 2020, Vol. 41
2020, Vol. 41



城市污水处理厂目前面临着进水碳氮比较低的情况, 因此解决碳源利用的问题以及探索低碳源情况下高效脱氮的方法受到了广泛关注[1, 2].许多研究表明, 溶解氧对活性污泥系统在低碳源条件下的处理效果有很大影响[3, 4], 氧浓度和氧传质直接影响着微生物的脱氮性能[5, 6].活性污泥系统的溶解氧与采用的供氧模式关系密切[7, 8], 目前常见的供氧模式为连续曝气(A2O[9]、SBR[10]等)和间歇曝气(氧化沟[11]等)两种, 采用不同供氧模式的污水处理工艺对低碳源污水的处理性能也有很大差异[12].近年来, 由于在一般进水情况下间歇曝气的供氧模式能节省能源[13]、改善出水水质[14]且有利于短程硝化的实现[15, 16]而引起了广泛关注, 但是对连续曝气和间歇曝气两种活性污泥系统在处理低C/N污水时的脱氮能力的比较研究较少, 间歇曝气系统能否在应对低碳源污水时仍有很好的效果鲜见报道.
为研究低碳源条件下供氧模式对活性污泥系统脱氮性能的影响, 本文分别建立了连续曝气和间歇曝气的两种SBR活性污泥系统, 研究了不同供氧模式对低碳源条件下活性污泥系统处理性能以及碳源利用途径的影响, 并对两种供氧模式下活性污泥系统中脱氮功能微生物的群落结构进行分析, 最后基于16S rRNA测序功能预测对不同供氧模式下微生物代谢机制的差异进行分析, 以期为探究低碳源条件下提高脱氮性能的新技术提供理论支持.
1 材料与方法 1.1 实验用水与接种污泥本实验用水采用人工配水, 碳源由无水乙酸钠提供, 氮源由氯化铵提供, 磷元素由磷酸二氢钾提供, 用碳酸氢钠调节pH在7.4~8.1, 实验各阶段进水指标如表 1所示.接种污泥取自西安市第五污水处理厂生物池的回流污泥, 回流污泥经过48 h闷曝后静沉并去掉杂质, 再将污泥加入反应器开始实验.
|
|
表 1 实验各阶段进水指标 Table 1 Influent indicators of each stage of the experiment |
1.2 实验装置
本实验所用SBR反应器如图 1所示, 反应器由进水泵、空气泵、曝气头、空气流量计、出水泵、搅拌器以及有机玻璃主体组成.

|
图 1 实验装置示意 Fig. 1 Experimental device |
连续供氧系统采用缺氧、好氧和缺氧方式的连续曝气系统, 类似于A2O工艺, 控制好氧段DO为1.5 mg ·L-1, 运行周期为6 h, 每天运行4个周期, 整个操作程序采用自动化控制.运行方式为:进水(10 min)、缺氧搅拌(60 min)、曝气搅拌(110 min)、缺氧搅拌(120 min)、沉淀(30 min)、排水(30 min).
间歇供氧系统采用好氧/缺氧的多循环间歇曝气系统, 类似于氧化沟工艺, 控制好氧段DO为1.5 mg ·L-1, 运行周期为6 h, 每天运行4个周期, 每一运行周期先经过10个小循环, 每个小循环时间为30 min, 包括进水(2 min)、曝气搅拌(10 min)、缺氧搅拌(18 min).在所有小循环结束时, 反应器经过30 min的沉淀后, 再通过30 min来排水.
连续曝气系统一个周期中总曝气时间为110 min, 间歇曝气系统一个周期中总曝气时间为100 min, 单位时间曝气量均为32 L ·h-1.
1.3 分析方法 1.3.1 常规分析项目与方法|
|
表 2 常规指标检测方法 Table 2 General indicator detection methods |
1.3.2 高通量测序方法
(1) DNA提取
采用OMEGA E.Z.N.ATM Mag-Bind Soil DNA Kit试剂盒提取污泥DNA.
