 2020, Vol. 41
2020, Vol. 41



2. 甘肃省轻工研究院, 兰州 730030;
3. 山东同济测试科技股份有限公司, 烟台 264000;
4. 烟台大学环境与材料工程学院, 烟台 264005
2. Gansu Province Light Industry Research Institute, Lanzhou 730030, China;
3. Shandong Tongji Testing Technology Co., Ltd., Yantai 264000, China;
4. School of Environment and Material Engineering, Yantai University, Yantai 264005, China
温度是影响细菌活性的主要因素之一.温度过低细胞膜呈凝胶状, 营养物质跨膜运输受阻, 细胞会停止生长.超过温度上限时, 对温度敏感的细胞组成开始变性, 细胞生长难以维持甚至死亡[1, 2].硝化细菌属于中温细菌, 对温度变化较为敏感[3].顾升波等[4]的研究发现在11~16℃下, 低温对两种硝化菌活性均存在不利影响, 但对氨氧化细菌(AOB)的抑制作用大于对亚硝酸盐氧化细菌(NOB)的抑制作用. Gabarró等[5]将温度控制在10~15℃条件下, 发现出水NO3--N浓度升高, 半亚硝化系统稳定运行遭到破坏, 证实了NOB的活性略高于AOB.但于莉芳等[6]在25℃和30℃条件研究发现随温度升高, AOB活性增强而NOB活性变化则恰好相反, 因而AOB成为系统实现短程硝化的优势菌种.目前探究温度对硝化细菌的影响主要集中于AOB和NOB共存的硝化系统中, 而对于研究AOB受到抑制的硝化系统中微生物菌群结构影响却鲜见报道.
近几年, 先进的高通量测序技术被广泛用于城市污水处理厂活性污泥微生物群落结构的研究.其具有通量高、测序快、准确度高等特点.同时能定性分析微生物菌群的多样性、群落结构及功能, 对城市污水处理厂活性污泥微生物优势菌群确定和群落相对完整性的研究有重要意义[7~9].
本试验考察了高温冲击对亚硝酸盐氧化过程中微生物菌群结构的影响, 该系统经过前期富集阶段已抑制AOB的生长, 并着重考察了高温对亚硝酸盐氧化过程中主要菌属(NOB)及系统内菌群结构变化的影响, 以期为应对实际工程中出现的水温过高突发状况提供可靠的理论依据.此外, 本研究采用Illumina MiSeq第二代测序方法探究高温冲击对亚硝酸盐氧化过程菌群结构的影响, 比较温度变化下微生物菌群结构的差异性, 同时量化分析主要脱氮微生物的菌群丰富度和多样性, 以深入研究温度对亚硝酸盐氧化过程中微生物的影响.
1 材料与方法 1.1 试验用水及接种污泥本试验用水为人工模拟废水, 包括:0.4 g NaHCO3、0.9 g KH2PO4、1.2 g K2HPO4及5 mL微量元素母液(成分为:1.25 g EDTA、0.55 g ZnSO4 ·7H2 O、0.4 g CoCl2 ·6H2 O、1.275 g MnCl2 ·4H2 O、0.4 g CuSO4 ·4H2 O、0.05 g Na2MoO4 ·2H2 O、1.375 g CaCl2 ·2H2 O、1.25 g FeCl3 ·6H2 O、44.4 g MgSO4 ·7H2 O), 每周期进水NO2--N浓度由计算量投加NaNO2.试验接种污泥取自兰州市西固区兰炼污水处理厂氧化沟好氧段污泥, 具有良好生物脱氮性能, 驯化后污泥MLVSS=(1 490±30)mg ·L-1.
1.2 试验装置及运行方案本试验采用序批式反应器(sequencing batch reator, SBR), 有效容积3 L, 排水比1 :3.试验考察温度梯度分别为:25、35、38和40℃.为了不使pH和DO成为限制因素, 控制两者浓度分别为7.5±0.1和4.0~4.5 mg ·L-1.每周期反应器运行方式为:进水(1 min)、硝化(通过实时监测DO值控制硝化终点)、沉淀(30 min)、排水(5 min).通过观察比亚硝态氮氧化速率以判断硝化细菌反应活性, 其按公式(1)计算.
