 2020, Vol. 41
2020, Vol. 41



2. 西安建筑科技大学西北水资源与环境生态教育部重点实验室, 西安 710055;
3. 西安建筑科技大学陕西省环境工程重点实验室, 西安 710055
2. Key Laboratory of Northwest Water Resource, Environment and Ecology, Ministry of Education, Xi'an University of Architecture and Technology, Xi'an 710055, China;
3. Shaanxi Key Laboratory of Environmental Engineering, Xi'an University of Architecture and Technology, Xi'an 710055, China
由于生物膜附着生长且微生物停留时间较长的特性, 生物膜工艺相对于活性污泥工艺对低温具有更强的耐受性[1, 2].一般认为, 传统活性污泥法在低于5~8℃时硝化作用就基本停止[3~7], 而Young等[7]首次发现MBBR系统即使在1℃下仍能保持一定的硝化活性, 其硝化速率约为20℃时的22.8%.近年来, 典型的生物膜工艺, 如移动床生物膜反应器(MBBR)和生物膜-活性污泥复合系统(IFAS), 由于污泥产量低、比表面积大和管理方便等优点, 被广泛应用于低温地区污水处理以强化硝化作用.其中由于IFAS工艺的改造成本较低, 仅需在原工艺中投加填料, 无需增加池容, 因此被越来越多地应用于现有污水处理厂的升级改造.
于亮[8]采用中试IFAS系统在9~12℃下对城市污水进行处理, 最终出水氨氮浓度仅为0.5~3 mg·L-1, 去除率高达93%, 远高于原工艺平均去除率(66%), 成功强化了硝化性能.裘湛[9]在11℃下运行IFAS工艺发现, 悬浮填料上的硝化活性是活性污泥的1.2倍, 硝化作用主要发生在生物膜上.而汪君晖等[10]研究了城市污水处理厂中生物膜的强化效果发现, 生物膜的硝化贡献率仅为5.5%, 而活性污泥为94.5%, 同样, Lo等[11]的研究发现悬浮污泥的硝化速率为生物膜的1.8倍, 填料区的硝化作用十分有限.因此, 有关IFAS中生物膜的强化效果仍存在争议.同时, 目前针对生物膜工艺的研究多集中于填料投配率、进水负荷等运行参数对硝化性能的影响[12], 而有关生物膜特性及群落结构等微观层面的研究相对较少, 且缺乏有关生物膜系统对低温适应性的深入研究, 无法有效地指导工程设计及实践.
为此, 本研究通过在常温(20℃)下启动MBBR及IFAS系统并逐渐降低系统温度至10℃, 对比分析3个温度条件下两种生物膜系统的硝化性能及生物膜特性, 并在此基础上利用qPCR技术对降温过程中的硝化菌种群结构进行定量分析, 从而探究生物膜系统对低温的适应特性, 以期为其在污水处理行业的应用提供理论依据.
1 材料与方法 1.1 试验装置及运行本试验所用的MBBR和IFAS反应器有效容积分别为4 L和3 L, 通过恒温水浴系统控制反应器内温度.为了快速启动硝化生物膜反应器, 试验所用的接种填料与接种污泥分别取自西安市某污水处理厂的缺氧池和好氧池, 填料比表面积为500 m2·m-3, 投配率设为30%;通过曝气使填料充分搅拌并防止反应器中溶解氧受限.
其中, MBBR反应器的运行周期为8 h(包括进水7 min, 曝气460 min, 出水7 min, 闲置6 min), 换水比为100%, 由于填料占有一部分体积, 每周期出水量为3 L; IFAS反应器的运行周期为6 h(包括进水并曝气6 min, 曝气300 min, 沉淀45 min, 出水6 min, 闲置3 min), 换水比为50%, 污泥停留时间(SRT)控制在15~20 d, 确保两反应器的好氧氮容积负荷均为235 g·(m3·d)-1.根据温度的变化, 反应器运行过程共分为20℃(0~85 d)、15℃(86~152 d)和10℃(153~245 d)这3个阶段.
