 2020, Vol. 41
2020, Vol. 41



2. 西南科技大学环境与资源学院, 绵阳 621010
2. School of Environment and Resources, Southwest University of Science and Technology, Mianyang 621010, China
水力发电具有廉价、清洁的巨大优势, 在全球气候变化的大背景下具有庞大的开发潜力.然而, 水电站的建造会破坏河流生态环境, 如筑坝破坏河流连通性, 改变河流原有的水文和水力学特性, 筑坝形成的水库变为静水环境, 水库下游则为受电站泄流控制的人工流量过程, 使下游河道中河流理化条件发生变化[1].水生生物的生命活动依赖于其栖息地的环境, 河流理化性质的变化进而对生物群落产生影响, 改变各种水生生物的数量和群落结构, 并影响它们的生命活动.大型底栖无脊椎动物(benthic macroinvertebrate, 下称底栖动物)位于河流生态系统食物链的中间环节, 因其活动范围大、对环境变化反应灵敏、监测成本低的特点, 被广泛应用于评估河流生境状况, 是研究以修建水电站为代表的人类活动对河流生境的影响的常用指示生物.
根据集中水头方式的差异, 水电站有坝式水电站和引水式水电站之分.国内外学者对坝式水电站的生态影响已有大量研究.筑坝形成的水库内流速低、泥沙沉积和水质变化, 导致库区内静水生物增加[2]; 坝下水温偏低, 水质的变化和营养物质的减少使敏感物种减少, 耐污物种增加, 总生物密度增加, 但丰度和生物多样性下降[3].另外, 如果坝式水电站承担日调度任务, 则会引起下游流量的剧烈波动, 显著减少底栖动物的数量, 改变群落结构[4].这些影响会随着栖息地到坝址距离的增加而有所减弱[5, 6].而引水式电站对河流的连通性影响较小, 但形成的减水段内流量减少, 会造成岸边底质裸露、植被贫瘠, 并影响部分水质参数.栖息地生境因子的变化进而影响河流底栖动物的生物密度、功能摄食类群等群落结构特征[7].
水电站的梯级开发会加剧生境的破碎化[1].现有研究多关注坝式水电站形成的梯级水库[2, 8, 9], 对实际开发中常见的梯级引水式电站的研究则较少.已有的一些研究指出, 电站库区内、减水段和下游混合段的生物多样性和生物量有所不同[10], 底栖动物摄食功能类群随河流生境物理因子的变化发生改变[11].
以往的研究多关注大坝附近或者引水段的底栖动物变化, 研究对象仅限于单个电站或者2~3个电站的梯级电站, 尚缺乏长距离的、梯级电站作用下的底栖动物的沿程变化规律研究.因此, 本文的研究目的在于:①探究梯级电站影响下的大型底栖动物群落结构沿程变化规律; ②比较引水式水电站和坝式水电站对底栖动物群落结构的影响, 分析不同水电站影响底栖动物群落结构的生境因子.
1 材料与方法 1.1 研究区与采样点布置牡丹江位于黑龙江省东南部, 是松花江干流右岸最大的支流, 全长约726 km, 流域面积37 023 km2, 年平均径流量168 m3·s-1, 主要支流有珠尔多河、海浪河[12].本文的研究范围从牡丹江中游的镜泊湖坝下起至石岩电站, 流长约56.5 km.
河段内共有水电站7座, 其中镜泊湖电站为混合式电站, 下辖新旧两个电站, 旧电站厂房位于堤坝后方, 新电站为地下厂房, 通过从镜泊湖水库中引水进行发电, 尾水经过沿河岸的管道汇入牡丹江, 在大坝和出水口之间形成一段流量较低的河段; 红卫一、红卫二和红农这3个电站为坝式水电站, 依次位于这段低流量河段内, 发电的水主要来自镜泊湖瀑布溢流和渗漏的水; 下游阿堡电站建有拦河坝形成阿堡水库, 江西-渤海电站为引水式水电站, 从阿堡水库库区引水发电, 并在河道内形成一段减水段.