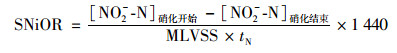
|
(1) |
式中, SNiOR为比亚硝态氮氧化速率[g ·(g ·d)-1, 以N/VSS计]; [NO2--N]硝化开始为曝气开始时NO2--N浓度(mg ·L-1); [NO2--N]硝化结束为曝气结束时NO2--N浓度(mg ·L-1); MLVSS为污泥浓度(mg ·L-1); tN为曝气反应时间(min).
1.3 水质分析项目及测定方法进出水NO2--N浓度及可挥发性悬浮固体(MLVSS)浓度采用国家标准方法测定[10], 温度、pH值及DO均采用WTW Multi-3420多参数测定仪及相应检测探头进行实时监测.
1.4 微生物高通量测序分别取25℃(第67周期)、35℃(第87周期)及40℃(第97周期)活性污泥样品(每个温度包括3个平行样品), 通过冷冻干燥(-80℃)处理后保存, 利用DNA提取试剂盒(DNeasyPowerSoil Kit)提取样品的总DNA.采用紫外分光光度计对DNA进行定量, 并通过0.8%琼脂糖凝胶电泳检测DNA提取质量.以3组样品原液作为PCR模板, 利用16S rRNA通用引物进行V4和V5区扩增, 所用引物为515F(GTGCCAGCMGCCGCGGTAA)和907R(CCGT CAATTCMTTTRAGTTT).PCR扩增反应条件为:预加热95℃, 2 min; 随后进行25周期扩增反应(变性95℃, 30s;退火55℃, 30 s; 延伸72℃, 40 s); 终止延伸72℃, 10 min.反应结束后将PCR扩增产物通过2%琼脂糖凝胶电泳进行检测, 并对目标片段进行切胶回收.DNA扩增样品利用Illumina MiSeq测序仪进行上机测序.
1.5 生物信息学分析采用QIIME软件对原始数据进行聚类分析, 主要步骤为双末端序列装配、去除低质量序列、去重复, 运用Mothur软件去嵌合体, 得到优质序列.对优质序列按97%的相似性作为一个单元来划分, 进行OTU(operational taxonomic unit)分类学鉴定.利用Mothur软件中的summary.single命令, 计算Chao1、ACE、Simpson和Shannon多样性指数.计算各OTUs在3组样品中的相对丰度并分别在门、属水平上进行汇总, 使用R语言, 对相对丰度大于5%的属进行聚类分析并绘制热图.
2 结果与讨论 2.1 反应器运行状况分析为了进一步研究高温冲击对亚硝酸盐氧化过程的影响, 本试验共进行2个阶段.如图 1所示, 阶段Ⅰ在25℃条件下对活性污泥进行驯化, 经过67个周期的运行, 通过逐步提升NO2--N浓度联合高溶解氧成功实现亚硝酸盐氧化过程中主要菌属(NOB)的富集培养.SNiOR(以N/VSS计, 下同)从开始时的0.01g ·(g ·d)-1提高到0.46 g ·(g ·d)-1.阶段Ⅱ通过逐步升温考察高温冲击对系统的影响, 分别在35、38和40℃进行了30个周期的批次试验, 可以看出高温冲击对亚硝酸盐氧化过程产生明显抑制作用:系统SNiOR在35℃达到峰值后随即开始逐渐降低, 从1.04 g ·(g ·d)-1降为0.29 g ·(g ·d)-1.因此, 本试验分别在25、35和40℃反应末期取活性污泥样品, 进行高通量测序拟从分子生物学角度探究高温冲击对亚硝酸盐氧化过程中菌群结构的影响.