1.2 试验用水本试验用水采用人工模拟污水, 通过添加(NH4)2SO4使进水氨氮维持在100 mg·L-1, 具体如下:(NH4)2SO4 471.64 mg·L-1, NaHCO3 1 300 mg·L-1, MgSO4·7H2O 325 mg·L-1, CaCl2·2H2O 29.34 mg·L-1, KH2PO4 79.09 mg·L-1, FeSO4·7H2O 4.98 mg·L-1.其中微量元素包括:MnCl2·4H2O 200 μg·L-1, NaMoO4·2H2O 49.62 μg·L-1, CuSO4·5H2O 205.1 μg·L-1, CoCl2·6H2O 2 μg·L-1, ZnSO4·7H2O 59.82 μg·L-1.碳源由C6H12O6 15.2 mg·L-1, NaAc 8.0 mg·L-1, 蛋白胨15.2 mg·L-1组成, BOD5约为40 mg·L-1[13].
1.3 分析项目及方法本试验中常规水质(如NH4+-N、NO2--N和NO3--N)指标根据文献[14]中标准方法进行测定; 溶解氧(DO)和pH分别采用Seven2Go S9型溶氧仪(Mettler Toledo, 瑞士)及雷磁PHS-3C pH计进行测定; 胞外聚合物(EPS)采用阳离子树脂交换法提取, 具体方法见文献[15]; 采用扫描电镜(SEM)观察生物膜形态, 预处理步骤见文献[16].
生物膜厚度:将待切片填料3~5个用包埋剂在4℃下包埋12 h后用冷冻切片机(Leica CM1950, 德国)切片, 切片厚度为20 μm, 每个填料制备3个切片样品, 最终通过尼康50i电子显微镜进行厚度的测定, 每个切片样品分别测定10~20个厚度, 取平均值.
生物膜干重:取3~4个填料烘干后称重记为W1, 将填料在60℃的20% NaOH溶液中水浴加热1 h, 然后用毛刷轻轻除去生物膜后再次烘干称重记为W2, 定义W1和W2的差值与填料数量之比为生物膜干重.
1.4 硝化活性氨氧化活性(AUR)和亚硝酸盐氧化活性(NUR)采用直接取样法进行测定.活性污泥的硝化活性测定前需移除所有填料, 并对活性污泥进行多次离心以去除基质; 生物膜的硝化活性测定前需移除活性污泥.加入初始浓度为30~40 mg·L-1的(NH4)2SO4溶液或NaNO2溶液, 充分曝气并定时取样测定氨氮、亚硝氮及硝氮浓度.根据生化反应速率与温度的关系[式(1)], 将各个温度下的硝化活性统一为20℃下的活性使结果具有可比性:
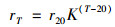
|
(1) |
式中, rT表示温度为T时测得的硝化活性, mg·(L·h)-1; r20表示温度为20℃时的硝化活性, mg·(L·h)-1; T表示温度, ℃; K表示温度校正系数, 对于生物膜系统, K取值为1.098[17], 对于活性污泥系统, K取值为1.04.
1.5 定量PCR(qPCR)采用qPCR技术对生物膜和活性污泥中的氨氧化菌(AOB)、亚硝酸盐氧化菌(NOB)(Nitrospira和Nitrotoga)进行定量分析.取一定数目的填料(视生物膜干重而定)剪碎后加入50 mL去离子水, 滤去填料残渣后于10 000 r·min-1高速离心5 min后弃去上清液, 取适量生物膜样品用土壤基因组DNA提取试剂盒(天根生化科技有限公司)提取DNA, 所提取的基因组DNA纯度及浓度经NANO DROP 2000分光光度计(Thermo Scientific, 美国)测定后置于-20℃下保存; 活性污泥的DNA提取过程与生物膜相似, 直接将适量活性污泥进行离心后提取即可. 50 μL的qPCR混合反应物由1 μL的正向引物, 1 μL的反向引物, 1 μL的模板DNA及25 μL的2×Taq MasterMix(北京康为世纪生物科技有限公司)组成.PCR扩增的条件为:95℃预变性5 min; 在94℃变性30 s, 55℃退火30 s, 72℃延伸30 s, 共30个循环, 最后一轮循环完成后再72℃延伸10 min.每个样品重复3次.
