 2020, Vol. 41
2020, Vol. 41



2. 武外英中学校, 武汉 430022;
3. 湖北省农业科学院植保土肥研究所, 武汉 430064
2. Wuhan Britain-China School, Wuhan 430022, China;
3. Institute for Plant Protection and Soil Science, Hubei Academy of Agricultural Sciences, Wuhan 430064, China
人类活动的干扰和磷肥的大量施用加快了自然界中的P元素进入水体的速度, 破坏了磷循环的正常速率, 导致了世界范围内的水体富营养化[1], 并进一步导致了全球范围内夏季藻华的恶化趋势[2].沉水植物是浅水湖泊重要的功能群, 不同沉水植物种类对磷变化的响应体现出明显的种间差异[3~5], 对水体磷富集适应性强的沉水植物种类会逐渐演化为优势种[6].
沉水植物可以通过茎叶拦截和吸附水体中的可溶性磷, 通过根部的吸收作用促进底质中的有机磷的迁移[7, 8].多数研究均以水体中磷的去除率或植物干重全磷表征植物对水体的净化效果, 一般去除率较高, 干重磷含量较高的植物被认定为除磷效果好[4, 9].然而随着沉水植物的衰亡和腐败, 植株生长旺盛时期所积累的磷将重新被释放到水体中.王立志等[10]发现黑藻和菹草在衰亡期会使沉积物和上覆水中各种磷含量显著增加.另外的研究表明水体中被植物吸收或者吸附的磷, 只有少量通过沉淀反应[11]或者植物的生物钙化作用[12, 13]形成CaCO3-P共沉淀, 富集在植株体表面.与CaCO3结合在一起的磷在植株经微生物原位分解后很难被生物重新利用, 因此对水体磷水平不再有影响[14].因此, 钙磷可以表征植物从水体相向基底迁移磷的能力, 比较不同植物的钙磷含量有利于更好地评估植物对水体磷的去除能力[15].
在湖泊生态系统中, 沉水生物的光合作用形成的微环境, 或局部水体Ca2+高度过饱和是控制湖泊CaCO3沉淀的直接原因和形成细胞外CaCO3自生沉淀的主要原因[16].低于50mg·L-1的低Ca2+浓度发生的钙化主要为生物钙化, 高于200mg·L-1的高Ca2+浓度则主要为以生物引发的物理化学钙化[17].此时, 水体SRP的去除可以通过CaCO3晶体生长或CaCO3沉淀时产生CaCO3-P共沉淀而去除[11].在低Ca2+浓度条件下, 光合作用可直接诱导14% CaCO3沉淀[18], CaCO3-P共沉淀除磷作用会因沉水植物光合作用加强而更加明显[19, 20].
粉绿狐尾藻[Myriophyllum aquaticum (Vell.) Verdc.]属于小二仙草科狐尾藻属, 原产地为南美亚马孙河流域[21], 为多年生草本植物, 可挺水或沉水生长, 适应水环境能力强[22], 因此广泛应用于人工湿地的建设和富营养化河道水体的修复.因其茎呈半蔓状, 上部挺出水面, 并伴有叶片匍匐于水面, 能利用空气中的二氧化碳作为碳源, 故不受水下光照和水体无机碳的限制.菹草(Potamogeton crispus L.)为眼子菜属冠层型多年生沉水植物, 分布我国南北各省, 为世界广布种, 也是长江中下游中富营养湖泊的优势种[23].菹草能利用水中的CO2和HCO3-, 属于HCO3-利用型[24].本实验选择粉绿狐尾藻和菹草两种不同生活型的植物为研究对象, 控制光照和磷含量两个因素, 以植物灰分磷和灰分磷组成为切入口, 通过将灰分磷分为水溶性磷、有机磷和钙磷这3个组分, 探究两种植物富集磷的实际能力和植物腐败后对水体磷增加的影响, 以期为选取水体富营养化修复的优势物种提供理论依据.
