 2020, Vol. 41
2020, Vol. 41

2. 上海海洋大学水产种质资源发掘与利用教育部重点实验室, 上海 201306;
3. 上海海洋大学水产科学国家级实验教学示范中心, 上海 201306
 , HAN Zi-yu1,2,3 , WANG Meng-xue1,2,3 , MA Yi-chi1,2,3 , WANG Ting1,2,3 , WANG Li-qing1,2,3 , ZHANG Wei1,2,3
, HAN Zi-yu1,2,3 , WANG Meng-xue1,2,3 , MA Yi-chi1,2,3 , WANG Ting1,2,3 , WANG Li-qing1,2,3 , ZHANG Wei1,2,3
2. Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306, China;
3. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China
近年来, 由于医药领域和养殖业对抗生素的大量使用, 使得该类物质在地表水体中呈逐年增加的趋势[1, 2].已有研究表明, 抗生素进入生物体后, 约70%不能被吸收, 而是以母体的形式直接排出体外;尽管抗生素在水体可自然降解, 但速率较慢, 外源性的不断排放, 使其成为水环境中的一种“持久性”污染物[3].水体中抗生素的积累会对水生生物群落结构及其稳定性产生严重影响, 破坏水生态平衡, 并且还可通过食物链的传递, 影响高级生物, 甚至人类健康[4].目前, 使用量较多的抗生素主要有四环素类、磺胺类、大环内酯类以及喹诺酮类等[5, 6].恩诺沙星是用量较大的喹诺酮类抗生素之一, 是畜禽和水产养殖中常用的抗菌药物[7], 已成为地表水环境中最常检测出的新型污染物之一[8, 9].恩诺沙星具有一定的亲脂性, 因而易造成生物体内累积, 可损害生物体肠道健康, 还可导致幼体动物畸形, 其代谢产物环丙沙星还可引起皮疹等疾病[10~12].因此, 对恩诺沙星的生态毒性分析和去除研究, 对于中国地表水环境健康评估和水生态保护非常重要.
淡水微藻是水生态系统中的重要初级生产者, 在水质调控和维持水生态系统稳定性方面具有重要的作用[13].已有研究表明, 排放至水体中的农药、工业染料、重金属和某些抗生素等能够对藻类的生理生化过程产生重要影响[14~17], 如金霉素、氯霉素和氟苯尼考会显著影响铜绿微囊藻(Microcystis aeruginosa)、蛋白核小球藻(Chlorella pyrenoidesa)和斜生栅藻(Scenedesmus obliquus)的生长、生物膜通透性及细胞色素的积累等[13, 18~20].但同时也有研究发现, 微藻对多种污染物具有显著的降解能力[21], 如小球藻(Chlorella sp.)和四尾栅藻(Scenedesmus quadricauda)等可以通过脱羧化和去甲基化的方式进行生物转化或生物催化引发环丙沙星和左氧氟沙星等抗生素的降解[22~26].对于降解机制, 有学者认为微藻细胞在光照条件下, 可以产生具有氧化性的羟基离子和超氧阴离子, 通过氧化或生物吸附的方式促进抗生素等污染物的降解[27, 28];也有学者认为是藻细胞的过氧化氢酶、超氧化物歧化酶及P-450同工酶等引起藻细胞的抗氧化反应, 催化降解污染物[29].但由于藻类物种繁多、不同微藻对于不同抗生素类污染物的适应性和处理效果也不尽相同, 因此筛选出适合不同抗生素污染物生长的微藻种类对于应用藻类资源化处理污染水体具有重要的意义.
目前, 虽已有学者对小球藻和栅藻等常见微藻与环丙沙星、诺氟沙星和恩诺沙星等抗生素的毒理作用进行了报道[25, 26], 但有关其他微藻对恩诺沙星的毒性响应及去除作用的研究较少.本文以绿藻门胶网藻属的纯种藻株胶网藻[30](Dictyosphaerium sp., 藻株编号:FACHB-1902)为实验材料, 通过添加不同浓度恩沙星进行室内培养实验, 定时取样测定胶网藻生物量、细胞色素积累量、光合作用活力以及其水溶性多糖(released polysaccharides, RPS)和胞外胶鞘多糖(capsular polysaccharides, CPS)的变化, 并对恩诺沙星的浓度去除效率进行检测, 探究胶网藻对水体中恩诺沙星的毒性响应和降解能力, 以期为我国地表水中抗生素污染的生态风险评价以及淡水微藻对水体新型污染物的生态去除作用提供新思路.