|
图 1 SBR系统内NO2--N浓度和SNiOR变化规律 Fig. 1 Variations of NO2--N concentration and SNiOR in the SBR system |
利用高通量测序技术, 经过序列过滤与去除嵌合体序列, 3种温度下活性污泥样品得到可用于后续分析的优质序列数共计326 392条, 优质序列长度分布在387~395 nt之间, 其中以长度为393 nt序列居多.进行OTU聚类分析, 统计各样品在不同OTU中的丰度信息, 3组样品共产生4 923条OTU, 其中25、35和40℃特有的OTU数目(图 2)分别为1 341、814和587.25℃样品特有OTU数目最多, 预示含有较多特有的细菌种类.25℃与35℃共有OTU数目较多, 为714条.对OTU利用QIIME软件生成从门到属依次进行分类, 结果见表 1.结合图 3丰度等级曲线分析, 25℃和35℃样品的等级丰度曲线在横轴上的长度都比较长, 且下降较为平滑, 表明这两个温度所含群落组成的均匀度较好, 群落丰度差异较小.40℃样品的等级丰度曲线则在横轴上的长度都比较短, 且快速下降, 表明该温度下所含物种均匀度较低.

|
图 2 OTU分布维恩图 Fig. 2 Venn diagram showing the distribution of OTUs |

|
图 3 OTU丰度等级曲线 Fig. 3 OTU rank-abundance distribution curves of samples diagrams showing the distribution of OTUs |
|
|
表 1 硝化细菌16S rRNA序列及丰富度 Table 1 Pearson's correlation coefficient of sequence and abundance of bacterial 16S rRNA in nitrifying bacteria |
利用α多样性分析样品微生物群落的丰富度和多样性.各样品的丰富度指数(Chao1)、均匀度指数(ACE)和多样性指数(Simpson、Shannon), 如表 2所示.
|
|
表 2 多样性指数统计 Table 2 The α diversity indices |
就物种丰富度而言, 可以看出各温度样品中菌群丰富度Chao1数值[11]表现为减小的趋势, 表明随温度的升高系统内菌群丰富度逐渐下降.ACE指数变化显示随温度升高系统内菌群均匀度下降, 这表明高温条件下系统内菌群中某些细菌种群发育较好, 在群落中的优势度上升; 可推测本系统中优势菌群的变化与菌群丰富度的变化有关, 高温环境可能为某些菌群提供良好的生长繁殖环境, 使其成为系统内的优势菌种.但高温冲击明显降低了系统内菌群的丰富度以及均匀度.
结合Shannon和Simpson指数, 25℃和35℃样品Shannon指数(8.24和8.16)明显高于40℃样品(5.98), Simpson指数(0.98和0.97)也高于40℃(0.87).由此可见, 25℃和35℃条件下系统内菌群多样性要高于40℃条件, 进一步证实了随着温度的升高菌群的多样性逐渐降低[12].
根据样品间微生物进化信息及丰度信息进行Unifrac分析, 计算得出样品差异距离矩阵, 并在距离矩阵的基础上采用非度量多维尺度法(nonmetric multidimensional scaling, NMDS)进行beta多样性分析.基于Unifrac加权NMDS(图 4)分析, 由于同时计算了样品的进化信息及丰度信息, 各温度样品点虽能大致实现组内聚类, 样品点之间均保持一定的距离, 无重叠现象; 但仍存在距离差异较明显的样点(25-2和40-1).由此可知高温冲击条件下各样品中菌群群落由于进化信息和丰度信息的不同而产生显著差异, 该差异主要由微生物不同的最适生长条件所决定[13].基于Unifrac非加权NMDS分析可知(图 4), 相同温度条件下平行样品点与点间的距离较小, 能很好地聚为一类, 说明所取相同温度条件下样品组内菌群群落结构在进化关系上相似度高, 具有较好的代表性; 而不同温度条件下组间样品点与点间距离较大, 表明温度变化可导致系统内菌群群落结构在进化关系上产生明显差异.