在ABI 7500型qPCR仪(Applied Biosystems, 美国)进行qPCR分析.其中定量PCR的扩增条件为95℃ 30 s后进行40个PCR循环:95℃ 5 s, 60℃ 40 s(收集荧光).试验过程中所涉及的目的基因及其序列见表 1.
|
|
表 1 试验中设计的目的基因及其引物序列 Table 1 Quantitative PCR (qPCR) primers used in this study |
2 结果与讨论 2.1 反应器的启动及运行
MBBR和IFAS反应器进、出水中氮浓度历时变化如图 1所示.比较MBBR与IFAS出水可以看出, 在反应器启动阶段, MBBR和IFAS的出水氨氮及亚硝氮浓度分别在第10 d和第5 d即降解完全, 并达到稳定状态, 这说明IFAS与MBBR相比, 有更好的环境适应能力; 当温度由20℃降低至15℃时(86~152 d), IFAS与MBBR仍维持稳定的出水水质, 受温度影响较小; 当温度降至10℃时(153~245 d), IFAS系统氨氮去除率仍高达(99.79±0.31)%, 出水亚硝氮低于检测下限, 但MBBR的氨氮去除率降至(90.88±5.73)%, 平均出水氨氮浓度为(9.22±5.71)mg·L-1.这说明IFAS比MBBR具有更好的低温适应性.

|
图 1 MBBR及IFAS反应器随温度降低的氮浓度历时变化 Fig. 1 Variation of influent/effluent nitrogen concentration with decreasing temperature in the MBBR and IFAS system |
图 2为MBBR与IFAS在3个温度下的硝化活性对比.如图 2(a)所示, 随着温度从20℃降低至10℃, MBBR中的AUR逐渐从10.91 mg·(L·h)-1降至6.02 mg·(L·h)-1; IFAS中AUR由(12.76±1.01)mg·(L·h)-1降低至6.70 mg·(L·h)-1, 其中悬浮相的AUR从8.84 mg·(L·h)-1降低至4.03 mg·(L·h)-1, 固着相由(3.92±1.01)mg·(L·h)-1降低至2.67 mg·(L·h)-1; 由图 3可知, IFAS中悬浮相的AUR贡献率在3个温度下均高于固着相, 但随着温度的降低, 固着相的贡献率逐渐增加, 由20℃的30.72%增至10℃的39.85%, 谭杰等[12]也得到了相似的结果.而对于NUR, 如图 2(b)所示, MBBR中的NUR逐渐从20℃的35.84 mg·(L·h)-1降至10℃的20.20 mg·(L·h)-1; IFAS中NUR由31.23 mg·(L·h)-1降低至18.23 mg·(L·h)-1, 其中悬浮相的NUR从17.28 mg·(L·h)-1降低至10.21 mg·(L·h)-1, 固着相由13.95 mg·(L·h)-1降低至8.02 mg·(L·h)-1; 同样地, IFAS中悬浮相的NUR贡献率均处优势地位, 但随着温度降低, 固着相的NUR贡献率无明显变化(图 3).由于硝化菌生长速率相对较慢且对温度较为敏感[21], 低温会使硝化活性显著降低, 但填料的投加会使一部分生长相对缓慢的微生物附着在固着相上生长且不会随出水流失[22], 理论上生物膜拥有无限长的SRT, 最终填料上会保持充足的生物量以应对温度的降低.此外, 考虑到生物膜的分层结构能够为不同生态特征的微生物提供生存场所[22, 23], 而通常情况下AOB对DO的亲和力相对于NOB较高[24], 导致AOB更适宜生长在生物膜外侧[25].因此, 相对于NOB, 较好的传质条件会促使生物膜中的AUR贡献率随温度降低呈上升趋势, 从而保证IFAS在低温下仍能保持较高的硝化活性.