1 材料与方法 1.1 实验材料与预培养2017年春, 将取自梁子湖的菹草(P. crispus)和取自内沙湖的粉绿狐尾藻(M. aquaticum)栽培在湖北工业大学资源与环境学院后面水池无遮荫的水泥池, 自来水培养.在2018年3月10日截取若干长势良好和叶片完整, 长约5~8 cm的植物顶端, 并用去离子水洗净植株表面, 再用软毛刷清洗植株叶片, 去除植株表面杂质和能与植株竞争营养的其他附着生物.将清洗后的植株分开放置于1 L的玻璃烧杯中, 取水池内的原水用玻璃纤维膜(上海新亚, 0.45 μm)过滤除去藻类与大颗粒物质后添加到放置植株的烧杯中[TP:(0.003 6±0.001 2) mg·L-1, SRP:(0.002 1±0.001 0)mg·L-1].烧杯置于恒温培养箱中, 光照强度为60%光照, 对应光量子强度为66 μmol·(m2·s)-1, 光暗比=12:12, 温度控制为20℃, 预培养一周, 培养期间不更换培养液.
1.2 实验设计预培养一周后, 选取长势状况良好, 已基本适应环境的植物顶端, 将其转入1 L烧杯中, 每个杯中约4~5株, 并记录其重量, 菹草鲜重约3 g, 粉绿狐尾藻鲜重约5 g, 每个烧杯中有不同磷浓度的培养液1 L, 并将其分成两组, 每组置于不同光照的培养箱中.外加磷源的浓度设置3个水平, 分别是无添加磷(0mg·L-1)、中浓度磷(0.2mg·L-1)和高浓度磷(2 mg·L-1);光照强度设置了两个水平, 分别是100%光照和60%光照.无机磷处理通过添加K2HPO4获得, 光照处理通过调节光照培养箱获得.本研究有6种处理(表 1), 平行处理为4个.
|
|
表 1 实验分组设计 Table 1 Experimental group design |
正式培养过程中的培养温度、光照周期设置与预实验保持一致, 实验每隔12 h, 即在每个光周期开始的时候更换培养液, 配置培养液的原水获取与预实验中一致.本实验周期为7 d.实验结束后将植株冲洗, 用纸擦拭, 分析天平称取其鲜重.并检测植物干重全磷和植株灰分磷及灰分磷组成.
1.3 数据获得植物鲜重通过分析天平(分度值0.001 g)测得, 相对生长速率使用公式:
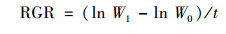
|
式中, RGR为植物相对生长速率[mg·(g·d)-1];W0为实验开始时植物鲜重(mg); W1为实验结束时植物鲜重(mg);t为实验时间(d).植株干重全磷通过钼锑抗比色法(GB 7887-87)测定[25], 然后将部分草样用马弗炉(SX-4-10型箱式电阻炉)进行烧灰处理(450℃, 24 h), 再分别用不同的试剂萃取以获得水溶性磷(H2O-P)、有机磷(NaOH-P)和钙磷(Ca-P).该方法包括:①用50 mL蒸馏水提取样品30 min以获得水溶性P(H2O-P); ②用50 mL 1.0 mol·L的NaOH提取20 h以获得有机P(NaOH-P); ③用50 mL 1.0 mol·L的HCl萃取30 min以释放Ca结合的钙P(Ca-P)[26], 水溶性磷(H2O-P)有机磷(NaOH-P)和钙磷(Ca-P)这3个组分相加得到灰分总磷.萃取后的溶液用过硫酸钾氧化法(GB 11893-89)测得对应磷含量.
1.4 数据分析光照和磷水平对两种植物相对生长速率、干重全磷、灰分磷和各组分的影响通过二元方差分析获得, 处理间的差异通过t-test获得, 所有的分析通过SPSS 23.0进行, 采用Origin 2016绘制图.
2 结果与分析 2.1 相对生长速率两种光照条件下, 2mg·L-1的磷添加显著提高了菹草的相对生长速率.在外加磷质量浓度较低(0和0.2 mg·L-1)时, 菹草的生长速率不受光照强度的影响.在外源性磷质量浓度为2mg·L-1、光照强度为60%时, 菹草的生长速率达到最大[图 1(a)].二元方差分析表明磷质量浓度对菹草的生长速率影响更为显著(69.657%), 见表 2.

|
不同的大写字母代表处理间差异显著, 下同 图 1 不同P添加-光照水平处理条件下的两种植物相对生长速率 Fig. 1 Relative growth rates of two plants under different P-addition-light treatment conditions |
|
|
表 2 外加磷和光照对植物生长速率、磷含量和组成差异的贡献百分比1) Table 2 Percentage of contribution of phosphorus and light to plant growth rate, phosphorus content, and compositional differences |
在光照强度相同时, 粉绿狐尾藻的生长速率随磷浓度的增加呈现上升的趋势.在外加磷浓度相同时, 粉绿狐尾藻的生长速率与光照无关[图 1(b)].二元方差分析表明(表 2)磷浓度对粉绿的生长速率影响更为显著(75%).