|
图 4 样品群落结构Unifrac无度量多维尺度排序 Fig. 4 Unifrac nonmetric multidimensional scaling of the samples |
高温冲击条件下亚硝酸盐氧化过程微生物群落在门水平上的组成如图 5所示, 其中检测出相对丰度大于1%的菌门, 分别有10种(25℃)、9种(35℃)及9种(40℃), 可检测到微生物总量关系为:97.59%(25℃)>96.69%(35℃)>95.46%(40℃).3组样品中群落组成百分比较高的菌门相对比例如图 5所示, 主要包括Proteobacteria、Bacteroidetes、Nitrospirae、Chloroflexi、Gemmatimonadetes、Planctomycetes、Deinococcus、Armatimonadetes、Acidobacteria和Firmicutes等.其中Proteobacteria、Bacteroidetes、Nitrospirae和Chloroflexi在样品中所占百分比最高, 4种菌门总和在3组样品中的相对比例均占80%以上.

|
图 5 门水平细菌群落结构及分布 Fig. 5 Bacterial community structure and distribution of the samples at the phylum level |
大量研究表明:具有硝化功能的微生物菌属大多属于Proteobacteria和Nitrospirae, 如NOB中Nitrospira属于Nitrospirae, AOB中Nitrosomonas、Nitrosococcus和NOB中的Nitrobacter、Nitrococcus和Nitrotoga均属于Proteobacteria[14~16].因此, 本研究主要富集NOB的3组样品中Proteobacteria和Nitrospirae所占百分比较高, 但其比例在样品组间却有所差异.Proteobacteria是细菌中最大的一门, 同时也是污水处理中微生物门水平上的主导门类[17].
在高温冲击的条件下, 40℃时Proteobacteria占比增大, 主要由于高温对微生物具有抑制作用, 使能够适应高温的Proteobacteria占据优势并得以增殖.Nitrospirae作为亚硝酸盐氧化过程中的主导微生物菌门, 在不同温度条件下相对丰度存在一定的差异:14.03%(25℃)>13.86%(35℃)>7.97%(40℃), 随着温度的上升, Nitrospirae相对丰度逐渐下降, 但在25℃和35℃条件下样品丰度差异较小.这说明当温度大于35℃时, Nitrospirae受抑制程度较明显.
另外, 在关于活性污泥反应器生物菌群结构相关报道中, Bacteroidetes和Chloroflexi也经常被检测到, 但占总菌门相对丰度较低, 约为1%[18~20], 同时所检测的样品大多为处理实际城市生活污水的活性污泥.然而本试验活性污泥样品是用来处理底物较为单一且成分简单的人工配水, 蓝色框表示硝化菌导致Bacteroidetes和Chloroflexi在研究中丰度较高.Bacteroidetes在25℃和35℃丰度差异较小, 在40℃时丰度下降明显.但在高温冲击35℃时, Bacteroidetes丰度呈现上升趋势, 说明35℃高温冲击有利于Bacteroidetes生长.Chloroflexi在高温冲击条件下, 丰度占比呈现明显下降趋势, 说明高温不利于该菌门的生长.由此可见, 高温冲击条件对亚硝酸盐氧化过程微生物群落的门水平菌群结构影响存在差异.
2.3.2 属水平微生物群落分布特征为了更深入了解高温冲击条件对亚硝酸盐氧化过程微生物菌群结构的影响, 本研究还从属水平上对3组样品进行聚类分析及比较.将相对丰度前20位的微生物菌属以及具有硝化功能的微生物菌属进行相似性聚类分析, 并绘制属水平热图(图 6), 通过颜色阶梯及相似程度来反映不同温度条件下活性污泥样品在属分类水平上群落组成的相似性和差异性[21].

|
图 6 属水平热图及聚类分析 Fig. 6 Heatmap and clustering tree analysis of dominant genera |
对于系统中的硝化菌属, 由图 7可知, 在高温冲击下的亚硝酸盐氧化系统中, 均检测到了两类氨氧化细菌(Nitrosomonas和Nitrosospira)和两类亚硝酸盐氧化细菌(Nitrospira和Nitrobacter), 但检测到的AOB相对丰度极低(相对丰度均<0.05%), 进一步表明该硝化系统活性污泥中仅含有NOB.

