠, 甄毓, 米铁柱, 等. 乳山湾近海沉积物潜在硝化速率的测定[J]. 中国环境科学, 2017, 37(3): 1082-1088. He H, Zhen Y, Mi T Z, et al. Measurement of potential nitrification rates in sediments from adjacent waters of Rushan Bay[J]. China Environmental Science, 2017, 37(3): 1082-1088. |
| [20] |
刘天琳, 任佳琪, 王天佑, 等. 中性紫色水稻土硝化作用中细菌和古菌的相对贡献[J]. 土壤通报, 2018, 49(5): 1091-1096. Liu T L, Ren J Q, Wang T Y, et al. Relative contribution of bacteria and archaea to soil nitrification in neutral paddy soil[J]. Chinese Journal of Soil Science, 2018, 49(5): 1091-1096. |
| [21] | Rotthauwe J H, Witzel K P, Liesack W. The ammonia monooxygenase structural gene amoA as a functional marker:molecular fine-scale analysis of natural ammonia-oxidizing populations[J]. Applied and Environmental Microbiology, 1997, 63(12): 4704-4712. DOI:10.1128/AEM.63.12.4704-4712.1997 |
| [22] | Park S J, Park B J, Rhee S K. Comparative analysis of archaeal 16S rRNA and amoA genes to estimate the abundance and diversity of ammonia-oxidizing archaea in marine sediments[J]. Extremophiles, 2008, 12(4): 605-615. DOI:10.1007/s00792-008-0165-7 |
| [23] |
路兴岚, 甄毓, 米铁柱, 等. 沉积物中氨氧化细菌amoA基因荧光定量PCR检测方法的改进[J]. 中国海洋大学学报, 2012, 42(S1): 176-181. Lu X L, Zhen Y, Mi T Z, et al. Improvements of quantitative real-time PCR assay for amoA gene of aerobic ammonium oxidizing bacteria in sediments[J]. Periodical of Ocean University of China, 2012, 42(S1): 176-181. |
| [24] | He H, Zhen Y, Mi T Z, et al. Ammonia-oxidizing archaea and bacteria differentially contribute to ammonia oxidation in sediments from adjacent waters of Rushan Bay, China[J]. Frontiers in Microbiology, 2018, 9: 116. DOI:10.3389/fmicb.2018.00116 |
| [25] |
郑艳玲.长江口潮滩湿地氨氧化菌群动态及活性研究[D].上海: 华东师范大学, 2015. 133-136. Zheng Y L. Aerobic/anaerobic ammonium oxidation microbial dynamics, and associated activity in intertidal wetlands of the Yangtze Estuary[D]. Shanghai: East China Normal University, 2015. 133-136. |
| [26] | Chen X P, Zhu Y G, Xia Y, et al. Ammonia-oxidizing archaea:important players in paddy rhizosphere soil?[J]. Environmental Microbiology, 2008, 10(8): 1978-1987. DOI:10.1111/j.1462-2920.2008.01613.x |
| [27] | Münch C, Kuschk P, Röske I. Root stimulated nitrogen removal:only a local effect or important for water treatment?[J]. Water Science & Technology, 2005, 51(9): 185-192. |
| [28] |
陆诗敏.淡水养殖池塘环境中氨氧化微生物的研究[D].武汉: 华中农业大学, 2014. 24-36. Lu S M. Study on the ammonia-oxidizing microorganisms in the freshwater aquaculture pond environment[D]. Wuhan: Huazhong Agricultural University, 2014. 24-36. |
| [29] |
苏瑜, 王为东. 我国北方四类土壤中氨氧化古菌和氨氧化细菌的活性及对氨氧化的贡献[J]. 环境科学学报, 2017, 37(9): 3519-3527. Su Y, Wang W D. Activity of AOA and AOB and their contributions to ammonia oxidization in four soils in North China[J]. Acta Scientiae Circumstantiae, 2017, 37(9): 3519-3527. |
| [30] | Sims A, Horton J, Gajaraj S, et al. Temporal and spatial distributions of ammonia-oxidizing archaea and bacteria and their ratio as an indicator of oligotrophic conditions in natural wetlands[J]. Water Research, 2012, 46(13): 4121-4129. DOI:10.1016/j.watres.2012.05.007 |