|
图 2 MBBR和IFAS反应器的氨氧化活性(AUR)及亚硝酸盐氧化活性(NUR)变化 Fig. 2 AUR and NUR in the MBBR and IFAS system at various temperatures |

|
图 3 IFAS反应器中生物膜与活性污泥的硝化活性及硝化贡献率 Fig. 3 Nitrification activity and contribution rate of biofilm and activated sludge in the IFAS system |
综上可知, 在试验条件下(10~20℃), 活性污泥的硝化贡献率始终高于生物膜, 而随着温度降低, IFAS中生物膜的硝化贡献率呈上升趋势, 起到了强化硝化的效果, 因此在季节性温差较大的北方地区, 采用IFAS工艺可以取得稳定的脱氮效率.
2.3 生物膜特性图 4所示为不同温度下MBBR和IFAS中填料表面的生物膜扫描电镜.从中可以看出, 悬浮填料表面较为粗糙, 适宜微生物附着生长, 填料表面的黏性物质将微生物紧紧黏附在一起, 形成了较为密实的生物膜结构.其中, 随着温度降低, MBBR系统的生物膜明显变厚[图 4(a)~4(c)]; 当温度降至10℃[图 4(c)]时, 部分填料的隔间中甚至发生堵塞现象, 无法形成完整的孔隙空间.Herrling等[26]的研究发现, 生物膜的厚度和隔间孔隙空间会影响载体的流动和传质, 从而导致载体的有效比表面积减小, 最终影响反应器硝化性能[13], 这也一定程度上影响了MBBR在10℃时的出水水质(图 1).而对于IFAS反应器[图 4(d)~4(f)], 不同温度下生物膜的厚度均小于MBBR, 且20℃下填料上附着的生物膜极少[图 4(d)], 这可能是由于常温下悬浮相更有利于传质, 从而影响了硝化菌在固着相上的附着.而随着温度降低, 生物量逐渐增加, 生物膜也变得更加致密.当温度降至10℃时[图 4(f)], 生物膜明显变厚但并未出现堵塞, 这是由于IFAS中同时存在两种生物相进行硝化作用, 且活性污泥的硝化贡献率始终占主导地位(图 3), 因此相同的好氧氮容积负荷下, IFAS生物膜能够利用的基质相对少于MBBR, 进而不易发生填料堵塞, 氨氮能被完全去除(图 1).以上结果表明, IFAS比MBBR具有更好的低温适应性.

|
(a)~(c)分别为MBBR中生物膜在20、15和10℃下的SEM图像; (d)~(f)分别为IFAS中生物膜在20、15和10℃下的SEM图像 图 4 不同温度下MBBR和IFAS生物膜的SEM图像 Fig. 4 SEM image of biofilm at various temperatures in the MBBR and IFAS system |
图 5为MBBR和IFAS中生物膜的EPS及各组分(包括多糖、蛋白质和DNA)随温度降低的变化规律.其中, 由于20℃时IFAS中生物膜厚度过低[图 4(d)], 无法进行EPS的提取, 造成20℃生物膜EPS数据缺失.由图 5可知, 随着温度降低, 生物膜中EPS含量均呈明显上升趋势, 这表明温度的降低会促使微生物分泌更多的EPS.通常认为, EPS是微生物分泌于体外而形成的一种高分子聚合物, 大多覆盖在微生物聚集体表面或者填充在细胞间.而温度的降低会促使微生物体分泌更多的EPS, 使功能菌在各个生物相中相互聚集以应对外界的不利影响[27], 从而形成较厚较重的生物膜.Zhang等[6, 28]的研究同样发现当温度由16.1℃升至28.3℃时, 生物膜浓度由于EPS的减少从348.7 mg·L-1降至188.9 mg·L-1, 硝化生物膜生长速率低于分离速率, 生物膜出现脱落, 反之, EPS的增加会使生物膜生长速率变高.