1 材料与方法 1.1 实验材料恩诺沙星(enrofloxacin, ENR, C19 H22 FN3 O3, 359.40), 购于生工生物工程(上海)股份有限公司.ENR在25℃条件下微溶于水, 易溶于氢氧化钠溶液和氯仿等溶剂, 实验使用时先精确称量1 g恩诺沙星, 溶于10 mL 0.1 mol·L-1的氢氧化钠溶液中, 待其完全溶解后, 用去离子水稀释定容至100 mL, 配置成10 000 mg·L-1的恩诺沙星标准母液用于实验.
1.2 藻种培养胶网藻(Dictyosphaerium sp., FACHB-1902)藻株购于中国科学院水生生物研究所(中国, 武汉), 以BG-11培养基进行培养, 培养条件为:25℃、光照度3 000 lx、光周期12 h:12 h;培养至对数生长期待用.
1.3 实验设计取对数生长期的藻液, 于50 mL无菌离心管中, 用Cetrifuge 5804R型离心机以6000 g离心力离心8 min, 移去上清液, 收集藻细胞, 用BG-11培养基重新悬浮藻细胞, 获得浓度较高的藻液(母液)待用, 用母液绘制梯度生物量及吸光度的标准曲线, 以确定每组中所添加的生物量相同.分别向BG-11培养基中加入初始生物量相同的胶网藻[(12.5±0.1) mg·L-1, 以干重计], 加入一系列梯度的恩诺沙星(0、5、25、50和100 mg·L-1), 每组设3个平行, 培养体积共240 mL, 置于恒温培养箱中培养, 每天定时摇动以使细胞均匀生长, 在0、2、4、6、8、10和12 d时分别取样测定微藻比生长率、叶绿素a和b浓度、类胡萝卜素、叶绿素荧光参数、胞外多糖(exopolysaccharides, EPS)含量和恩诺沙星浓度变化等指标, 另设置无藻组为对照, 以分析胶网藻对恩诺沙星的毒性响应及去除作用.
1.4 指标参数测定方法 1.4.1 胶网藻生物量及比生长率测定取一系列梯度体积的藻细胞母液稀释至15 mL, 取3 mL用UV-2800型分光光度计在680 nm波长下测定其对应吸光度, Whatman GF/F(孔径0.7 μm)玻璃纤维滤膜于0.01 mol·L-1稀盐酸溶液中浸泡10 min后, 去离子水冲洗3次, 以去除滤膜表面的杂质, 减小使用时的称量误差.洗涤干净后于105℃条件下烘干, 分别取10 mL上述梯度的藻液抽滤, 并编号, 对折后于相同条件下烘干, 再次称重, 计算微藻生物量, 以吸光度为横坐标, 干重生物量为纵坐标绘制胶网藻标准曲线为: y=0.826 2x-0.003 3, R2=0.998 9.实验时每次取摇匀后的藻液3 mL于680 nm波长下测定其吸光度, 根据标准曲线计算取样当日胶网藻的干重生物量, 计算比生长率(μ), 公式如下:
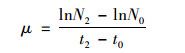
|
式中, N2表示测定当天(t2)的干重生物量(mg·L-1), N0表示第0天(t0)的初始生物量;t表示天数(d).
1.4.2 叶绿素和类胡萝卜素的测定取5 mL藻液6000g离心15 min, 移去上清液, 然后加入10 mL 90%乙醇;混合均匀后, 于60℃恒温水浴锅热浴10 min, 再于6000 g离心15 min;分光光度计测上清液在波长为665、652和470 nm下的吸光度.叶绿素a、叶绿素b和类胡萝卜素的计算公式如下[14, 31]:
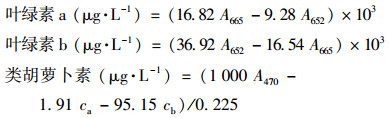
|
式中, ca表示叶绿素a的浓度(mg·L-1);cb表示叶绿素b的浓度(mg·L-1).