为探究底栖动物的沿程变化规律以及受水电站运行的影响, 在研究河段内沿程布置了21个采样点(位置见图 1, 名称见表 1).红卫一、红卫二和红农电站的挡水建筑物上下游1km内各有1个采样点; 镜泊湖电站有2个出水口, 第1个出水口下游及第2个出水口的上下游各有1个采样点; 阿堡电站拦河坝上下游各有1个采样点.江西-渤海电站的引水段、出水口处以及下游各有1个采样点; 石岩电站库区内无采样点, 下游有1个采样点.

|
图 1 研究区域及采样点位置示意 Fig. 1 Sketch of the study area and sampling sites |
|
|
表 1 各采样点的编号与名称 Table 1 Numbers and names of the sampling sites |
1.2 采样方法
2014年秋天对研究区域的底栖动物进行了野外采样.在每个采样点选择3个位置采样, 在计数时取平均值.采样方法为踢网法, 使用1 m×1 m、网孔直径为0.375 mm的踢网, 采样面积1 m2.将采集的样品经孔径为0.5 mm的网筛筛洗后分别装袋, 带回实验室进行鉴定与计数.将采集的样品置于白瓷盘上, 用镊子挑选出底栖动物并使用75%酒精固定.参考底栖动物图谱在显微镜下对底栖动物进行鉴定, 区分至尽可能低的分类单元(科或属), 按照分类计数、称重.底栖动物中的蜉蝣目(Ephemeroptera)、襀翅目(Plecoptera)和毛翅目(Trichoptera)动物是对水体污染非常敏感的动物, 因此将这3种动物(合称EPT)归为一类, 统计其总数[13].
在底栖动物采样的同时, 测量栖息地的生境因子, 包括水深、流速、河流底质组成和水质指标等[14].使用水深探杆测量采样点的水深, LS300便携式流速仪测量流速, CT-6023 pH计测量pH, YSI Pro ODO溶氧仪测量水体溶氧含量.这些生境因子取3个位置测量值的均值.同时, 在每个采样点使用水样采集器采集水样, 装入统一规格聚乙烯贮样容器, 测定水样水质, 包括COD、高锰酸盐指数、氨氮、总磷和总氮等指标.
1.3 数据分析无重复双因素分析:对河段内除石岩电站的5座电站的上下游共10个采样点的Margalef丰富度指数、物种丰度、底栖动物密度和EPT密度进行无重复双因素分析, 以探究底栖动物受梯级电站影响的沿程变化和采样点相对于水电站位置的影响.其中Margalef指数(IM)反映一个区域的物种丰富程度, 能较好地区分群落差异.其计算公式为[13]:
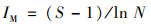
|
(1) |
式中, S为物种丰度, N为采样点的底栖动物总数量.
凝聚法聚类分析:对所有采样点的物种组成进行凝聚法聚类分析, 以比较各采样点群落组成的相似程度.衡量各采样点物种组成相似度的指标为Bray-Crutis距离(dBC):
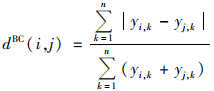
|
(2) |
式中, n为采样点中物种的总数, yi, k和yj, k分别为第i和j个采样点中第k个物种的数量.Bray-Crutis距离的取值范围为0~1, 0代表两个采样点的物种组成完全相同, 1则代表完全不同.采用组平均法进行聚类分析[15].
典范对应分析:对河段内除石岩电站的5座电站的上下游的采样点进行典范对应分析, 分析与底栖动物组成相关的生境因子以及生境因子在不同采样点的特点.
2 结果与分析 2.1 底栖动物群落结构状况在研究范围内, 共采集到物种4纲14目32科, 包括昆虫纲、寡毛纲、双壳纲和腹足纲.其中以昆虫纲动物最多, 共9目26科, 优势物种为原石蛾科(Rhyacophilidae)动物.各采样点采集到的底栖动物的个体数相差较大.采集到底栖动物个体总数最少的为S21石岩电站下游采样点, 仅采集到底栖动物4只; 最多的为S18哈达湾采样点, 共采集到182只.采集到EPT最少的是S3红卫二库区采样点和S21石岩电站下游采样点, 没有采集到EPT; 最多的为S16江西-渤海电站下游采样点, 共采集到107只.