(2) PCR反应与高通量测序
① 第一轮扩增用Qubit 3.0 DNA检测试剂盒精确定量DNA量.PCR所用引物为MiSeq测序平台的V3-V4通用引物;②第二轮扩增引入Illumina桥式PCR兼容引物;③ DNA纯化回收对细菌和古菌扩增的PCR产物和正常扩增片段在400 bp以上的PCR产物, 用0.6倍的磁珠处理.对于真菌PCR产物和其他扩增片段小于400 bp的PCR产物, 用0.8倍的磁珠处理;④定量混合利用Qubit3.0 DNA检测试剂盒对回收的DNA精确定量.等量混合时, 每个样品DNA量取10 ng, 最终上机测序浓度为20 pmol.
2 结果与讨论 2.1 供氧模式对低碳源污水处理效果的影响低碳源期连续曝气系统[图 2(a)]和间歇曝气系统[图 2(b)]出水中氨氮的浓度分别为3.27 mg ·L-1和4.57 mg ·L-1, 氨氮去除率分别为92.45%和90.66%, 硝氮浓度分别为11.18 mg ·L-1和13.59 mg ·L-1, 总氮浓度分别为13.51 mg ·L-1和20.36 mg ·L-1, 总氮去除率分别为75.21%和57.31%.调整期连续供氧系统随着C/N由5逐渐降低至3, 出水中氨氮的去除率降低了4.01%, 总氮去除率降低了14.29%.而间歇供氧系统碳源逐渐降低时, 出水中氨氮去除率降低了5.23%, 总氮去除率降低了27.74%.

|
图 2 两个反应器出水氮去除效果 Fig. 2 Nitrogen removal effect of two reactor effluents |
反应器出水COD浓度如图 3所示, 连续曝气系统[图 3(a)]在驯化期的COD出水浓度为24.02 mg ·L-1, 去除率为92.01%;低碳源期的出水COD浓度为30.74 mg ·L-1, 去除率为81.51%, 去除效果较为稳定.间歇曝气系统[图 3(b)]出水COD浓度波动较大, 驯化期的COD出水浓度为26.49 mg ·L-1, 去除率为91.17%;低碳源期的出水COD浓度为31.44 mg ·L-1, 去除率为79.35%.

|
图 3 两个反应器出水COD去除效果 Fig. 3 COD removal effect of two reactor effluents |
结果表明, 虽然间歇曝气系统较连续曝气系统可以节省能耗, 但原水碳源浓度降低后连续曝气系统的总氮去除率较间歇曝气系统高17.92%, 连续曝气系统中总氮去除率受低碳源条件影响程度较间歇曝气系统低44.74%.以连续曝气为供氧模式的活性污泥系统在低碳源条件下总氮的去除率和对碳源不足的适应能力均高于间歇曝气系统, 由此可见, 连续供氧条件下活性污泥系统应对低碳源污水的效果更好.
2.2 不同供氧模式下氮的去除及碳源利用途径为了具体研究供氧模式对低碳源情况下活性污泥系统脱氮过程的影响, 分别对连续供氧系统和间歇供氧系统的典型周期内物质变化情况和碳源利用途径进行了分析.