2.2 植株灰分磷组成 2.2.1 植物Ca-P在光照强度相同时, 菹草的Ca-P随磷质量浓度的增加呈现上升的趋势.在外加磷质量浓度较低(0和0.2 mg·L-1)时, 光照强度对菹草的Ca-P含量无显著性的影响.在磷2 mg·L-1时, 60%光照强度时菹草的Ca-P含量高于100%光照强度, 也是最大Ca-P含量[图 2(a)].二元方差分析表明(表 2)磷质量浓度对菹草的Ca-P影响更为显著(94.772%).

|
图 2 不同P添加-光照水平处理条件下的两种植物灰分中的钙磷 Fig. 2 Ca-P of two plants under different P-addition-light treatment conditions |
在外加磷质量浓度为2mg·L-1和无外源性磷添加时, 粉绿狐尾藻的Ca-P含量不受光照强度的影响.在外加磷质量浓度为0.2 mg·L-1时, 高光条件有更高的Ca-P含量.两种光照条件下, 最大的Ca-P值均出现在高磷浓度添加(2 mg·L-1).二元方差分析表明磷质量浓度对粉绿狐尾藻Ca-P的影响更为显著(65.019%), 见表 2.在0和0.2 mg·L-1两种质量浓度磷添加时, 菹草的Ca-P均比粉绿狐尾藻小(P < 0.001), 但在外加磷质量浓度为2mg·L-1时, 菹草的Ca-P含量迅速升高, 大于粉绿狐尾藻(P < 0.05).
2.2.2 植物H2O-P在光照强度相同时, 菹草的H2O-P含量随磷质量浓度的增加呈上升趋势.在外加磷质量浓度较低(0和0.2 mg·L-1)时, 菹草的H2O-P含量不受光照强度的影响.在2mg·L-1时, 60%光照强度时菹草的H2O-P含量高于100%光照强度, 也是最大H2O-P含量[图 3(a)].二元方差分析表明(表 2)磷质量浓度对菹草H2O-P含量影响更为显著(94.014%).

|
图 3 不同P添加-光照水平处理条件下的两种植物灰分中的水溶性磷 Fig. 3 H2O-P of two plants under different P-addition-light treatment conditions |
在外加磷质量浓度为0.2 mg·L-1和2 mg·L-1时, 光照对粉绿狐尾藻的H2O-P含量无影响.两种光照情况下, 最大的H2O-P值均出现在高磷浓度添加(2mg·L-1).二元方差分析表明(表 2)磷质量浓度对粉绿狐尾藻的H2O-P影响更为显著(47.875%).在0和0.2 mg·L-1两种质量浓度添加时, 菹草的H2O-P含量比粉绿狐尾藻小(P<0.001).但在外加磷质量浓度为2 mg·L-1时, 菹草的H2O-P含量迅速升高, 大于粉绿狐尾藻(P<0.05).
2.2.3 植物NaOH-P在光照强度相同时, 菹草的NaOH-P含量随磷质量浓度的增加呈上升趋势.在外加磷质量浓度较低(0和0.2 mg·L-1)时, 菹草的NaOH-P含量不受光照强度的影响.在2 mg·L-1时, 60%光照强度时菹草的NaOH-P含量高于100%光照强度, 也是最大NaOH-P含量[图 4(a)].二元方差分析表明(表 2)磷质量浓度对菹草NaOH-P含量影响更为显著(94.689%).

|
图 4 不同P添加-光照水平处理条件下的两种植物灰分中的NaOH-P Fig. 4 NaOH-P levels of two plants under different P-addition-light treatment conditions |
在外加磷质量浓度相同时, 粉绿狐尾藻的NaOH-P含量存在着显著差异, 60%光照强度时粉绿狐尾藻的NaOH-P含量高于100%光照强度.两种光照条件下, 粉绿狐尾藻的NaOH-P含量最大值均出现在2mg·L-1.二元方差分析表明磷质量浓度和光照均对粉绿的NaOH-P含量有显著性影响(磷质量浓度影响占比45.737%;光照强度影响占比44.868%, 表 2).在0和0.2 mg·L-1两种质量浓度添加时, 菹草的NaOH-P含量比粉绿狐尾藻小(P<0.001).但在外加磷质量浓度为2 mg·L-1时, 菹草的NaOH-P含量迅速升高, 大于粉绿狐尾藻(P<0.05).