|
图 7 属水平细菌群落丰度柱状图 Fig. 7 Bacterial community abundance histogram of the samples at the genus level |
作为3组样品中占绝对优势的NOB菌, 属于Nitrospirae的Nitrospira[22], 其相对丰度为:15.73%(35℃)>14.20%(25℃)>8.37%(40℃).由图 6可知, 35-1和35-3样品中Nitrospira含量远大于其他样品, 同时结合亚硝酸盐氧化过程中系统SNiOR可以看出, 在35℃达到峰值为1.04 g ·(g ·d)-1, 说明高温冲击条件在35℃时有利于Nitrospira的生长.这与姚倩等[23]的研究结果一致:运用SBR反应器及人工配水进水研究发现, 当温度变化范围在15~35℃时, Nitrospira的活性随着温度的上升而不断增大, 35℃左右时Nitrospira的活性达到最大值.但当温度达到40℃时, Nitrospira的相对丰度减少一半, 其对应该温度下的SNiOR也降为0.29g ·(g ·d)-1, 活性降低70%.在检测到的另一种NOB菌, 属于Proteobacteria的Nitrobacter, 其相对丰度为:0.10%(25℃)>0.04%(35℃)>0.02%(40℃).由于含量过低, 可认为Nitrospira为该系统内亚硝酸盐氧化过程中的主要硝化菌属.
另外, 属水平上不同温度条件下, 样品的微生物系统中还存在2个优势菌属(相对丰度大于5%), Halomonas和Pseudomonas.Halomonas在高温冲击条件下, 相对丰度呈现逐渐上升的趋势, 40℃时在系统中大量富集, 推测是高温条件下混合液蒸发致使系统中盐度增加, 适宜Halomonas生长[24, 25]; Pseudomonas是专性好氧菌, 具有好氧反硝化功能[26].高温冲击条件均有利于这两种菌属的生长.
综上所述, 高温冲击条件会导致亚硝酸盐氧化过程中微生物菌群结构的差异性变化.由此可见, 温度突变是导致实际污水处理系统菌群结构变化的重要因素之一.同时, 本试验证明了作为NOB中的主要菌属, Nitrospira菌属可以抵抗35℃高温冲击, 同时在25~35℃的温度区间内, 该菌属相对丰度能够逐步提升, 系统内SNiOR可以达到峰值, NOB活性达到最强.而对于实际脱氮污水处理系统来说, 亚硝酸盐氧化过程是生物脱氮的一个主要环节, 因此可以通过适当提高温度优化系统亚硝态氮去除效果.在实际工程中如遇35℃以下的高温冲击情况, 亚硝酸盐氧化过程不会受到冲击影响.
3 结论(1) 本研究无论从微生物群落丰富度、多样性以及组成差异分析, 均表明高温冲击对亚硝酸盐氧化过程活性污泥微生物菌群造成了明显的结构差异.
(2) 本研究高温冲击试验表明, 随温度的升高亚硝酸盐氧化过程系统内菌群丰富度和均匀度逐渐下降, 同时多样性也逐渐降低.不同温度各样品组间菌群群落由于进化信息和丰度信息的不同而产生显著差异, 该差异主要由微生物不同的最适生长条件所决定; 但相同温度各样品组内菌群群落结构在进化关系上相似度高, 具有较好的代表性.
(3) 本研究活性污泥样品中的NOB隶属于Nitrospirae的Nitrospira, 当温度在25~35℃条件下该菌属具有良好的硝化活性; 当温度达到40℃时, 其活性降低70%.