连续曝气系统在驯化期时[图 4(a)], 进水后COD浓度迅速由133.21 mg ·L-1下降至59.14 mg ·L-1, 这主要是由于微生物对有机物的吸附作用.好氧段氨氮浓度由15.82 mg ·L-1迅速降低至6.13 mg ·L-1, 硝酸盐浓度由0.15 mg ·L-1迅速上升至7.22 mg ·L-1, 总氮浓度由16.55 mg ·L-1降低至12.94 mg ·L-1, COD浓度由59.14 mg ·L-1缓慢降低至42.52 mg ·L-1, 说明在好氧段发生了较为明显的SND现象[18].缺氧段氨氮浓度由6.13 mg ·L-1降低至2.82 mg ·L-1, 硝氮浓度由7.22 mg ·L-1降低至5.23 mg ·L-1, 总氮浓度由12.94 mg ·L-1降低至8.15 mg ·L-1, COD浓度由42.52 mg ·L-1降低至25.21 mg ·L-1.而在低碳源期[图 4(b)], 进水后COD浓度从81.13 mg ·L-1降至45.54 mg ·L-1.好氧段氨氮浓度由16.51 mg ·L-1迅速降低至6.32 mg ·L-1, 硝酸盐浓度由0.51 mg ·L-1迅速上升至9.12 mg ·L-1, 但由于碳源已经处在较低水平, SND现象不明显, 总氮浓度由18.52 mg ·L-1降低至15.74 mg ·L-1, COD浓度由45.53 mg ·L-1缓慢降低至36.81 mg ·L-1.缺氧段COD浓度由36.82 mg ·L-1降低至31.61 mg ·L-1, 此时碳源已经严重不足, 反硝化反应大幅受限.氨氮浓度由6.32 mg ·L-1降低至5.54 mg ·L-1, 硝氮浓度仅由9.11 mg ·L-1降低至8.22 mg ·L-1, 总氮浓度仅由15.71 mg ·L-1降低至13.45 mg ·L-1.

|
图 4 连续曝气系统典型周期内物质的变化 Fig. 4 Changes in matter during a typical cycle of a continuous aeration system |
间歇曝气系统在不同时期的典型周期内的物质变化情况如图 5所示.在驯化期时[图 5(a)], 最后一个小循环在曝气段, COD浓度由49.11 mg ·L-1快速降低至30.35 mg ·L-1, 氨氮浓度由5.13 mg ·L-1快速降低至3.02 mg ·L-1, 硝氮浓度由4.54 mg ·L-1上升至5.95 mg ·L-1, 总氮浓度由9.81 mg ·L-1降低至8.93 mg ·L-1.而在缺氧段, COD浓度由30.32 mg ·L-1缓慢降低至24.21 mg ·L-1, 氨氮浓度由3.01 mg ·L-1缓慢降低至2.65 mg ·L-1, 硝氮浓度由5.92 mg ·L-1缓慢降低至5.71 mg ·L-1, 总氮仅由8.94 mg ·L-1降低至8.42 mg ·L-1.在低碳源期时[图 5(b)], 最后一个小循环曝气段的COD浓度由39.12 mg ·L-1降低至28.52 mg ·L-1, 这时碳源已经处于很低的水平, 氨氮浓度由5.15 mg ·L-1降低至4.41 mg ·L-1, 硝氮浓度由9.54 mg ·L-1上升至10.02 mg ·L-1, 总氮浓度由14.72 mg ·L-1降低至14.44 mg ·L-1.缺氧段的COD浓度由28.52 mg ·L-1降低至26.51 mg ·L-1, 氨氮浓度由4.43 mg ·L-1降低至4.14 mg ·L-1, 硝氮浓度由于硝化与反硝化作用几乎没有降低, 总氮浓度仅由14.44 mg ·L-1降低至14.32 mg ·L-1, 反硝化作用几乎完全被抑制.

|
图 5 间歇曝气系统典型周期内物质的变化 Fig. 5 Changes in matter during a typical cycle of an intermittent aeration system |
反应器进入低碳源期后, 连续曝气系统在第一个缺氧段COD去除率由55.62%降低至43.91%, 在好氧段COD去除率由28.14%降低至19.22%, 总氮去除率由21.92%降低至15.25%, 在第二个缺氧段COD去除率由40.71 mg ·L-1降低至14.21%, 总氮去除率由37.22%降低至17.64%.间歇曝气系统每个小循环的曝气段的COD平均去除率由6.08%降低至5.95%, 总氮平均去除率由4.27%降低至4.05%.缺氧段的COD平均去除率由3.04%降低至1.98%, 总氮平均去除率由4.18%降低至1.94%.碳源不足对连续曝气系统和间歇曝气系统的脱氮性能影响主要在反应后期的缺氧段, 连续曝气系统原水碳源浓度降低导致缺氧段总氮去除率下降较少, 同时连续曝气系统在好氧段能发生较明显的SND现象, 所以, 在碳源不足情况下连续曝气系统的脱氮性能优于间歇曝气系统.