图 2展示了各采样点的底栖动物密度、EPT密度、物种丰度与Margalef指数的沿程变化.从河流整体的变化趋势上看, 直至S16江西-渤海电站下游采样点, 底栖动物的密度、丰度与Margalef丰富度指数有沿程增加的趋势, 从S17三陵桥采样点开始则有所下降.这一下降主要表现为, 在底栖动物密度较高的S17三陵桥采样点和S18哈达湾采样点采集到的底栖动物以潜水蝽科动物为主, 分别占采样点采集到总底栖动物数量的41%和78%.单一物种的巨大优势影响了栖息地的物种丰富度.

|
图 2 各采样点的底栖动物密度、EPT密度与物种丰度和Margalef指数 Fig. 2 Total benthic macroinvertebrate density, EPT density, and richness and Margalef index of the sampling sites |
为探究不同类型的水电站对底栖动物的影响, 选取坝式水电站库区内(或引水式电站减水段内)和下游段的采样点, 比较底栖动物数量与敏感物种数量的变化.镜泊湖电站与石岩电站库区内无采样点, 因此比较除镜泊湖电站和石岩电站的5座水电站.除了S6红农电站下游EPT低于库区内, 其他电站的底栖动物密度和EPT密度都在下游采样点增加(表 2).
|
|
表 2 各电站库区(或减水段内)采样点底栖动物密度与EPT密度与下游采样点的对比/ind.·m-2 Table 2 Comparison of total benthic macroinvertebrate and EPT between sites in the reservoir (or depleted reach) and sites downstream of the plant/ind.·m-2 |
将这5个水电站按照上下游顺序排列, 分别比较电站的库区内(或减水段内)和下游段的底栖动物密度、丰度及EPT密度.如图 3所示, 从上游镜泊湖始至江西-渤海电站, 底栖动物的密度与丰度有增加的趋势.除S16江西-渤海电站下游的采样点的EPT密度相较上游减水段显著增加以外, 其余4个电站的EPT密度沿程未表现出明显的增减趋势.除江西-渤海电站, 其他各电站的库区内底栖动物的丰度和数量均低于上一电站下游采样点处的值; 江西-渤海电站减水段(S14)的底栖动物丰度、总密度高于阿堡电站下游(S13)的值.下游采样点的物种丰富度均高于库区(减水段内)采样点.

|
图 3 牡丹江上5座电站的库区内(或减水段内)和下游段采样点的生物总密度、EPT密度、物种丰度和Margalef指数 Fig. 3 Comparison of total benthic macroinvertebrate density, EPT density, richness, and Margalef index of five hydropower plants between sites in the reservoir (or depleted reach) and sites downstream of the plants |
根据地表水环境质量标准(GB 3838-2002), 研究江段的水质整体为Ⅳ类水, 个别采样点的水质为Ⅴ类或劣Ⅴ类.S5(红农电站库区)、S12(阿堡电站库区)和S15(江西-渤海电站出水口)由于总氮含量较高(表 3), 为Ⅴ类水.S17(三陵桥)和S18(哈达湾)水样氨氮与总氮超标, 为劣Ⅴ类水.
|
|
表 3 主要监测点的生境因子 Table 3 Environmental variables at the main sampling sites |
除S15江西-渤海电站出水口采样点, 各采样点的流速不超过0.6 m·s-1, 在库区或减水段内采样点较低, 在电站下游则会相对其上游有所增加.除S15采样点外, 底栖动物丰度、密度与流速具有相近的变化趋势.而在S15采样点处, 受到江西-渤海电站发电尾流的直接影响, 流速为全部采样点中的最大值(1.50 m·s-1), 过高的流速不利于底栖动物的附着与生存, 因此采集到的底栖动物明显减少, 总密度仅22 ind.·m-2, 丰度(S=4)和物种丰富度(Margalef指数=0.97)也大幅下降(图 4).