| [1] | Ruiz-Hernando M, Martín-Díaz J, Labanda J, et al. Effect of ultrasound, low-temperature thermal and alkali pre-treatments on waste activated sludge rheology, hygienization and methane potential[J]. Water Research, 2014, 61: 119-129. |
| [2] | Guo J B, Fu X, Baquero A, et al. Trade-off between carbon emission and effluent quality of activated sludge processes under seasonal variations of wastewater temperature and mean cell retention time[J]. Science of the Total Environment, 2016, 547: 331-344. |
| [3] | 郑平, 徐向阳, 胡宝兰, 等. 新型生物脱氮理论与技术[M]. 北京: 科学出版社, 2004. |
| [4] |
顾升波, 王淑莹, 彭永臻. 短程深度脱氮中试工艺的低温启动和维持[J]. 环境科学, 2013, 34(8): 3164-3170. Gu S B, Wang S Y, Peng Y Z. Start-up and maintenance method for short-cut advanced nitrogen removal process in a pilot-scale reactor at low temperature[J]. Environmental Science, 2013, 34(8): 3164-3170. |
| [5] | Gabarró J, Ganigué R, Gich F, et al. Effect of temperature on AOB activity of a partial nitritation SBR treating landfill leachate with extremely high nitrogen concentration[J]. Bioresource Technology, 2012, 126: 283-289. |
| [6] |
于莉芳, 傅学焘, 杨秀玲, 等. 2种温度下污泥水富集硝化菌的对比[J]. 环境工程学报, 2018, 12(1): 102-109. Yu L F, Fu X T, Yang X L, et al. Comparison of nitrifier accumulation in sludge liquor treatment system at two different temperatures[J]. Chinese Journal of Environmental Engineering, 2018, 12(1): 102-109. |
| [7] |
曾涛涛, 蒋小梅, 韩科昌, 等. 生活污水处理厂微生物群落结构解析[J]. 安全与环境学报, 2018, 18(2): 697-703. Zeng T T, Jiang X M, Han K C, et al. Analysis of microbial community constituent composition of some sewage treatment and processing plant[J]. Journal of Safety and Environment, 2018, 18(2): 697-703. |
| [8] |
宋云龙, 张金松, 朱佳, 等. 基于高通量测序的微生物强化污泥减量工艺中微生物群落解析[J]. 中国环境科学, 2016, 36(7): 2099-2107. Song Y L, Zhang J S, Zhu J, et al. Analysis of microbial community in in-situ sludge reduction process by bioaugmentation using high-throughput sequencing technology[J]. China Environmental Science, 2016, 36(7): 2099-2107. |
| [9] |
彭永臻, 钱雯婷, 王琦, 等. 基于宏基因组的城市污水处理厂生物脱氮污泥菌群结构分析[J]. 北京工业大学学报, 2019, 45(1): 95-102. Peng Y Z, Qian W T, Wang Q, et al. Unraveling microbial structure of activated sludge in a full-scale nitrogen removal plant using metagenomic sequencing[J]. Journal of Beijing University of Technology, 2019, 45(1): 95-102. |
| [10] | 国家环境保护局.水和废水监测分析方法[M]. (第四版).北京: 中国环境科学出版社, 2002. 210-285. |
| [11] | Chao A N, Shen T J. Nonparametric prediction in species sampling[J]. Journal of Agricultural, Biological, and Environmental Statistics, 2004, 9(3): 253-269. |
| [12] |
于雪, 孙洪伟, 李维维, 等. 温度对硝化杆菌(Nitrobacter)活性动力学影响[J]. 环境科学, 2019, 40(3): 1426-1430. Yu X, Sun H W, Li W W, et al. Effect of temperature on the activity kinetics of nitrobacter[J]. Environmental Science, 2019, 40(3): 1426-1430. |
| [13] |
吴鹏, 陆爽君, 徐乐中, 等. 温度对ABR-MBR复合工艺处理生活污水的影响及其微生物群落分析[J]. 环境科学, 2014, 35(9): 3466-3472. Wu P, Lu S J, Xu L Z, et al. Effects of temperature on combined process of ABR and MBR for domestic sewage treatment and analysis of microbial community[J]. Environmental Science, 2014, 35(9): 3466-3472. |
| [14] | Ida T, Kugimiya M, Takahashi R, et al. Phylogenetic relationships among ammonia-oxidizing bacteria as revealed by gene sequences of glyceraldehyde 3-phosphate dehydrogenase and phosphoglycerate kinase[J]. Journal of Bioscience and Bioengineering, 2005, 99(6): 569-576. |