1.4.3 叶绿素荧光参数的测定取3 mL摇匀后的藻液于测量杯中, 暗适应5 min后, 用浮游植物荧光分析仪(Phyto-PAM, Walz, Germany)测定光合系统Ⅱ的(PSⅡ)最大光合速率(Fv/Fm)、实际光合效率(Yield)、快速光曲线的初始斜率(α)和最大相对电子传递速率(ETRmax).其中Fv/Fm与光化学量子产额成正比, 运用Fv/Fm=(Fm-F0)/Fm计算, 式中, Fv为最大可变荧光发射, Fm为最大荧光, F0为暗适应状态下的打开饱和脉冲前的最大荧光;当处于暗适应状态时Yield值即为Fv/Fm;α表示初始斜率(胶网藻对光能的利用效率);具体参数指标测定及分析参照文献[32, 33].
1.4.4 胞外多糖(EPS)含量测定胞外多糖的测定包括两部分, 藻类分泌到培养液中的水溶性多糖(RPS)及包裹在藻细胞外的胶鞘多糖(CPS), 提取及测定方法参照文献[34, 35].首先用标准蔗糖溶液绘制标准曲线, 吸光度为横坐标, 对应糖含量为横坐标, 求出线性回归方程:
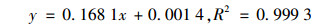
|
取3 mL藻液于离心管中, 10 000 g离心15 min收集含RPS的上清液, 将上清液转移至另一干净的离心管, 加入0.5 mL蒽酮乙酸乙酯试剂和5 mL浓硫酸, 充分振荡, 立即将试管放入沸水浴中, 准确保温1 min.取出后自然冷却至室温, 以空白作参比, 在630 nm波长下测其吸光度;CPS从上述离心所得沉淀的藻细胞中提取, 用3 mL 0.05%的NaCl溶液重悬藻细胞, 60℃水浴30 min, 随后10 000 g离心15 min收集含CPS的上清液, 将上清液全部转移至另一干净的离心管中, 其余步骤同RPS, 由标准线性方程求出糖的量(mg).根据以下公式计算每克干重藻细胞中多糖量(CPS和RPS):
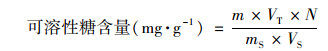
|
式中, m表示从标准曲线查得的糖量(mg);VT表示提取液总体积(mL);VS表示测定时取用的样品体积(mL);N表示稀释倍数;mS表示样品质量(g).
1.4.5 恩诺沙星浓度的测定利用高效液相色谱仪(HPLC, Agilent 1100, USA)测定恩诺沙星浓度[10].经色谱柱:Agilent Zorbax XDB-C18分析柱;流动相:甲醇:(0.05 mol·L-1柠檬酸、0.01 mol·L-1醋酸铵混合溶液, 柠檬酸铵)=25:75(体积比);激发波长:280 nm, 发射波长:450 nm;流速1.5 mL·min-1;柱温40℃;进样量为10 μL.首先, 配置不同浓度恩诺沙星(0.1、0.25、0.5、1、2.5和5 mg·L-1)标准样品, 绘制标准曲线, 以峰面积为横坐标, 恩诺沙星浓度为纵坐标, 拟合曲线方程为y=0.003 3x+0.044 8, R2=0.999 9.实验样品取样测定时先将藻液摇匀, 取1 mL, 用0.22 μm无菌滤头过滤至进样瓶中, 避光保存于-20℃冰箱, 于高效液相色谱仪测定其峰面积, 根据标准曲线计算恩诺沙星浓度.
1.5 数据处理与分析利用单因素方差分析(ANOVA)比较不同浓度恩诺沙星处理条件下各组间的微藻生物量、胞外多糖(RPS和CPS)、叶绿素a、叶绿素b、类胡萝卜素、叶绿素荧光参数和恩诺沙星浓度的差异性, 首先进行方差齐性检验, 符合正态分布, P < 0.05即为处理组和对照组间具有显著差异性, P < 0.01即认为处理组和对照组间在双侧水平上具有极显著的差异性, 所有统计分析均在SPSS 17.0中完成, 运用Origin 9.0进行图表的绘制.