|
图 4 各采样点底栖动物密度、丰度与流速的沿程变化趋势对比 Fig. 4 Trends along the study reach of total benthic macroinvertebrate density, richness, and flow velocity |
各采样点底栖动物的数量与群落结构与其相对于水电站的位置有关, 并存在一定的沿程变化的趋势(图 3和表 2).因此对5座电站的库区内(或减水段内)和下游段采样点进行无重复双因素分析, 以分析沿程变化与水电站对各采样点底栖动物的影响.结果显示, 与沿程位置显著相关的是物种丰度和底栖动物密度.电站相对位置则与Margalef指数、物种丰度和底栖动物密度显著相关.EPT密度与这两个因素的相关性均不显著(表 4).
|
|
表 4 无重复双因素分析结果1) Table 4 Results of two-way analysis of variance (ANOVA) without repetition |
2.4 凝聚法聚类分析
使用凝聚法聚类分析来比较各采样点的群落相似程度.聚类分析结果表明, S21石岩电站下游采样点与其余各采样点的差异很大.去除S21, 其余各采样点被分到2个主要分支下.为研究水电站的影响, 仍然对5座电站库区/减水段内和下游的采样点进行分析.采样点S1、S3、S5、S6和S12位于同一分支下(记为G1), S2、S4、S13、S14和S16位于另一分支下(记为G2).G1中除S6红农电站下游采样点, 均为电站上游库区(或减水段内)采样点; G2中除S14江西-渤海电站减水段采样点, 均为电站下游采样点(图 5).

|
图 5 凝聚法聚类分析结果 Fig. 5 Results of hierarchical clustering analysis |
G1采样点中采集到的底栖动物主要为耐污性较强的物种, 敏感物种(耐污值为1~3)的数量占总数的32.2%, 在S6和S12采集到了耐污值为9的霍甫水丝蚓, 另外在S1、S3和S5采集到了较耐污的摇蚊亚科和扁泥甲科动物.在G1采样点中零星采集到不耐污物种, 但数量较少, 且没有在多个采样点都出现的物种.G2采样点采集到的底栖动物总量远高于G1, 采集到的物种中有对水质要求较高的蜉蝣科、纹石蛾科、原石蛾科和钩虾科, 敏感物种数量占总数的38.4%;部分点也有摇蚊亚科、椎实螺科等耐污性中等的生物, S13和S16出现了耐污性强的颤蚓科动物.两组采样点中, 对水质敏感的EPT数量占底栖动物总数量的比例有显著区别(P=0.016).总体而言G2组采样点的底栖动物数量更大, 且多样性更好, 敏感物种的比例更高(图 6).

|
图例中的数值表示该耐污值的底栖动物数量 图 6 G1和G2两组采样点中不同耐污值底栖动物占总数量的比例 Fig. 6 Percentages of species of different tolerance to pollution in the G1 and G2 sites |
G1和G2采样点的pH、氨氮含量、高锰酸盐指数、总氮和总磷没有显著不同, COD存在显著区别(P=0.028), G1、G2两组的平均COD分别为14.72 mg·L-1和14.16 mg·L-1.EPT所占比例与COD具有较强的负相关性(P=-0.826).另外, 虽然方差分析认为两组的流速无显著不同, 但G1组内除S6红农电站下游流速较高外, 其余采样点的流速均低于G2组内采样点的流速, 两组的平均流速分别为0.18 m·s-1和0.36 m·s-1(表 5).
|
|
表 5 对G1和G2两组采样点, 对各水质指标与EPT百分比进行方差分析的P值1) Table 5 ANOVA P-value of water quality variables and EPT% between G1 and G2 |
2.5 典范对应分析
对5个水电站附近的采样点进行典范对应分析.由于部分物种仅在少数采样点出现, 且采集到的数量少, 因此在排序图中仅标出权重不低于20%的物种(图 7).前4个排序轴共解释了物种与生境因子相互关系的79.5%, 其中第一排序轴与第二排序轴分别解释了28.2%和22.5%.对第一排序轴作用最大的生境因子是COD(典范系数b=1.052 7), 对第二排序轴作用最大的生境因子是高锰酸盐指数(b=1.452 9)和流速(b=1.237 7). COD与第一排序轴强相关(相关系数r=0.973 4);与第二排序轴弱相关(r=0.002 3).流速与第一排序轴有较强的负相关性(r=-0.255 1), 与第二排序轴有较强的正相关性(r=0.493 7).

|
图 7 典范对应分析结果 Fig. 7 Results of canonical correspondence analysis |
典范对应分析中采样点的分布与聚类分析的结论吻合, G1组内的采样点均分布在y轴左侧, G2组内的采样点分布在y轴右侧.对比不同水电站附近的采样点, 在坝式水电站库区内的采样点(S1、S3、S5和S12)均位于第四象限, 以低流速和较高的COD为特征; 而引水式水电站减水段内的采样点S14则位于第三象限, 流速中等, COD低, 与S2、S16和S4等位于电站下游段的采样点更接近.