| [15] | Nogueira R, Melo L F. Competition between Nitrospira spp. and Nitrobacter spp. in nitrite-oxidizing bioreactors[J]. Biotechnology and Bioengineering, 2006, 95(1): 169-175. |
| [16] | Marin B, Nowack E C, Glöckner G, et al. The ancestor of the Paulinella chromatophore obtained a carboxysomal operon by horizontal gene transfer from a Nitrococcus-like γ-proteobacterium[J]. BMC Evolutionary Biology, 2007, 7(1): 85. |
| [17] |
包鹏, 王淑莹, 高瑶远, 等. 利用高通量测序技术分析不同溶解氧条件下硝化活性污泥的菌群结构特性[J]. 北京工业大学学报, 2017, 43(5): 801-808. Bao P, Wang S Y, Gao Y Y, et al. Effect of dissolved oxygen on microbial community of nitrify activated sludge based on high-throughput sequencing technology[J]. Journal of Beijing University of Technology, 2017, 43(5): 801-808. |
| [18] |
王腾旭, 马星宇, 王萌萌, 等. 中高温污泥厌氧消化系统中微生物群落比较[J]. 微生物学通报, 2016, 43(1): 26-35. Wang T X, Ma X Y, Wang M M, et al. A comparative study of microbial community compositions in thermophilic and mesophilic sludge anaerobic digestion systems[J]. Microbiology China, 2016, 43(1): 26-35. |
| [19] |
闫媛, 黎力, 王亚宜, 等. 采用高通量测序分析全程自养脱氮(CANON)系统不同脱氮效能下的微生物群落结构[J]. 北京工业大学学报, 2015, 41(10): 1485-1492. Yan Y, Li L, Wang Y Y, et al. Microbial community characteristics of a completely autotrophic nitrogen removal over nitrite (CANON) system based on high-throughput sequencing technology[J]. Journal of Beijing University of Technology, 2015, 41(10): 1485-1492. |
| [20] | Fernandes H, Jungles M K, Hoffmann H, et al. Full-scale sequencing batch reactor (SBR) for domestic wastewater:Performance and diversity of microbial communities[J]. Bioresource Technology, 2013, 132: 262-268. |
| [21] |
张冰, 吴林蔚, 文湘华. 全国城市污水处理厂中微生物群落的溯源分析[J]. 环境科学, 2019, 40(8): 3699-3705. Zhang B, Wu L W, Wen X H. Potential source environments for microbial communities in wastewater treatment plants (WWTPs) in China[J]. Environmental Science, 2019, 40(8): 3699-3705. |
| [22] |
高晨晨, 游佳, 陈轶, 等. 丝状菌污泥膨胀对脱氮除磷功能菌群的影响[J]. 环境科学, 2018, 39(6): 2794-2801. Gao C C, You J, Chen Y, et al. Effect of denitrification and phosphorus removal microorganisms in activated sludge bulking caused by filamentous bacteria[J]. Environmental Science, 2018, 39(6): 2794-2801. |
| [23] |
姚倩, 彭党聪, 赵俏迪, 等. 活性污泥中硝化螺菌(Nitrospira)的富集及其动力学参数[J]. 环境科学, 2017, 38(12): 5201-5207. Yao Q, Peng D C, Zhao Q D, et al. Enrichment of nitrospira in activated sludge and kinetic characterization[J]. Environmental Science, 2017, 38(12): 5201-5207. |
| [24] |
王越, 李秋芬, 张艳. 嗜碱盐单胞菌X3中异化型硝酸盐还原酶编码基因簇的功能验证及蛋白结构预测[J]. 中国海洋大学学报(自然科学版), 2019, 49(4): 33-40. Wang Y, Li Q F, Zhang Y. Function verification and protein structure prediction of gene cluster narGYJV encoding dissimilatory nitrate reductase in Halomonas alkaliphila X3[J]. Periodical of Ocean University of China, 2019, 49(4): 33-40. |
| [25] | Wang T, Li J, Zhang L H, et al. Simultaneous heterotrophic nitrification and aerobic denitrification at high concentrations of NaCl and ammonia nitrogen by Halomonas bacteria[J]. Water Science and Technology, 2017, 76(2): 386-395. |
| [26] | Niu H J Y, Leung D Y, Wong C, et al. Nitric oxide removal by wastewater bacteria in a biotrickling filter[J]. Journal of Environmental Sciences, 2014, 26(3): 555-565. |