2 结果与分析 2.1 胶网藻生物量变化各处理组胶网藻生物量在12 d内均逐渐增加;但随着恩诺沙星浓度的升高, 胶网藻生物量的增加率降低(图 1).对照组由初始的(12.5±0.01) mg·L-1升高至(108.64±6.83) mg·L-1, 比生长率为0.203, 而当恩诺沙星浓度为100 mg·L-1时, 生物量由(12.5±0.01) mg·L-1升高至(64.89±0.70) mg·L-1, 比生长率为0.154, 显著低于对照组(P < 0.01).此外, 与对照组相比, 实验开始的前2 d, 各处理组组间未表现出显著差异性, 但从第4 d开始, 各组均显著(P < 0.05)或极显著(P < 0.01)低于对照组, 且恩诺沙星浓度越高, 抑制作用越强[图 1(a)].通过生物量和恩诺沙星浓度变化拟合计算恩诺沙星对胶网藻96 h半致死浓度(LC50)为(241.29±7.33) mg·L-1.不同浓度恩诺沙星处理条件下的比生长率在第2 d时各组间无显著性差异, 而在4~12 d内, 除第6 d时的25 mg·L-1处理组及12 d时的5 mg·L-1处理组与对照组有显著差异(P < 0.05), 其余组均与对照组有极显著差异性(P < 0.01), 实验结束时(第12 d)的比生长率大小为对照组>5 mg·L-1>25 mg·L-1>50 mg·L-1>100 mg·L-1, 分别为0.203、0.190、0.185、0.168和0.154 [图 1(b)].

|
*表示双侧水平上具有显著性差异(P < 0.05), **表示双侧水平上具有极显著性差异(P < 0.01), 下同 图 1 不同浓度恩诺沙星胁迫下胶网藻干重和比生长率变化 Fig. 1 Effects of enrofloxacin concentrations on microalgal growth in terms of dry cell weight and specific growth rate of Dictyosphaerium sp. |
在实验周期内, 各处理组的叶绿素[图 2(a)、2(b)]和类胡萝卜素[图 2(c)]含量均逐渐增加, 但增长率随恩诺沙星浓度的增加逐渐降低.在第2 d时, 仅有高浓度组(100 mg·L-1)的叶绿素a和对照组有显著的差异性(P < 0.05), 而在4~12 d内, 高于5 mg·L-1的其余3组叶绿素a含量均极显著低于对照组(P < 0.01), 叶绿素b和类胡萝卜素的变化趋势类似于叶绿素a.此外, 在第12 d时, 对照组的叶绿素a、叶绿素b和类胡萝卜素积累量分别为2 051.61、414.57和677.27 μg·L-1, 是0 d时的20.3、16.3和23倍, 而100 mg·L-1恩诺沙星胁迫条件下三者的积累量分别为982.67、284.45和343.59 μg·L-1, 是实验初期的9.7、11.1和11.7倍, 极显著低于对照组(P < 0.01), 添加不同浓度恩诺沙星对于胶网藻藻细胞色素积累有不同程度的抑制作用.

|
图 2 不同浓度恩诺沙星胁迫下胶网藻叶绿素a、叶绿素b和类胡萝卜素含量的变化 Fig. 2 Effects of enrofloxacin concentrations on the chlorophyll a (Chl-a), chlorophyll b (Chl-b), and carotenoid levels of Dictyosphaerium sp. |
胶网藻光合系统Ⅱ(PSⅡ)的最大光合效率(Fv/Fm)在12 d内整体呈现先升高后降低的变化趋势(图 3).第4 d时升至最高, 5 mg·L-1处理组平均值达到0.75, 并且由图 3(a)可以看出, 5 mg·L-1可以促进Fv/Fm的增加, 使其测定值高于对照组和其他处理组.在第2 d时仅有高浓度组与对照组有显著差异性, 其余组无显著差异, 在6~10 d内, 50 mg·L-1和100 mg·L-1组的Fv/Fm均显著低于对照组, 最后一天各组的Fv/Fm与对照组未表现出差异性.实际光合速率(Yeild)在12 d内呈现先降低后有所升高的变化趋势[图 3(b)], 与Fv/Fm的变化有所不同, 其测定值在第6 d时最低为0.09, 随后有所回升.除第8 d外, 快速光曲线的斜率α均随着恩诺沙星浓度的增加逐渐降低的变化趋势, 并且50 mg·L-1和100 mg·L-1处理组的α均显著(P < 0.05)或极显著(P < 0.01)低于对照组, 12 d时100 mg·L-1处理组的斜率α最低为0.14, 是对照组的0.77倍[图 3(c)].最大电子传递速率(ETRmax)在12 d内的变化趋势与Yeild相似, 第6 d降至最低后逐渐回升, 但回升率较低, 第12 d时各组的ETRmax分别是第0 d时的0.32、0.39、0.43、0.31和0.27倍[图 3(d)].