|
图 5 MBBR和IFAS中生物膜的EPS及组分 Fig. 5 EPS component of biofilm in the MBBR and IFAS system |
图 6所示为MBBR和IFAS反应器在不同温度下的生物膜厚度及干重变化情况.可以看出, MBBR中生物膜厚度从(114.73±30.36)μm(20℃)升高至(782.36±273.80)μm(10℃); IFAS中生物膜厚度从(63.73±17.72)μm(20℃)升高至(565.58±198.49)μm(10℃).相对于接种填料, MBBR中生物膜干重在20、15和10℃下分别增加了7.47%、10.50%和99.86%, 而IFAS中除了20℃时生物膜干重明显下降外, 整体上仍呈上升趋势(20℃:-77.07%; 15℃:11.56%; 10℃:18.65%).在中温环境下(20℃), 生物膜能够利用到的基质相对少于活性污泥, 再加上充分曝气带来较强的水力冲刷作用, 最终导致20℃下生物膜厚度及干重明显较低.有研究[29]同样发现污水处理厂中常温环境下生物膜对IFAS工艺的硝化贡献率仅为1.3%.因此, 低温环境下EPS含量的增加会导致生物膜中生物量出现增长, 从而一定程度上缓解低温环境下硝化活性的下降, 而在中/常温下生物膜对硝化的强化作用并不显著.

|
图 6 不同温度下的生物膜干重及厚度 Fig. 6 Biofilm mass and biofilm thickness of the MBBR and IFAS system at various temperatures |
此外, 本研究中硝化MBBR系统的总氮去除率约为22.98%, 大于IFAS系统的12.16%(图 1).这是由于IFAS相对于MBBR同时存在生物膜相和活性污泥相共同利用基质, 因此在相同的好氧容积负荷下, MBBR中的生物膜更厚(图 6).有研究通过传质模型证明DO在絮体中的扩散会限制在100 μm以内[30], 因此较厚的生物膜一定程度上会阻碍DO向生物膜内层的传质, 最终导致生物膜内部形成厌氧/缺氧层进行反硝化作用, 产生氮气, 使系统出现氮损失[31].由此可见, MBBR相对于IFAS有更好的总氮去除效果.
2.4 硝化菌群落结构考虑到已有大量文献证明Nitrotoga作为一种耐冷型NOB(4~28℃)广泛存在于污水处理厂中[20, 32], 为了探究硝化生物膜系统对低温的适应特征, 本研究将Nitrotoga以及目前污水处理厂中最为常见的NOB(Nitrospira)作为NOB定量检测对象.
图 7(a)为MBBR生物膜中AOB和NOB丰度的变化趋势.从中可以看出, 随着温度降低, AOB和NOB拷贝数并未下降.其中, AOB丰度分别为9.37×108、1.18×109和2.29×109 copies·L-1, 呈上升趋势.而对于NOB, 相对于Nitrotoga, Nitrospira在MBBR系统中始终占优势, 随着温度降低, Nitrospira拷贝数从20℃的3.73×108 copies·L-1增加至10℃的1.12×109 copies·L-1.而Nitrotoga丰度维持在(1.39±0.69)×108 copies·L-1, 有小幅增加但并不明显.

|
图 7 MBBR及IFAS中生物膜和活性污泥随温度降低的硝化菌丰度变化 Fig. 7 Cell abundance of AOB and NOB (Nitrospira and Nitrotoga) with decreasing temperature in the MBBR and IFAS system |
而对于IFAS系统, 由图 7(b)可见, Nitrospira同样始终为优势NOB菌属.当温度由15℃降至10℃, 除了Nitrotoga出现小幅减少外, 生物膜中的AOB和Nitrospira拷贝数均呈增加趋势, 其中Nitrospira拷贝数从3.18×108copies·L-1明显增加至8.76×108copies·L-1.而活性污泥中的AOB拷贝数从20℃时的5.23×108 copies·g-1逐渐降低至10℃时的3.64×108 copies·g-1, NOB丰度整体上也呈下降趋势, 且Nitrospira在15~20℃时下降不明显[图 7(c)].值得一提的是, Nitrotoga作为一种嗜冷型NOB, 其丰度随温度变化在本研究中并未显示出明显的相关性, 且温度降低过程中Nitrotoga并未成为优势NOB, 因此尽管可以在生物膜中检测出Nitrotoga, 但其对低温环境的适应性并不是导致生物膜对低温适应性的主要原因, 目前也并未发现有关生物膜系统中Nitrotoga占优势的报道.