2.3 植物灰分总磷和植物全磷 2.3.1 植物灰分总磷植物灰分磷由可溶性磷(H2O-P)、有机磷(NaOH-P)和钙磷(Ca-P)组成.
在光照强度相同时, 菹草的灰分总磷含量随磷质量浓度的增加呈上升趋势.在外加磷质量浓度较低(0和0.2mg·L-1)时, 菹草的总磷含量不受光照强度的影响.在2 mg·L-1时, 60%光照强度时菹草的总磷含量高于100%光照强度, 也是最大总磷含量[图 5(a)].二元方差分析表明磷质量浓度对菹草总磷含量影响更为显著(95.681%, 表 2).

|
图 5 不同P添加-光照水平处理条件下的两种植物灰分总磷 Fig. 5 Ash phosphorus content of the two plants under different P-addition-light treatment conditions |
在外加磷质量浓度为0.2 mg·L-1和无外源性磷添加时, 粉绿狐尾藻的总磷含量不受光照强度的影响.两种光照情况下, 最大的总磷值均出现在高磷浓度添加(2 mg·L-1).二元方差分析表明磷质量浓度对粉绿狐尾藻灰分总磷的影响更为显著(85.432%, 表 2).在0和0.2mg·L-1两种质量浓度添加时, 菹草的灰分总磷含量比粉绿狐尾藻小(P<0.05).但在外加磷质量浓度为2 mg·L-1时, 菹草的灰分总磷含量迅速升高, 大于粉绿狐尾藻(P<0.05)
2.3.2 植株全磷在光照强度相同时, 菹草的干重全磷随磷质量浓度的增加呈现上升的趋势.在无外加磷源时, 菹草的全磷在两种光照强度无显著性差异.在较高磷质量浓度(0.2 mg·L-1和2 mg·L-1)时, 光照强度不同, 菹草的全磷含量存在着显著差异, 在外源性添加磷质量浓度为2 mg·L-1、光照强度为60%时, 菹草的全磷含量最高[图 6(a)].二元方差分析表明光照和磷质量浓度对菹草的干重全磷有显著的交互影响, 其中光照强度的影响更为显著(95.551%, 表 2).

|
图 6 不同P添加-光照水平处理条件下的两种植物干重全磷 Fig. 6 Total phosphorus contents of the two plants under different P-addition-light treatment conditions |
2 mg·L-1的磷添加显著提高了粉绿狐尾藻的干重全磷含量.在相同磷质量浓度时, 粉绿狐尾藻的全磷含量不受光照强度的影响.在磷质量浓度较低(0和0.2 mg·L-1)时, 100%光照强度粉绿狐尾藻的全磷含量高于60%光照强度;在高磷质量浓度时, 100%光照强度粉绿狐尾藻的全磷含量低于60%光照强度.在光照强度相同时, 粉绿狐尾藻的全磷随磷质量浓度的增加呈现上升的趋势.二元方差分析表明磷质量浓度对粉绿狐尾藻的干重全磷影响更为显著(70.842%, 表 2).在外源性添加磷质量浓度为2mg·L-1, 光照强度为60%时, 粉绿狐尾藻的全磷含量最高.
3 讨论 3.1 光照水平对植物生长的影响在实验设置的3组生长条件下, 菹草的相对生长速率都高于粉绿狐尾藻.在本实验设置的两组光照条件中, 66 μmol·(m2·s)-1光照条件下菹草生长状况显著好于110 μmol·(m2·s)-1光照条件;而粉绿狐尾藻在两种光照水平下的相对生长速率无显著差异.分析其主要原因是两种植物在水中的生长状态不同, 故其对光照的需求也不同.菹草为多年生沉水草本, 对光照需求较低, 因此较低光照有利于植物生长.而粉绿狐尾藻的多数叶片匍匐在水面, 喜强光照.Tan等[27]的研究表明, 光照强度的降低和过度遮阴将抑制水体中粉绿狐尾藻的生长.
3.2 灰分磷各组分的意义本实验通过不同的处理方法将植物灰分磷分成水溶性磷(H2O-P)、有机磷(NaOH-P)和钙磷(Ca-P)这3个部分.其中, 水溶性磷(H2O-P)用来表征植物对水体磷的吸附作用, 而这种吸附作用是可逆的[12, 13].因此, 沉水植物死亡后, 作为可逆性吸附反应的结果, 水溶性磷(H2O-P)以可溶性磷的形式重新进入水体, 直接被其它水生生物利用, 而有机磷(NaOH-P)则通过微生物的分解作用成为可溶性无机磷[28, 29], 这能解释白洋淀菹草大量死亡后水体磷的急剧上升[30].