|
图 3 不同浓度恩诺沙星胁迫下胶网藻最大光合效率(Fv/Fm)、实际光合效率(Yeild)、初始斜率(α)和最大相对电子传递速率(ETRmax)的变化趋势 Fig. 3 Effects of different concentrations of enrofloxacin on the maximum photosynthetic efficiency (Fv/Fm), the potential photosynthetic activity of PSⅡ(Yield), the initial slope (α), and the maximum relative electronic rate (ETRmax) of Dictyosphaerium sp. |
在不同浓度恩诺沙星胁迫条件下, 胶网藻细胞的胶鞘多糖(CPS)呈现逐渐下降的趋势, 而释放到周围环境中的水溶性多糖(RPS)却表现为先升后降的趋势[图 4(a)和4(b)].在2~10 d内, 恩诺沙星胁迫浓度为50 mg·L-1时的CPS测定值均高于其余几组, 并且在6 d时与对照组有极显著差异性(P < 0.01), 第12 d时的CPS有所回升, 但各组与对照组无显著性差异.对于RPS而言, 第8 d时的测定值升至最高, 且5 mg·L-1处理组与对照组无显著差异, 25 mg·L-1处理组与对照组有显著差异性(P < 0.05), 另外两组与对照组存在极显著性差异(P < 0.01), 恩诺沙星浓度为100 mg·L-1条件下, 1 g干重的胶网藻RPS升高至251.67 mg·g-1 [图 4(b)].RPS的初始含量是CPS的5.78倍, 释放到培养液中的多糖含量远高于包裹在细胞外的胶鞘多糖含量.

|
图 4 不同浓度恩诺沙星胁迫下胶网藻胞外胶鞘多糖(CPS)和水溶性多糖(RPS)含量变化 Fig. 4 Effects of enrofloxacin concentrations on the capsular polysaccharides (CPS) and released polysaccharides (RPS) of Dictyosphaerium sp. |
通过比较各组对恩诺沙星的去除量可发现, 除了在第2 d时, 25 mg·L-1恩诺沙星浓度条件下, 有藻组和无藻组(对照组)之间无显著差异外, 添加胶网藻的处理组对于恩诺沙星的去除量总体显著高于各对照组(P < 0.01)(图 5).在第2 d时, 添加胶网藻对5 mg·L-1的去除率达到9.6%, 高于其它几组的去除率[图 5(a)], 但随培养天数的增加, 各组的去除速率逐渐加快.对于只添加BG-11培养基的条件下, 4个梯度(5、25、50和100 mg·L-1)在12 d内分别降至4.64、23.61、47.35和95.12 mg·L-1, 去除率分别为7.27%、5.56%、5.30%和4.88%.而添加胶网藻的4组在12 d内分别降至3.83、20.82、42.87和86.20 mg·L-1, 去除率分别为23.34%、16.71%、14.26%和13.80%, 是无藻组的2.69~3.21倍.