通过式(1)~(3)计算碳源利用途径[19, 20], 结果如表 3所示, 由碳源充足到碳源不足的过程中, 连续供氧系统缺氧段反硝化碳源利用率下降了4.65%, 好氧段反硝化碳源利用率下降了2.21%, 总碳源脱氮利用率下降了3.76%.间歇曝气系统缺氧段反硝化碳源利用率下降了6.67%, 好氧段反硝化碳源利用率下降了0.91%, 总碳源脱氮利用率下降了8.34%.间歇曝气系统的缺氧段反硝化碳源利用率和总碳源反硝化利用率在碳源不足的情况下下降率均大于连续曝气系统.
|
|
表 3 碳源利用途径计算 Table 3 Calculation of carbon source utilization route |
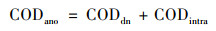
|
(1) |
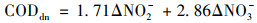
|
(2) |
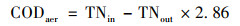
|
(3) |
式中, CODano为缺氧段的COD总消耗量, mg ·L-1; CODdn为反硝化的COD消耗量, mg ·L-1; CODintra为储存内碳源的COD消耗量, mg ·L-1; 1.71和2.86为每反硝化单位NO2--N和NO3--N所消耗的COD量(以COD/N计), mg ·mg-1; CODaer为好氧段反硝化的COD消耗量, mg ·L-1; TNin和TNout为好氧段在初始时和结束时的总氮浓度, mg ·L-1.
将典型周期物质变化情况与碳源利用途径与反应器脱氮性能相结合可以得出, 进水碳源不足抑制了活性污泥系统缺氧段反硝化的进行从而导致系统脱氮性能的下降.而在进水碳源降低时连续曝气系统的缺氧段反硝化碳源利用率较高, 反硝化脱氮所受影响较小, 从而使连续曝气系统的总体脱氮性能受影响较小, 因此, 连续曝气系统在应对低碳源污水时处理效果优于间歇曝气系统.
2.3 供氧模式与微生物群落的关联关系由表 4可知, 所有样品Coverage指数均大于0.995, 说明测序结果能反映微生物样品的真实情况.连续曝气系统中驯化期末和低碳源期末的微生物样品分别用HCA和LCA表示, 间歇曝气系统中驯化期末和低碳源期末的微生物样品分别用HIA和LIA表示.
|
|
表 4 微生物多样性分析 Table 4 Analysis of microbial diversity |
驯化期时间歇曝气系统的微生物群落相对丰度略高于连续曝气系统, 但是在低碳源情况下, 连续曝气系统的微生物群落相对丰度略高于间歇曝气系统.反应器进入低碳源期后, 连续曝气系统的微生物群落相对丰度降低了15.29%, 而间歇曝气系统的微生物群落相对丰度降低了22.73%, 说明低碳源情况下间歇曝气系统的微生物群落相对丰度受到很大影响, 而连续曝气系统受影响较小.图 6为4个活性污泥样品微生物OTU维恩图, 连续曝气系统中, 驯化期和低碳源期活性污泥样本中OTUs数量分别为2 859和2 022个, 在间歇曝气系统系统中, 驯化期和低碳源期活性污泥样本中OTUs数量分别为3 267和2 570个.在进入低碳源期后, 间歇曝气系统的微生物群落结构相比于连续曝气系统有更大的变化, 说明低碳源情况对连续曝气系统的微生物群落结构的影响小于间歇曝气系统.