通过对比MBBR和IFAS中生物膜的硝化群落结构发现, 随着温度降低, 生物膜相中的AOB和Nitrospira拷贝数呈上升趋势, 而IFAS活性污泥相中的硝化菌拷贝数均出现不同程度的减少, 这表明低温会使生物膜中的活性细胞数增加, 而活性污泥则恰好相反.Young等[7]通过对20℃及1℃下硝化MBBR中的生物膜进行LIVE/DEAD细胞活性检测同样发现, 1℃下活性硝化细胞占比高于20℃.因此, 尽管细胞活性会随着温度降低而降低[13], 但具有硝化活性的细胞数目及占比呈增加趋势, 再加上低温环境下细胞衰亡率会显著降低[33], 生物膜厚度及干重明显增加(图 6), 最终一定程度上能够弥补生物膜中由于温度降低而导致的硝化活性衰减, 从而体现出生物膜系统对低温的适应性.
3 结论(1) 生物膜系统对低温具有一定适应性.低温下生物膜中硝化细胞拷贝数会小幅上升, 同时伴随着生物量的增加, 从而一定程度上弥补低温下硝化活性的衰减.
(2) IFAS比MBBR具有更好的低温适应性.温度降低会导致生物膜系统中EPS含量明显上升, 进而引起生物膜厚度增加, 相同负荷下的MBBR在低温下更容易发生填料堵塞, 导致水质恶化.
(3) 在10~20℃下, IFAS中活性污泥的硝化贡献率始终占主导地位, 而随着温度降低, 生物膜的硝化贡献率呈上升趋势, 起到了强化硝化的作用.因此, 在季节性温差较大的北方地区, 采用IFAS工艺可以缓解因冬季水温较低而导致的硝化不充分.
| [1] | Young B, Delatolla R, Ren B S, et al. Pilot-scale tertiary MBBR nitrification at 1℃:characterization of ammonia removal rate, solids settleability and biofilm characteristics[J]. Environmental Technology, 2016, 37(16): 2124-2132. DOI:10.1080/09593330.2016.1143037 |
| [2] | Hoang V, Delatolla R, Abujamel T, et al. Nitrifying moving bed biofilm reactor (MBBR) biofilm and biomass response to long term exposure to 1℃[J]. Water Research, 2014, 49: 215-224. DOI:10.1016/j.watres.2013.11.018 |
| [3] | Delatolla R, Young B, Stintzi A. Application of the MBBR technology to achieve nitrification below 1℃: biofilm and microbiome analysis[A]. In: Mannina G (Ed.). Frontiers in Wastewater Treatment and Modelling[M]. Cham: Springer, 2017. 513-517. |
| [4] | Hurse T J, Connor M A. Nitrogen removal from wastewater treatment lagoons[J]. Water Science and Technology, 1999, 39(6): 191-198. DOI:10.2166/wst.1999.0296 |
| [5] | Choi E, Rhu D, Yun Z, et al. Temperature effects on biological nutrient removal system with weak municipal wastewater[J]. Water Science and Technology, 1998, 37(9): 219-226. DOI:10.2166/wst.1998.0360 |
| [6] | Zhang S F, Wang Y Y, He W T, et al. Impacts of temperature and nitrifying community on nitrification kinetics in a moving-bed biofilm reactor treating polluted raw water[J]. Chemical Engineering Journal, 2014, 236: 242-250. DOI:10.1016/j.cej.2013.09.086 |
| [7] | Young B, Delatolla R, Kennedy K, et al. Low temperature MBBR nitrification:microbiome analysis[J]. Water Research, 2017, 111: 224-233. DOI:10.1016/j.watres.2016.12.050 |
| [8] |
于亮. 低温条件生物膜与活性污泥复合工艺强化生物脱氮性能的中试研究[J]. 环境科学与管理, 2017, 42(4): 96-99. Yu L. Pilot-scale study on enhancing nitrogen removal efficiency under low temperature by biofilm and activated sludge hybrid process[J]. Environmental Science and Management, 2017, 42(4): 96-99. DOI:10.3969/j.issn.1673-1212.2017.04.022 |
| [9] |
裘湛. 生物膜-活性污泥复合系统的强化硝化[J]. 上海电力学院学报, 2016, 32(4): 385-388. Qiu Z. Nitrification enhancement by biofilm-coupled activated sludge system[J]. Journal of Shanghai University of Electric Power, 2016, 32(4): 385-388. DOI:10.3969/j.issn.1006-4729.2016.04.016 |
| [10] |