与水溶性磷(H2O-P)和有机磷(NaOH-P)相比, 一般情况下钙磷(Ca-P)不易被溶解和吸附, 是生物难以利用的磷, 在植物死亡分解后这部分磷经过一系列的迁移转化最终以稳定的形态埋藏在沉积物中[12~14].朱端卫等[11]的研究发现, 由沉水植物光合作用驱动的钙泵会促进水体磷、CO32-和钙离子共沉淀形成CaCO3-P共沉淀, 从而维持上覆水较低的磷水平.这也是在植株上形成稳定Ca-P复合物的钙化的大型植物轮藻群落内水体可溶性磷含量低, 并且基质中含有大量碳酸盐的主要原因之一[31].因此, 钙磷实际上可以表征植物对水体磷的真正去除能力.陈琦[15]的研究表明无机碳添加通过影响菹草和粉绿狐尾藻灰分中的钙磷来影响水体磷向基质的迁移.在本实验中两种植物菹草和粉绿狐尾藻的钙磷(Ca-P)含量均受到外源性添加磷浓度的影响, 这与陈琦[15]的实验结果一致.在磷浓度为2mg·L-1时两种植物的钙磷(Ca-P)含量达到最高, 印证了较高的磷浓度可以促进钙磷共沉淀的产生[32].
3.3 干重磷和灰分磷的差异作为主要的初级生产者, 水生植物在改善水体环境和维持良好水质中具有不可替代的作用[33].水生植物能直接从水体中吸收N、P等营养物质, 也可通过影响微生物代谢等方式减少水体营养盐的循环, 从而降低水体富营养化水平[8], 杨文斌等的研究表明, 狐尾藻和金鱼藻均可有效吸收上覆水和间隙水中的磷, 使水体磷浓度保持较低水平[35].其中植物自身的代谢吸收是最为主要的方式[32].因此, 植株干重磷含量和磷吸收量通常作为筛选净化水质优势植物的重要指标[35~37].
本实验中, 磷的添加和浓度的变化显著影响了菹草和粉绿狐尾藻的干重磷(表 2).两种植物干重磷更高值均出现在高磷浓度水平(图 6), 说明两种植物能适应不同磷浓度的水体, 且在腐败衰亡前对磷浓度较高水体处理效果好.菹草的干重磷在各种处理下都低于粉绿狐尾藻, 若以干重磷含量作为植物富集水体磷的能力指标, 菹草的富集能力低于粉绿狐尾藻.然而水生植物的腐解是导致水体内源污染的重要环节, 在植物腐败衰亡时, 磷会优于其他元素被大量释放, 加重湖泊污染[30, 38].本研究中, 菹草灰分总磷和钙磷在高磷水平大于粉绿狐尾藻, 如前所述, 钙磷才是植物从水体最终去除的磷, 因此, 更高的钙磷含量表明高磷浓度(2mg·L-1)下菹草比粉绿狐尾藻的磷去除能力更强.这与陈琦等[15, 32]的研究一致, 即比较不同植物的钙磷含量有利于更好地评估植物对水体磷的去除能力.
4 结论(1) 菹草相对生长速率显著高于粉绿狐尾藻;两种植物的最大生长速率均出现在高磷水平, 但由于生长型的差异, 菹草在高磷质量浓度和低光照水平条件下生长速率最大, 而光照对粉绿狐尾藻的生长无影响.
(2) 菹草和粉绿狐尾草在生长过程中均能有效富集水体中的磷, 二元方差分析表明无机磷的添加对两种水生植物的水溶性磷(H2O-P)、有机磷(NaOH-P)和钙磷(Ca-P)均有显著影响.且菹草和粉绿狐尾藻的钙磷(Ca-P)含量更多地受到外源性添加磷的影响, 在磷质量浓度为2 mg·L-1时钙磷(Ca-P)含量均达到最高, 证明磷浓度增加能促进Ca-P共沉淀的生成.