|
图 6 OTU韦恩图 Fig. 6 OTU Venn diagram |
4个微生物样品门水平群落结构的分布如图 7所示, 相对丰度最高的为Proteobacteria、Bacteroidetes、Acidobacteria和Planctomycetes这4个菌门, 反应器进入低碳源期后, 连续曝气系统中Proteobacteria门的相对丰度由33.01%升高到36.36%, Bacteroidetes门的相对丰度由42.02%升高到50.16%, Acidobacteria门的相对丰度由4.21%降低到2.57%, Planctomycetes门的相对丰度由1.37%升高到1.89%.间歇曝气系统中Proteobacteria门的相对丰度由26.24%降低到13.51%, Bacteroidetes门的相对丰度由31.29%升高到42.7%, Acidobacteria门的相对丰度由13.89%降低到9.11%, Planctomycetes门的相对丰度由6.72%升高到12.25%.由此可知, Proteobacteria、Bacteroidetes和Planctomycetes门在低碳源情况下适应能力较强, 其中Proteobacteria门可能是使低碳源条件下连续曝气系统活性污泥脱氮性能优于间歇曝气系统的重要菌门.

|
图 7 门水平群落结构分布 Fig. 7 Distribution of horizontal community structure |
在属水平上, 如图 8所示, 反应器进入低碳源期后连续曝气系统中Comamonas、Methylocystis、Gemmobacter、Clostridium XI、Acinetobacter和Lysobacter这6个菌属的相对丰度分别从0.13%、0.53%、0.22%、1.08%、0.03%和0.09%上升至1.42%、1.45%、0.69%、2.04%、1.42%和2.97%.而间歇曝气系统中这6个菌属的相对丰度分别从0.35%、0.65%、1.35%、0.5%、0.35%和0.93%降低至0.06%、0.44%、0.04%、0.1%、0.01%和0.01%.连续曝气系统中这些菌属在低碳源情况下的相对丰度有所增加, 而间歇曝气系统在低碳源时这些菌属相对丰度却在降低, 说明在这些菌属的作用下可能使连续曝气系统相比于间歇曝气系统更加适应对低碳源污水的处理.

|
图 8 属水平群落结构分布 Fig. 8 Distribution of horizontal community structure |
不同供氧模式下活性污泥系统中相对丰度较高的脱氮功能菌见表 5.其中属于氨氧化细菌(AOB)的菌属为Nitrosomomas, 属于亚硝酸盐氧化菌(NOB)的菌属为Nitrospira[21, 22].具有反硝化功能的菌属包括:Clostridium sensu stricto、Acinetobacter、Comamonas、Hyphomicrobium、Azonexus和Flavobacterium[23].
|
|
表 5 两种供氧模式活性污泥系统脱氮功能菌相对丰度(属水平) /% Table 5 Relative abundance of denitrifying functional bacteria in activated sludge systems in two oxygen supply modes (genus level)/% |
连续曝气系统在进入低碳源期后, 硝化菌中Nitrosomonas的相对丰度由2.25%降低至0.16%, Nitrospira的相对丰度由0.61%降低至0.14%.反硝化菌总的相对丰度由5.66%降低至5.44%, 其中Clostridium sensu stricto的相对丰度由3.94%降低至2.56%, Flavobacterium的相对丰度由0.10%升高至0.22%, Hyphomicrobium的相对丰度由0.51%降低至0.33%, Comamonas的相对丰度由0.23%升高至0.99%, Acinetobacter的相对丰度由0.22%升高至0.94%, Azonexus的相对丰度由0.66%降低至0.40%.而间歇曝气系统在进水碳源不足后, 硝化菌中Nitrosomonas的相对丰度由0.25%降低至0.13%, Nitrospira的相对丰度由3.54%降低至0.18%.反硝化菌总的相对丰度由3.50%降低至1.98%, 其中Clostridium sensu stricto的相对丰度由0.72%降低至0.15%, Flavobacterium的相对丰度由0.19%升高至0.25%, Hyphomicrobium的相对丰度由0.58%升高至0.29%, Comamonas的相对丰度由0.35%降低至0.06%, Acinetobacter的相对丰度由0.35%降低至0.08%, Azonexus的相对丰度由1.31%升高至1.15%.