汪君晖, 孙红芳, 蔡虎林, 等. 生物膜对A2/O系统硝化过程的强化效果分析[J]. 水处理技术, 2016, 42(2): 113-117. Wang J H, Sun H F, Cai H L, et al. The analysis of nitrification effect enhanced by biofilm in A2/O process[J]. Technology of Water Treatment, 2016, 42(2): 113-117. |
| [11] | Lo I W, Lo K V, Mavinic D S, et al. Contributions of biofilm and suspended sludge to nitrogen transformation and nitrous oxide emission in hybrid sequencing batch system[J]. Journal of Environmental Sciences, 2010, 22(7): 953-960. DOI:10.1016/S1001-0742(09)60204-7 |
| [12] |
谭杰, 董滨, 戴晓虎. 温度对生物膜-活性污泥复合工艺硝化特性及硝化菌种群的影响[J]. 净水技术, 2016, 35(2): 21-25. Tan J, Dong B, Dai X H. Influence of temperature on nitrification characteristics and nitrifying bacteria community in integrated fixed film-activated sludge process[J]. Water Purification Technology, 2016, 35(2): 21-25. DOI:10.3969/j.issn.1009-0177.2016.02.005 |
| [13] | Young B, Banihashemi B, Forrest D, et al. Meso and micro-scale response of post carbon removal nitrifying MBBR biofilm across carrier type and loading[J]. Water Research, 2016, 91: 235-243. DOI:10.1016/j.watres.2016.01.006 |
| [14] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版).北京: 中国环境科学出版社, 2002: 105-271. |
| [15] | Wang B B, Zhang L, Peng D C, et al. Extended filaments of bulking sludge sink in the floc layer with particulate substrate[J]. Chemosphere, 2013, 93(11): 2725-2731. DOI:10.1016/j.chemosphere.2013.08.099 |
| [16] |
赵远哲, 杨永哲, 王海燕, 等. 新型填料A/O生物滤池处理低碳氮比农村污水脱氮[J]. 环境科学, 2020. Zhao Y Z, Yang Y Z, Wang H Y, et al. Nitrogen removal in low C/N rural sewage treatment by A/O biofilter packing with new types fillers[J]. Environmental Science, 2020. DOI:10.13227/j.hjkx.201910118 |
| [17] | Salvetti R, Azzellino A, Canziani R, et al. Effects of temperature on tertiary nitrification in moving-bed biofilm reactors[J]. Water Research, 2006, 40(15): 2981-2993. DOI:10.1016/j.watres.2006.05.013 |
| [18] | Rotthauwe J H, Witzel K P, Liesack W. The ammonia monooxygenase structural gene amoA as a functional marker:molecular fine-scale analysis of natural ammonia-oxidizing populations[J]. Applied and Environmental Microbiology, 1997, 63(12): 4704-4712. DOI:10.1128/AEM.63.12.4704-4712.1997 |
| [19] | Pester M, Maixner F, Berry D, et al. NxrB encoding the beta subunit of nitrite oxidoreductase as functional and phylogenetic marker for nitrite-oxidizing Nitrospira[J]. Environmental Microbiology, 2014, 16(10): 3055-3071. DOI:10.1111/1462-2920.12300 |
| [20] | Lücker S, Schwarz J, Gruber-Dorninger C, et al. Nitrotoga-like bacteria are previously unrecognized key nitrite oxidizers in full-scale wastewater treatment plants[J]. The ISME Journal, 2015, 9(3): 708-720. DOI:10.1038/ismej.2014.158 |
| [21] | Guo J H, Peng Y Z, Huang H J, et al. Short- and long-term effects of temperature on partial nitrification in a sequencing batch reactor treating domestic wastewater[J]. Journal of Hazardous Materials, 2010, 179(1-3): 471-479. DOI:10.1016/j.jhazmat.2010.03.027 |