| [31] |
刘正辉.东江氨氮污染河段的微生物群落特征[D].广州: 华南理工大学, 2011. 105-107. Liu Z H. Microbial community characteristics in the water column contaminated with ammonia-nitrogen along the Dongjiang River[D]. Guangzhou: South China University of Technology, 2011. 105-107. |
| [32] |
陈攀, 李晓梅, 王梅, 等. 旅游旺季三亚河水体中有机物含量的分析[J]. 琼州学院学报, 2014, 21(5): 93-98. Chen P, Li X M, Wang M, et al. Analysis of organic matters content in water of Sanya River in tourist season[J]. Journal of Qiongzhou University, 2014, 21(5): 93-98. |
| [33] | Pett-Ridge J, Petersen D G, Nuccio E, et al. Influence of oxic/anoxic fluctuations on ammonia oxidizers and nitrification potential in a wet tropical soil[J]. FEMS Microbiology Ecology, 2013, 85(1): 179-194. |
| [34] | Auguet J C, Triadó-Margarit X, Nomokonova N, et al. Vertical segregation and phylogenetic characterization of ammonia-oxidizing archaea in a deep oligotrophic lake[J]. The ISME Journal, 2012, 6(9): 1786-1797. DOI:10.1038/ismej.2012.33 |
| [35] | Caffrey J M, Bano N, Kalanetra K, et al. Ammonia oxidation and ammonia-oxidizing bacteria and archaea from estuaries with differing histories of hypoxia[J]. The ISME Journal, 2007, 1(7): 660-662. DOI:10.1038/ismej.2007.79 |
| [36] | Abell G C J, Banks J, Ross D J, et al. Effects of estuarine sediment hypoxia on nitrogen fluxes and ammonia oxidizer gene transcription[J]. FEMS Microbiology Ecology, 2011, 75(1): 111-122. |
| [37] |
王超, 单保庆. 子牙河水系水和沉积物好氧氨氧化微生物分布特征[J]. 环境科学学报, 2012, 32(12): 2943-2950. Wang C, Shan B Q. The distribution of aerobic ammonia oxidizing microorganisms in Ziya River, Haihe Basin[J]. Acta Scientiae Circumstantiae, 2012, 32(12): 2943-2950. |
| [38] | Burton S A Q, Prosser J I. Autotrophic ammonia oxidation at low pH through urea hydrolysis[J]. Applied and Environmental Microbiology, 2001, 67(7): 2952-2957. DOI:10.1128/AEM.67.7.2952-2957.2001 |
| [39] |
郝永俊, 吴松维, 吴伟祥, 等. 好氧氨氧化菌的种群生态学研究进展[J]. 生态学报, 2007, 27(4): 1573-1582. Hao Y J, Wu S W, Wu W X, et al. Research progress on the microbial ecology of aerobic ammonia-oxidizing bacteria[J]. Acta Ecologica Sinica, 2007, 27(4): 1573-1582. DOI:10.3321/j.issn:1000-0933.2007.04.038 |
| [40] | Erguder T H, Boon N, Wittebolle L, et al. Environmental factors shaping the ecological niches of ammonia-oxidizing archaea[J]. FEMS Microbiology Reviews, 2009, 33(5): 855-869. DOI:10.1111/j.1574-6976.2009.00179.x |
| [41] | Wang Y F, Li X Y, Gu J D. Differential responses of ammonia/ammonium-oxidizing microorganisms in mangrove sediment to amendment of acetate and leaf litter[J]. Applied Microbiology and Biotechnology, 2014, 98(7): 3165-3180. DOI:10.1007/s00253-013-5318-7 |
| [42] | Hallam S J, Mincer T J, Schleper C, et al. Pathways of carbon assimilation and ammonia oxidation suggested by environmental genomic analyses of marine Crenarchaeota[J]. PLoS Biology, 2006, 4(4): e95. DOI:10.1371/journal.pbio.0040095 |
| [43] |