在驯化期时, 连续曝气系统中主要的硝化菌为Nitrosomonas, 而间歇曝气系统中主要的硝化菌为Nitrospira, 但在碳源不足时两种系统中这两种硝化菌的相对丰度基本相当.连续曝气系统中Clostridium sensu stricto为主要的反硝化菌, 而间歇曝气系统中主要的反硝化菌为Azonexus.在进入低碳源期后, 连续曝气系统中Comamonas和Acinetobacter的相对丰度上升, 而间歇曝气系统中两者的相对丰度下降.而Clostridium sensu stricto的相对丰度相比于间歇曝气系统降低较少且绝对值较高, 这说明在连续曝气系统中, 低碳源条件更有利于这3种菌的生长.
综上, 连续曝气的供氧模式促使Clostridium sensu stricto、Comamonas和Acinetobacter等功能微生物在低碳源条件下更好地生长, 进而导致连续曝气系统具有较好的低碳源污水处理性能.
2.4 不同供氧模式下微生物代谢机制差异为了从基因水平上解释低碳源情况下供氧模式对活性污泥系统处理性能的影响, 基于16S rRNA测序, 通过KEGG数据库预测功能基因的丰度[24, 25].
根据图 9可知, 在进水碳源不足时, 连续曝气系统中细胞运动功能基因(cell motility)、碳水化合物代谢功能基因(carbohydrate metabolism)、膜运输功能基因(membrane transport)、异生物素生物降解和代谢功能基因(xenobiotics biodegradation and metabolism)和细胞组成功能基因(cellular processes and signaling)在微生物中的相对丰度均高于间歇曝气系统, 尤其是膜运输功能基因.膜运输功能越强说明微生物与环境系统的物质交换越频繁, 碳水化合物代谢功能越强说明功能微生物对碳源的利用越充分, 而细胞组成功能越强则说明微生物的生长代谢越旺盛[26~28].综上, 在低碳源条件下连续曝气的供氧模式加强了微生物的上述功能, 从而使活性污泥系统更适应进水碳源不足的情况.

|
图 9 基于KEGG预测功能基因的相对丰度 Fig. 9 Predicting the relative abundances of functional genes based on KEGG |
(1) 连续曝气系统在低碳源期的总氮去除率较间歇曝气系统高17.92%, 反硝化碳源利用率较间歇曝气系统高44.29%, 连续曝气系统的总氮的去除率与反硝化碳源利用率高于间歇曝气系统, 应对低碳源污水的效果更好.
(2) 低碳源条件下连续曝气系统和间歇曝气系统脱氮功能菌相对丰度分别为5.44%和1.98%, 并且连续曝气的供氧模式使Clostridium sensu stricto、Comamonas和Acinetobacter等功能微生物在低碳源条件下的生长更好, 进而导致连续曝气系统具有较好的低碳源污水处理性能.
(3) 基于16S rRNA测序功能预测, 连续曝气系统中微生物的膜输送功能、碳水化合物代谢功能以及细胞组成功能均强于间歇曝气系统, 这导致连续曝气的供氧模式下脱氮功能微生物生长代谢较旺盛, 因此连续曝气系统可以更好地处理低碳源污水.