3 讨论 3.1 生境因子对底栖动物分布的影响G1与G2两组采样点之间的底栖动物群落结构组成存在显著区别, 其中G2组采样点的物种更为丰富、敏感物种更多、偏好较高流速的物种更多, 如纹石蛾科和原石蛾科动物, 分别占总数量的27.6%和26.4%.引起群落结构差异的主要生境因子可能为COD和流速.COD与敏感物种的数量相关[16], 流速则造成了G2中偏好高流速物种的增加.研究河段内底栖动物丰度、密度与流速相近的变化趋势, 也说明了流速对底栖动物群落结构的影响.过低的水流流速会降低生产力, 限制物质和能量的交换, 而高流速则会产生过大的水流剪切力, 使得部分种类的底栖动物难以固着在河床底质上, 不利于其生存.一般认为超过1.2 m·s-1的流速会限制大多数生物的生存[13].
有研究认为河道流量是影响底栖动物栖息地环境的重要因素, 河段内流量的补充有利于生态环境恢复[13, 17].在本研究2个流量减少的河段中, 江西-渤海电站下游采样点的底栖动物群落结构指标高于减水段内; 而镜泊湖电站出水口下游底栖动物群落结构的恢复有一定滞后, 出水口下游的2个采样点(S8和S9)与出水口上游低流量河段内的采样点相比, 底栖动物总量与丰度并无明显增加, 直至更下游的S10五七大桥采样点, 底栖动物总量、丰度、EPT密度才有所增加.造成这一滞后的原因可能与镜泊湖电站间歇式发电有关.镜泊湖水电站是黑龙江省电网主要调峰电站[18], 下游水流条件频繁变化, 影响了生活于此的大型底栖动物[11].在经过河道调蓄作用削弱水流频繁变化的影响后, 则能观察到底栖动物由于河道水量增加而有所恢复的现象.然而, S14江西-渤海电站减水段采样点大型底栖动物物种丰富性更高, 聚类分析显示该点的群落组成更接近生境已有所恢复的下游采样点, 这可能是由于河床的阶梯状结构[19]使采样点处产生了较大的流速(0.28 m·s-1), 创造出了良好的栖息地条件.
3.2 不同水电站对河流生态的影响坝式水电站和引水式电站以不同的机制影响河流底栖动物.坝式水电站对河流生态的影响主要与水温、水质、流速以及营养物质的含量有关.而引水式电站的影响主要与以下两方面有关:河道内挡水建筑物的建设和发电引水导致流量减少[7].引水式电站对河流水化学指标的影响不显著, 但显著改变了流速、水深、河道湿周等物理生境因子, 使减水段中动水栖息地缩减、加剧种间竞争与种内斗争、破坏滨河植被, 进而影响底栖动物群落[7, 11, 17].
对比研究河段中的坝式水电站和引水式水电站, 坝式水电站附近的采样点相对于电站的上下游关系, 与其在聚类分析中的分组基本吻合; 而对引水式电站江西-渤海电站, 其减水段采样点(S14)的底栖动物数量和多样性虽然也都低于下游采样点(与坝式水电站类似), 但聚类分析、方差分析和典范对应分析的结果都显示, S14的群落组成与流速、COD等生境因子更接近电站下游采样点.可见就影响程度而言, 虽然引水式电站改变了底栖动物栖息地的部分生境因子, 但对生境因子和底栖动物造成的影响均弱于坝式水电站, 底栖动物的数量和多样性相较于坝式水电站库区内更高.