鲍林林, 陈永娟, 王晓燕. 北运河沉积物中氨氧化微生物的群落特征[J]. 中国环境科学, 2015, 35(1): 179-189. Bao L L, Chen Y J, Wang X Y. Diversity and abundance of ammonia-oxidizing prokaryotes in surface sediments in Beiyun River[J]. China Environmental Science, 2015, 35(1): 179-189. |
| [44] | Bernhard A E, Bollmann A. Estuarine nitrifiers:new players, patterns and processes[J]. Estuarine, Coastal and Shelf Science, 2010, 88(1): 1-11. DOI:10.1016/j.ecss.2010.01.023 |
| [45] |
张秋芳, 徐继荣, 苏建强, 等. 河口生态系统氨氧化菌生态学研究进展[J]. 生态学报, 2012, 32(18): 5878-5888. Zhang Q F, Yu J R, Su J Q, et al. Research progress on ammonia-oxidizing microorganisms in estuarine ecosystem[J]. Acta Ecologica Sinica, 2012, 32(18): 5878-5888. |
| [46] | Li M, Gu J D. Community structure and transcript responses of anammox bacteria, AOA, and AOB in mangrove sediment microcosms amended with ammonium and nitrite[J]. Applied Microbiology and Biotechnology, 2013, 97(22): 9859-9874. DOI:10.1007/s00253-012-4683-y |
| [47] | Zhou X H, Li Y M, Zhang J P, et al. Diversity, abundance and community structure of ammonia-oxidizing archaea and bacteria in riparian sediment of Zhenjiang ancient canal[J]. Ecological Engineering, 2016, 90: 447-458. DOI:10.1016/j.ecoleng.2016.01.068 |
| [48] | Lu X D, Bottomley P J, Myrold D D. Contributions of ammonia-oxidizing archaea and bacteria to nitrification in Oregon forest soils[J]. Soil Biology and Biochemistry, 2015, 85: 54-62. DOI:10.1016/j.soilbio.2015.02.034 |
| [49] | Di H J, Cameron K C, Shen J P, et al. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils[J]. Nature Geoscience, 2009, 2(9): 621-624. DOI:10.1038/ngeo613 |
| [50] | Norman J S, Barrett J E. Substrate and nutrient limitation of ammonia-oxidizing bacteria and archaea in temperate forest soil[J]. Soil Biology and Biochemistry, 2014, 69: 141-146. DOI:10.1016/j.soilbio.2013.11.003 |
| [51] | Lehtovirta-Morley L E, Stoecker K, Vilcinskas A, et al. Cultivation of an obligate acidophilic ammonia oxidizer from a nitrifying acid soil[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(38): 15892-15897. DOI:10.1073/pnas.1107196108 |
| [52] | Prosser J I, Nicol G W. Archaeal and bacterial ammonia-oxidisers in soil:the quest for niche specialisation and differentiation[J]. Trends in Microbiology, 2012, 20(11): 523-531. DOI:10.1016/j.tim.2012.08.001 |
| [53] | Zhang Q F, Peng J J, Chen Q, et al. Impacts of Spartina alterniflora invasion on abundance and composition of ammonia oxidizers in estuarine sediment[J]. Journal of Soils and Sediments, 2011, 11(6): 1020-1031. DOI:10.1007/s11368-011-0369-9 |
| [54] | Jia Z J, Conrad R. Bacteria rather than archaea dominate microbial ammonia oxidation in an agricultural soil[J]. Environmental Microbiology, 2009, 11(7): 1658-1671. DOI:10.1111/j.1462-2920.2009.01891.x |
| [55] |
阎佳, 胡勇有. 2类氨氧化菌的生态特性及其应用研究进展[J]. 华南师范大学学报(自然科学版), 2016, 48(4): 1-9. Yan J, Hu Y Y. Eco-physiology and research progress of two ammonium oxidizing microbes[J]. Journal of South China Normal University (Natural Science Edition), 2016, 48(4): 1-9. |