(3) 菹草的干重磷在各种处理下都低于粉绿狐尾藻, 但灰分总磷和钙磷在高磷水平大于粉绿狐尾藻.因此, 仅以植物干重磷比较植物对磷的去除能力容易引起争议, 以植物灰分磷组成和钙磷含量比较植物去除水体磷的能力更为准确.
| [1] | Yuan Z W, Jiang S Y, Sheng H, et al. Human perturbation of the global phosphorus cycle:changes and consequences[J]. Environmental Science & Technology, 2018, 52(5): 2438-2450. |
| [2] | Ho J C, Michalak A M, Pahlevan N. Widespread global increase in intense lake phytoplankton blooms since the 1980s[J]. Nature, 2019, 574(7780): 667-670. DOI:10.1038/s41586-019-1648-7 |
| [3] |
高敏, 刘鑫, 邓建才, 等. 不同水质对沉水植物马来眼子菜主要生理指标的影响研究[J]. 生态环境学报, 2015, 24(11): 1886-1892. Gao M, Liu X, Deng J C, et al. Studies on effects of water quality in different lake zones on main physiological indices of Potamogeton wrightii morong[J]. Ecology and Environmental Sciences, 2015, 24(11): 1886-1892. |
| [4] |
熊剑, 黄建团, 聂雷, 等. 不同营养条件对金鱼藻净化作用及其生理生态的影响[J]. 水生生物学报, 2013, 37(6): 1066-1072. Xiong J, Huang J T, Nie L, et al. The effects of nutrient concentration on purificaion ability and eco-physiology of Ceratophyllum demersum[J]. Acta Hydrobiologica Sinica, 2013, 37(6): 1066-1072. |
| [5] |
蒋鑫焱, 翟建平, 黄蕾, 等. 不同水生植物富集氮磷能力的试验研究[J]. 环境保护科学, 2006, 32(6): 13-16. Jiang X Y, Zhai J P, Huang L, et al. Experimental study on enrichment of nitrogen and phosphor in several hydrophytes[J]. Environmental Protection Science, 2006, 32(6): 13-16. DOI:10.3969/j.issn.1004-6216.2006.06.005 |
| [6] |
吴爱平, 吴世凯, 倪乐意. 长江中游浅水湖泊水生植物氮磷含量与水柱营养的关系[J]. 水生生物学报, 2005, 29(4): 406-412. Wu A P, Wu S K, Ni L Y. Study of macrophytes nitrogen and phosphorus contents of the shallow lakes in the middle reaches of Changjiang river[J]. Acta Hydrobiologica Sinica, 2005, 29(4): 406-412. DOI:10.3321/j.issn:1000-3207.2005.04.009 |
| [7] | Gabrielson J O, Perkins M A, Welch E B. The uptake, translocation and release of phosphorus by Elodea densa[J]. Hydrobiologia, 1984, 111(1): 43-48. DOI:10.1007/BF00007379 |
| [8] |
吴振斌, 邱东茹, 贺锋, 等. 沉水植物重建对富营养水体氮磷营养水平的影响[J]. 应用生态学报, 2003, 14(8): 1351-1353. Wu Z B, Qiu D R, He F, et al. Effects of rehabilitation of submerged macrophytes on nutrient level of a eutrophic lake[J]. Chinese Journal of Applied Ecology, 2003, 14(8): 1351-1353. DOI:10.3321/j.issn:1001-9332.2003.08.031 |
| [9] |
任文君, 田在锋, 宁国辉, 等. 4种沉水植物对白洋淀富营养化水体净化效果的研究[J]. 生态环境学报, 2011, 20(2): 345-352. Ren W J, Tian Z F, Ning G H, et al. Purification efficiency of four species submerged macrophytes for the eutrophic water in Baiyangdian Lake[J]. Ecology and Environmental Science, 2011, 20(2): 345-352. DOI:10.3969/j.issn.1674-5906.2011.02.024 |
| [10] |
王立志, 王国祥. 衰亡期沉水植物对水和沉积物磷迁移的影响[J]. 生态学报, 2013, 33(17): 5426-5437. Wang L Z, Wang G X. Influence of submerged macrophytes on phosphorus transference between sediment and overlying water in decomposition period[J]. Acta Ecologica Sinica, 2013, 33(17): 5426-5437. |
| [11] |
朱端卫, 朱红, 倪玲珊, 等. 沉水植物驱动的水环境钙泵与水体磷循环的关系[J]. 湖泊科学, 2012, 24(3): 355-361. Zhu D W, Zhu H, Ni L S, et al. Relationship between calcium pump driven by submerged macrophytes and phosphorus cycle in water[J]. Journal of Lake Sciences, 2012, 24(3): 355-361. DOI:10.3969/j.issn.1003-5427.2012.03.005 |