| [1] | Wang X X, Wang S Y, Xue T L, et al. Treating low Carbon/Nitrogen (C/N) wastewater in Simultaneous Nitrification-Endogenous Denitrification and Phosphorous Removal (SNDPR) systems by strengthening anaerobic intracellular carbon storage[J]. Water Research, 2015, 77: 191-200. |
| [2] | 郭泓利, 李鑫玮, 任钦毅, 等. 全国典型城市污水处理厂进水水质特征分析[J]. 给水排水, 2018, 44(6): 12-15. |
| [3] | Song J Y, Zhang W, Gao J F, et al. A pilot-scale study on the treatment of landfill leachate by a composite biological system under low dissolved oxygen conditions:performance and microbial community[J]. Bioresource Technology, 2020, 296: 122344. DOI:10.1016/j.biortech.2019.122344 |
| [4] | Terada A, Sugawara S, Yamamoto T, et al. Physiological characteristics of predominant ammonia-oxidizing bacteria enriched from bioreactors with different influent supply regimes[J]. Biochemical Engineering Journal, 2013, 79: 153-161. |
| [5] | Holenda B, Domokos E, Rédey á, et al. Dissolved oxygen control of the activated sludge wastewater treatment process using model predictive control[J]. Computers & Chemical Engineering, 2018, 32(6): 1270-1278. |
| [6] |
赵丽君, 方芳, 郭劲松, 等. 溶解氧对SBR脱氮性能与脱氮方式的影响[J]. 环境工程学报, 2015, 9(3): 1148-1154. Zhao L J, Fang F, Guo J S, et al. Impacts of dissolved oxygen level on nitrogen remval efficiencies and pathways in a sequential batch reactor[J]. Chinese Journal of Environmental Engineering, 2015, 9(3): 1148-1154. |
| [7] | Fan H T, Qi L, Liu G Q, et al. Aeration optimization through operation at low dissolved oxygen concentrations:evaluation of oxygen mass transfer dynamics in different activated sludge systems[J]. Journal of Environmental Sciences, 2017, 55: 224-235. |
| [8] | Wu C Y, Peng Y Z, Li X L, et al. Effect of carbon source on biological nitrogen and phosphorus removal in an anaerobic-anoxic-oxic (A2O) Process[J]. Journal of Environmental Engineering, 2010, 136(11): 1248-1254. |
| [9] | Bernet N, Delgenes N, Akunna J C, et al. Combined anaerobic-aerobic SBR for the treatment of piggery wastewater[J]. Water Research, 2000, 34(2): 611-619. |
| [10] | Zhou N, Dang C Y, Zhao Z R, et al. Role of sludge retention time in mitigation of nitrous oxide emission from a pilot-scale oxidation ditch[J]. Bioresource Technology, 2019, 292: 121961. DOI:10.1016/j.biortech.2019.121961 |
| [11] |
王先宝.基于破源有效利用与曝气模式调控的氧化沟强化脱氮技术研究[D].西安: 西安建筑科技大学, 2015. Wang X B. The study on improvement of nitrogen removal in oxidation ditch by optimization of carbon source utilization and aeration mode[D]. Xi'an: Xi'an University of Architecture and Technology, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10703-1016741594.htm |
| [12] | Li S, Li D, Zhang S R, et al. Effect of aeration modes on simultaneous nitrogen and phosphorus removal and microbial community in a continuous flow reactor with granules[J]. Bioresource Technology, 2019, 294: 122-154. |
| [13] |
黄雪玲, 刘慧敏, 何启帆, 等. 低温条件下不同曝气方式对硫自养湿地脱氮效能的影响[J]. 环境工程学报, 2019, 13(11): 2619-2628. Huang X L, Liu H M, He Q F, et al. Effect of different aeration modes at low temperature on nitrogen removal efficiency of sulfur autotrophic wetland[J]. Chinese Journal of Environmental Engineering, 2019, 13(11): 2619-2628. |
| [14] |
张雯, 邓风, 徐华. 间歇曝气和连续曝气对完全混合式反应器系统脱氮性能的影响[J]. 环境化学, 2013, 32(11): 2176-2185. Zhang W, Deng F, Xu H. The impact of intermittent aeration and continuous aeration on CSTR system denitrification performance[J]. Environmental Chemistry, 2013, 32(11): 2176-2185. |