3.3 底栖动物分布的沿程变化图 3和图 5展示了研究河段内底栖动物的沿程变化.直至S16江西-渤海电站下游采样点, 底栖动物的密度、丰度均有沿程增加的趋势.这一增加的总体趋势主要与两个因素有关:①研究河段由镜泊湖电站和江西渤海电站的减水段和下游段交替组成, 下游段内流量增加, 使生态环境得以恢复; ②随着到拦河建筑物或电站出水口的距离增加, 水电站运行产生的频繁扰动等影响因素沿程逐渐减弱[4, 17, 20].因此从研究河段整体来看, 底栖动物的密度和物种多样性沿程逐渐增加.而在研究河段末端, 底栖动物丰度和密度的整体下降趋势可能与水质的恶化有关.S18哈达湾采样点的水质最差, 虽然在该点采集到了最多的底栖动物, 但其中78%为潜水蝽科动物(同目动物的耐污值为5~8, 属于一般耐污~耐污物种), EPT的数量反而下降.水质恶化导致了该耐污物种的优势和EPT的减少.下游河段水质逐步恢复, 但生态系统的恢复则有所滞后, 加之受石岩电站运行干扰, 并未恢复至上游的状态.
在河流的全尺度上, 不同河段所处地区自然环境的差异, 造成了河流上、中、下游栖息地生境因子(如水温、河道宽度、流速和水深等)的差别, 造就了底栖动物分布的沿程变化[21].这一差别在较大的河流中表现得更明显.而在本研究关注的较小尺度的河段内, 宏观自然环境的差异不大, 底栖动物的分布则更多地取决于河段内修建水电站等设施、水质变化和能量输入[10]等因素.
4 结论(1) 牡丹江江段内梯级电站的建设对多种生境因子造成了影响, 从而改变了江段内底栖动物的群落组成.电站的建设改变了河道内的水质参数(如COD)和流速, 反映在EPT所占比例的差异、底栖动物的密度和丰度的沿程变化上, 造成底栖动物群落组成的不同.
(2) 比较坝式水电站与引水式水电站对底栖动物的影响, 引水式电站同样会改变河流减水段内底栖动物栖息地的水动力条件和水质条件, 对底栖动物(尤其是敏感物种)的生存产生影响, 但其造成的影响弱于坝式水电站.
(3) 随着电站运行的影响沿程逐渐减弱以及河道内流量的补充, 下游生态环境逐步恢复, 底栖动物的密度和丰度有沿程增加的趋势.
| [1] |
毛战坡, 王雨春, 彭文启, 等. 筑坝对河流生态系统影响研究进展[J]. 水科学进展, 2005, 16(1): 134-140. Mao Z P, Wang C Y, Peng W Q, et al. Advances in effects of dams on river ecosystem[J]. Advances in Water Science, 2005, 16(1): 134-140. DOI:10.3321/j.issn:1001-6791.2005.01.024 |
| [2] |
李晋鹏, 董世魁, 彭明春, 等. 梯级水坝运行对漫湾库区底栖动物群落结构及分布格局的影响[J]. 应用生态学报, 2017, 28(12): 4101-4108. Li J P, Dong S K, Peng M C, et al. Effects of cascading hydropower dams operation on the structure and distribution pattern of benthic macroinvertebrate assemblages in Manwan Reservoir, Southwest China[J]. Chinese Journal of Applied Ecology, 2017, 28(12): 4101-4108. |
| [3] | Wu H P, Chen J, Xu J J, et al. Effects of dam construction on biodiversity:a review[J]. Journal of Cleaner Production, 2019, 221: 480-489. DOI:10.1016/j.jclepro.2019.03.001 |
| [4] | Parasiewicz P, Schmutz S, Moog O. The effect of managed hydropower peaking on the physical habitat, benthos and fish fauna in the River Bregenzerach in Austria[J]. Fisheries Management and Ecology, 1998, 5(5): 403-417. DOI:10.1046/j.1365-2400.1998.550403.x |
| [5] | Chaves-Ulloa R, Umaña-Villalobos G, Springer M. Downstream effects of hydropower production on aquatic macroinvertebrate assemblages in two rivers in Costa Rica[J]. Revista de Biologia Tropical, 2014, 62(S2): 179-201. DOI:10.15517/rbt.v62i0.15787 |
| [6] | Mwedzi T, Bere T, Siziba N, et al. Longitudinal macroinvertebrate assemblages in contrasting discontinuities:the effects of damming in tropical streams[J]. African Journal of Ecology, 2016, 54(2): 183-194. DOI:10.1111/aje.12281 |
| [7] | Anderson D, Moggridge H, Warren P, et al. The impacts of "run-of-river' hydropower on the physical and ecological condition of rivers[J]. Water and Environment Journal, 2015, 29(2): 268-276. DOI:10.1111/wej.12101 |
| [8] | Jorcin A, Nogueira M G. Benthic macroinvertebrates in the Paranapanema Reservoir cascade (southeast Brazil)[J]. Brazilian Journal of Biology, 2008, 68(4): 1013-1024. |