| [12] | Kufel L, Biardzka E, Strzałek M. Calcium carbonate incrustation and phosphorus fractions in five charophyte species[J]. Aquatic Botany, 2013, 109: 54-57. DOI:10.1016/j.aquabot.2013.04.002 |
| [13] | Kufel L, Strzalek M, Biardzka E. Site-and species-specific contribution of charophytes to calcium and phosphorus cycling in lakes[J]. Hydrobiologia, 2016, 767(1): 185-195. DOI:10.1007/s10750-015-2498-4 |
| [14] | James W F, Barko J W, Field S J. Phosphorus mobilization from littoral sediments of an inlet region in Lake Delavan, Wisconsin[J]. Archiv fur Hydrobiologie, 1996, 138(2): 245-257. |
| [15] |
陈琦, 黄飞, 刘瑛, 等. 无机碳和磷添加对两种沉水植物磷富集和灰分磷组成的影响[J]. 水生态学杂志, 2019, 40(3): 58-64. Chen Q, Huang F, Liu Y, et al. Effect of adding bicarbonate and phosphorus on phosphorus accumulation in two submerged plants and on the content and fractioning of phosphorous in plant residues[J]. Journal of Hydroecology, 2019, 40(3): 58-64. |
| [16] | 吴丰昌, 万国江. 淡水湖泊碳酸钙自生沉淀生物成因新证据和成因解释[J]. 矿物岩石地球化学通报, 1994(2): 71-72. |
| [17] |
张道勇, 潘响亮, 张京梅. 环境因子对Synechocystis sp.钙化动力学的影响[J]. 矿物岩石地球化学通报, 2008, 27(2): 105-111. Zhang D Y, Pan X L, Zhang J M. Effects of environmental factors on calcification kinetics of Synechocystis sp.[J]. Bulletin of Mineralogy, Petrology and Geochemistry, 2008, 27(2): 105-111. DOI:10.3969/j.issn.1007-2802.2008.02.001 |
| [18] | Pentecost A, Franke U. Photosynthesis and calcification of the stromatolitic freshwater cyanobacterium Rivularia[J]. European Journal of Phycology, 2010, 45(4): 345-353. DOI:10.1080/09670262.2010.492914 |
| [19] | Dittrich M, Koschel R. Interactions between calcite precipitation (natural and artificial) and phosphorus cycle in the hardwater lake[J]. Hydrobiologia, 2002, 469(1-3): 49-57. |
| [20] | House W A. Geochemical cycling of phosphorus in rivers[J]. Applied Geochemistry, 2003, 18(5): 739-748. |
| [21] | Li Z Y, Hsieh C F. New materials of the genus Myriophyllum L. (Haloragaceae) in Taiwan[J]. Taiwania, 1996, 41(4): 322-328. |
| [22] | Hussner A, Meyer C, Busch J. The influence of water level and nutrient availability on growth and root system development of Myriophyllum aquaticum[J]. Weed Research, 2009, 49(1): 73-80. DOI:10.1111/j.1365-3180.2008.00667.x |
| [23] |
高镜清, 熊治廷, 张维昊, 等. 常见沉水植物对东湖重度富营养化水体磷的去除效果[J]. 长江流域资源与环境, 2007, 16(6): 796-800. Gao J Q, Xiong Z T, Zhang W H, et al. Removal efficiency of phosphorus in hypertrophic lake donghu water by common submerged macrophytes[J]. Resources and Environment in the Yangtze Basin, 2007, 16(6): 796-800. DOI:10.3969/j.issn.1004-8227.2007.06.020 |
| [24] | Kahara S N, Vermaat J E. The effect of alkalinity on photosynthesis-light curves and inorganic carbon extraction capacity of freshwater macrophytes[J]. Aquatic Botany, 2003, 75(3): 217-227. |
| [25] | 董鸣. 陆地生物群落调查观测与分析[M]. 北京: 中国标准出版社, 1997. |
| [26] | Dou Z, Toth J D, Galligan D T, et al. Laboratory procedures for characterizing manure phosphorus[J]. Journal of Environmental Quality, 2000, 29(2): 508-514. |
| [27] | Tan B C, He H, Gu J, et al. Effects of nutrient levels and light intensity on aquatic macrophyte (Myriophyllum aquaticum) grown in floating-bed platform[J]. Ecological Engineering, 2019, 128: 27-32. DOI:10.1016/j.ecoleng.2018.12.011 |