| [22] |
杨岚, 彭永臻, 李健伟, 等. 缺氧MBBR耦合部分厌氧氨氧化强化城市生活污水深度脱氮[J]. 环境科学, 2019, 40(8): 3668-3674. Yang L, Peng Y Z, Li J W, et al. Advanced denitrification of municipal wastewater achieved via partial ANAMMOX in anoxic MBBR[J]. Environmental Science, 2019, 40(8): 3668-3674. |
| [23] | Soliman M, Eldyasti A. Ammonia-oxidizing bacteria (AOB):opportunities and applications-a review[J]. Reviews in Environmental Science and Bio/Technology, 2018, 17(2): 285-321. DOI:10.1007/s11157-018-9463-4 |
| [24] |
杨庆, 杨玉兵, 刘秀红, 等. 不同溶解氧环境下氨氧化菌的氧半饱和常数比较[J]. 中国给水排水, 2017, 33(23): 22-26. Yang Q, Yang Y B, Liu X H, et al. Oxygen half-saturation constants of ammonia oxidizing bacteria (AOB) in nitrification sludge under different dissolved oxygen[J]. China Water & Wastewater, 2017, 33(23): 22-26. |
| [25] | Poot V, Hoekstra M, Geleijnse M A A, et al. Effects of the residual ammonium concentration on NOB repression during partial nitritation with granular sludge[J]. Water Research, 2016, 106: 518-530. DOI:10.1016/j.watres.2016.10.028 |
| [26] | Herrling M P, Guthausen G, Wagner M, et al. Determining the flow regime in a biofilm carrier by means of magnetic resonance imaging[J]. Biotechnology and Bioengineering, 2015, 112(5): 1023-1032. DOI:10.1002/bit.25510 |
| [27] | Sheng G P, Yu H Q, Li X Y. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems:a review[J]. Biotechnology Advances, 2010, 28(6): 882-894. DOI:10.1016/j.biotechadv.2010.08.001 |
| [28] | Zhang S F, Wang Y Y, He W T, et al. Responses of biofilm characteristics to variations in temperature and NH4+-N loading in a moving-bed biofilm reactor treating micro-polluted raw water[J]. Bioresource Technology, 2013, 131: 365-373. DOI:10.1016/j.biortech.2012.12.172 |
| [29] |
王玉.悬浮填料对城市污水A2/O工艺硝化过程强化研究[D].西安: 西安建筑科技大学, 2016. 28-30. Wang Y. Effect of suspended carrier on the enhancement of nitrification in A2/O process for urban sewage treatment[D]. Xi'an: Xi'an University of Architecture and Technology, 2016. 28-30. |
| [30] | Picioreanu C, Xavier J B, Van Loosdrecht M C M. Advances in mathematical modeling of biofilm structure[J]. Biofilms, 2004, 1(4): 337-349. DOI:10.1017/S1479050505001572 |
| [31] | Winkler M K H, Bassin J P, Kleerebezem R, et al. Unravelling the reasons for disproportion in the ratio of AOB and NOB in aerobic granular sludge[J]. Applied Microbiology and Biotechnology, 2012, 94(6): 1657-1666. DOI:10.1007/s00253-012-4126-9 |
| [32] | Kitzinger K, Koch H, Lücker S, et al. Characterization of the first "Candidatus Nitrotoga" isolate reveals metabolic versatility and separate evolution of widespread nitrite-oxidizing bacteria[J]. mBio, 2018, 9(4): e01186-18. |
| [33] |
刘占孟, 徐宇峰, 李思敏. 温度对活性污泥产率的影响研究[J]. 中国给水排水, 2015, 31(19): 84-88. Liu Z M, Xu Y F, Li S M. Effect of temperature on activated sludge yield[J]. China Water & Wastewater, 2015, 31(19): 84-88. |