| [15] |
张哲, 张姚, 刘清华, 等. 连续/间歇曝气MBBR-亚硝化工艺性能及N2O释放特性[J]. 中国环境科学, 2019, 39(12): 5056-5062. Zhang Z, Zhang Y, Liu Q H, et al. MBBR-nitrification process performance and N2O emission characteristics with continuous/intermittent aeration regimes[J]. China Environmental Science, 2019, 39(12): 5056-5062. |
| [16] |
巩有奎, 李永波, 彭永臻. 不同曝气方式下亚硝化反应器的启动及N2O释放[J]. 工业水处理, 2019, 39(4): 21-24, 57. Gong Y K, Li Y B, Peng Y Z. Start-up and N2O emission of the nitrosation reactor in different aeration modes[J]. Industrial Water Treatment, 2019, 39(4): 21-24, 57. |
| [17] | 国家环境保护总局. 水和废水监测分析方法[M]. ((第四版)). 北京: 中国环境科学出版社, 2002. |
| [18] |
殷旭东, 黄梅, 毛玉凤, 等. 分段进水一体化工艺同步硝化反硝化脱氮影响因素研究[J]. 工业用水与废水, 2018, 49(6): 14-18. Yin X D, Huang M, Mao Y F, et al. Research on influencing factors of nitrogen removal by simultaneous nitrification and denitrification with step-feed integrated process[J]. Industrial Water & Wastewater, 2018, 49(6): 14-18. |
| [19] | Jenni S, Vlaeminck S E, Morgenroth E, et al. Successful application of nitritation/anammox to wastewater with elevated organic carbon to ammonia ratios[J]. Water Research, 2014, 49: 316-326. |
| [20] | Zhang J, Wang X X, Liu X Y, et al. Combining partial nitrification and post endogenous denitrification in an EBPR system for deep-level nutrient removal from low Carbon/Nitrogen (C/N) domestic wastewater[J]. Chemosphere, 2018, 210: 19-28. |
| [21] |
白婷婷.低碳源污水硝化及固体碳源反硝化系统的微生物特性[D].重庆: 重庆大学, 2016. Bai T T. Microbiological characteristics of nitrification and solid-phase denitrification system for low-carbon sewage[D]. Chongqing: Chongqing University, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10611-1016908142.htm |
| [22] |
罗晓, 郑向阳, 赵丛丛, 等. A/O工艺中污泥浓度对微生物群落结构的影响[J]. 中国环境科学, 2018, 38(1): 275-283. Luo X, Zheng X Y, Zhao C C, et al. Effects of sludge concentration on microbial community structure in A/O process[J]. China Environmental Science, 2018, 38(1): 275-283. |
| [23] |
郑林雪, 李军, 胡家玮, 等. 同步硝化反硝化系统中反硝化细菌多样性研究[J]. 中国环境科学, 2015, 35(1): 116-121. Zheng L X, Li J, Hu J W, et al. Analysis of denitrifying bacteria community composition in simultaneous nitrification and denitrification systems[J]. China Environmental Science, 2015, 35(1): 116-121. |
| [24] | Kanehisa M, Sato Y, Kawashima M, et al. KEGG as a reference resource for gene and protein annotation[J]. Nucleic Acids Research, 2016, 44(D1): D457-F462. |
| [25] | Wang Z B, Liu X L, Ni S Q, et al. Weak magnetic field:A powerful strategy to enhance partial nitrification[J]. Water Research, 2017, 120: 190-198. |
| [26] | Reuscher S, Fukao Y, Morimoto R, et al. Quantitative proteomics-based reconstruction and identification of metabolic pathways and membrane transport proteins related to sugar accumulation in developing fruits of pear (Pyrus communis)[J]. Plant and Cell Physiology, 2016, 57(3): 505-518. |
| [27] | Herring L E, Grant K G, Blackburn K, et al. Development of a tandem affinity phosphoproteomic method with motif selectivity and its application in analysis of signal transduction networks[J]. Journal of Chromatography B, 2015, 988: 166-174. |
| [28] | Guo K, Liu Y K, Zhou H J, et al. Involvement of protein kinase C β-extracellular signal-regulating kinase1/2/p38 mitogen-activated protein kinase-heat shock protein 27 activation in hepatocellular carcinoma cell motility and invasion[J]. Cancer Science, 2008, 99(3): 486-496. |