| [28] | Maurer M, Abramovich D, Siegrist H, et al. Kinetics of biologically induced phosphorus precipitation in waste-water treatment[J]. Water Research, 1999, 33(2): 484-493. DOI:10.1016/S0043-1354(98)00221-8 |
| [29] | Drizo A, Comeau Y, Forget C, et al. Phosphorus saturation potential:a parameter for estimating the longevity of constructed wetland systems[J]. Environmental Science & Technology, 2002, 36(21): 4642-4648. |
| [30] |
张菊, 邓焕广, 吴爱琴, 等. 东平湖菹草腐烂分解及其对水环境的影响[J]. 环境科学学报, 2013, 33(9): 2590-2596. Zhang J, Deng H G, Wu A Q, et al. Decomposition of Potamogeton crispus and its effect on the aquatic environment of Dongping Lake[J]. Acta Scientiae Circumstantiae, 2013, 33(9): 2590-2596. |
| [31] | Pełechaty M, Pukacz A, Apolinarska K, et al. The significance of Chara vegetation in the precipitation of lacustrine calcium carbonate[J]. Sedimentology, 2013, 60(4): 1017-1035. DOI:10.1111/sed.12020 |
| [32] |
陈琦, 王和云. 钙离子添加对沉水植物菹草(Potamogeton crispus L.)和金鱼藻(Ceratophyllum demersum L.)富集水体磷的影响[J]. 湖泊科学, 2020, 32(2): 406-416. Chen Q, Wang H Y. Effect of calcium addition on phosphorus enrichment capacity of Potamogeton crispus L. and Ceratophyllum demersum L. in water bodies[J]. Journal of Lake Science, 2020, 32(2): 406-416. |
| [33] |
吴振斌, 邱东茹, 贺锋, 等. 水生植物对富营养水体水质净化作用研究[J]. 武汉植物学研究, 2001, 19(4): 299-303. Wu Z B, Qiu D R, He F, et al. Studies on eutrophicated water quality improvement by means of aquatic macrophytes[J]. Journal of WuHan Botanical Research, 2001, 19(4): 299-303. DOI:10.3969/j.issn.2095-0837.2001.04.003 |
| [34] |
王立志, 王国祥, 俞振飞, 等. 沉水植物生长期对沉积物和上覆水之间磷迁移的影响[J]. 环境科学, 2012, 33(2): 385-392. Wang L Z, Wang G X, Yu Z F, et al. Influence of submerged macrophytes on phosphorus transference between sediment and overlying water in the growth period[J]. Environmental Science, 2012, 33(2): 385-392. |
| [35] |
杨文斌, 高顺峰, 万锐, 等. 两种沉水植物对上覆水和间隙水中各形态磷的影响[J]. 环境科学, 2018, 39(5): 2145-2153. Yang W B, Gao S F, Wan R, et al. Effects of submerged macrophytes on different phosphorus fractions in overlying water and interstitial water[J]. Environmental Science, 2018, 39(5): 2145-2153. |
| [36] |
黄子贤, 张饮江, 马海峰, 等. 4种沉水植物对富营养化水体氮磷的去除能力[J]. 生态科学, 2011, 30(2): 102-106. Huang Z X, Zhang Y J, Ma H F, et al. A comparative study on removal efficiency of four submerged plants on phosphorus and nitrogen in eutrophic water under different planting density[J]. Ecological Science, 2011, 30(2): 102-106. DOI:10.3969/j.issn.1008-8873.2011.02.02 |
| [37] |
金树权, 周金波, 包薇红, 等. 5种沉水植物的氮、磷吸收和水质净化能力比较[J]. 环境科学, 2017, 38(1): 156-161. Jin S Q, Zhou J B, Bao W H, et al. Comparison of nitrogen and phosphorus uptake and water purification ability of five submerged macrophytes[J]. Environmental Science, 2017, 38(1): 156-161. |
| [38] |
韩红娟, 翟水晶, 胡维平. 马来眼子菜腐烂分解氮磷转化模型研究[J]. 环境科学, 2010, 31(6): 1483-1488. Han H J, Zhai S J, Hu W P. Modelling nitrogen and phosphorus transfer in Potamogeton malaianus Miq. decompostion[J]. Environmental Science, 2010, 31(6): 1483-1488. |