| [9] | Lacerda Dos Santos N C, García-Berthou E, Dias J D, et al. Cumulative ecological effects of a Neotropical reservoir cascade across multiple assemblages[J]. Hydrobiologia, 2018, 819(1): 77-91. DOI:10.1007/s10750-018-3630-z |
| [10] |
郭伟杰, 赵伟华, 王振华. 梯级引水式水电站对底栖动物群落结构的影响[J]. 长江科学院院报, 2015, 32(6): 87-93. Guo W J, Zhao W H, Wang Z H. Effects of cascade diversion-type hydropower stations on benthic macroinvertebrate Communities[J]. Journal of Yangtze River Scientific Research Institute, 2015, 32(6): 87-63. |
| [11] |
傅小城, 唐涛, 蒋万祥, 等. 引水型电站对河流底栖动物群落结构的影响[J]. 生态学报, 2008, 28(1): 45-52. Fu X C, Tang T, Jiang W X, et al. Impacts of small hydropower plants on macroinvertebrate communities[J]. Acta Ecologica Sinica, 2008, 28(1): 45-52. DOI:10.3321/j.issn:1000-0933.2008.01.005 |
| [12] |
王皓冉, 陈永灿, 刘昭伟, 等. 牡丹江中游底栖动物分布及其与栖境因子的关系[J]. 中国环境科学, 2015, 35(4): 1197-1204. Wang H R, Chen Y C, Liu Z W, et al. Correlation analysis of macroinvertebrate community and habitat factors in the middle reaches of Mudan River[J]. China Environmental Science, 2015, 35(4): 1197-1204. |
| [13] |
段学花, 王兆印, 徐梦珍. 底栖动物与河流生态评价[M]. 北京: 清华大学出版社, 2010: 12-55. Duan X H, Wang Z Y, Xu M Z. Benthic macroinvertebrate and application in the assessment of stream ecology[M]. Beijing: Tsinghua University Press, 2010: 12-55. |
| [14] | Kelly D J, Hayes J W, Allen C, et al. Evaluating habitat suitability curves for predicting variation in macroinvertebrate biomass with weighted usable area in braided rivers in New Zealand[J]. New Zealand Journal of Marine and Freshwater Research, 2015, 49(3): 398-418. DOI:10.1080/00288330.2015.1040424 |
| [15] | 覃林. 统计生态学[M]. 北京: 中国林业出版社, 2009: 116-128. |
| [16] | 迟国梁, 童晓立.基于典范对应分析的大型底栖动物群落与水环境因子关系研究[A].见: 2015第七届全国河湖治理与生态文明发展论坛论文集[C].广州: 中国水利技术信息中心, 2015. http://cpfd.cnki.com.cn/Article/CPFDTOTAL-SLJS201504001073.htm |
| [17] | Wang H R, Chen Y C, Liu Z W, et al. Effects of the "run-of-river" hydro scheme on macroinvertebrate communities and habitat conditions in a mountain river of Northeastern China[J]. Water, 2016, 8(1): 31. |
| [18] | 周全, 吕婷婷, 李晓军. 镜泊湖水库综合利用要求及调度研究[J]. 东北水利水电, 2012, 30(10): 17-18, 35. DOI:10.3969/j.issn.1002-0624.2012.10.008 |
| [19] | Wang Z Y, Lee J H W, Melching C S. River dynamics and integrated river management[M]. Berlin, Heidelberg: Springer, 2015: 154-174. |
| [20] | Holt C R, Pfitzer D, Scalley C, et al. Longitudinal variation in macroinvertebrate assemblages below a large-scale hydroelectric dam[J]. Hydrobiologia, 2015, 755(1): 13-26. DOI:10.1007/s10750-015-2212-6 |
| [21] | Jiang X M, Xiong J, Xie Z C. Longitudinal and seasonal patterns of macroinvertebrate communities in a large undammed river system in Southwest China[J]. Quaternary International, 2017, 440: 1-12. |