|
图 5 不同浓度的恩诺沙星在有藻和无藻条件下的去除量比较 Fig. 5 Removal of enrofloxacin at different concentrations under the Dictyosphaerium sp. and no algae |
自然水体中抗生素含量一般为ng·L-1级别, 虽然其含量较低, 但在水体中留存时间长、并且由于有持续性的外源增补, 长期残留, 会对水生生物产生愈发严重的毒害作用, 因此有必要设置一定的浓度梯度, 探究高于水体含量的抗生素浓度对于水生生物所造成的影响, 来预估抗生素对水生生物的潜在危害[36].单细胞藻类对抗生素的应激反应主要表现为生长、细胞形态变化及生理生化等各个方面[37, 38], 具体是通过影响藻类细胞成分、改变细胞膜的通透性、影响基因表达、阻碍光合作用进程、诱导产生活性氧自由基引发氧化胁迫及破坏抗氧化酶系统等方式[39].目前, 关于微藻和恩诺沙星相互作用的研究较少, 仅有Xiong等[40]研究了1~100 mg·L-1的恩诺沙星对斜生栅藻、墨西哥衣藻(Chlamydomonas mexicana)、普通小球藻和微茫藻(Micractinium resseri)等4种绿藻对恩诺沙星之间的响应关系, 该研究发现, 当恩诺沙星浓度≥1 mg·L-1时, 对几种微藻的生长及色素积累(光合作用)均有显著地抑制作用, 但这4种绿藻的形态特征及生长特性与胶网藻有所不同, 不能反映胶网藻对恩诺沙星胁迫的应激能力及对恩诺沙星的去除能力.因而本研究在此基础上设置了0、5、25、50和100 mg·L-1进行实验探究.结果表明5~100 mg·L-1浓度的恩诺沙星胁迫均会对胶网藻的生长、光合作用及胞外多糖的含量产生影响(图 1~4), 与Andrieu等[41]和Ebert等[42]的研究结果一致.虽然恩诺沙星对胶网藻的96 h半数致死浓度(LC50)高达241.29 mg·L-1, 但恩诺沙星为5 mg·L-1时即可对胶网藻产生显著的抑制作用, 且浓度越高, 抑制作用越强;单就96 h的LC50浓度而言, 胶网藻对该抗生素的耐受力高于小球藻和四尾栅藻等其它藻类[13].另有研究发现, 抗生素等污染物能够影响微藻细胞膜的通透性, 使其分泌的多糖和蛋白质等物质增加[39], 在本研究中发现, 胶网藻在受到恩诺沙星刺激时, 其分泌到培养液中的RPS含量在短期(8 d)内逐渐升高, 随后有所下降, 而CPS积累量呈现逐渐降低的趋势(图 4), 可能是由于恩诺沙星致使藻细胞膜受损, 细胞黏附性降低[43]导致溶液中的RPS短期升高而CPS快速下降.此外, 不同浓度恩诺沙星对胶网藻的光合作用过程也产生了一定的影响(图 3), 首先表现为叶绿素的积累速率随恩诺沙星浓度的升高而降低, 针对此项结果, 已有研究表明光合色素的降低是微藻的一种常见应激反应, 是由于类囊体脂质过氧化和PSⅡ复合物的降解所引起的[44].另外, 在低浓度(5 mg·L-1)恩诺沙星胁迫条件下, 可以在一定程度上促进胶网藻的光合系统Ⅱ(PSⅡ)最大光合效率(Fv/Fm), 这主要是由于类胡萝卜素的抗氧化特性发挥了重要作用, 研究表明低浓度恩诺沙星的暴露可使类胡萝卜素通过抑制单线态氧和细胞内产生的自由基来保护细胞, 防止脂质过氧化, 促进光合器官的稳定性和功能, 使光合作用增强[45].另外, 类胡萝卜素还可以促进细胞膜的完整性, 这对细胞存活至关重要[40], 本研究中类胡萝卜素的积累量逐天增加, 正是在对恩诺沙星胁迫所做出的应激反应.但高于5 mg·L-1后, 对几个光合参数均有一定的抑制作用, 这主要是大量进入藻细胞的抗生素可以引发一系列氧化还原反应, 产生大量自由基, 阻碍叶绿体上的电子传递过程, 导致膜脂过氧化, 抑制PSⅠ和PSⅡ的光合活性;另外也可以直接抑制叶绿体蛋白质的表达或降低光合作用电子传递链上递氢电子体的表达, 从而抑制光合作用过程[46~48].
前期已有研究发现, 微藻具有促进水体中抗生素等污染物的去除潜力[26, 49].Xiong等[40]的研究也表明多种微藻协同作用比单种藻对抗生素的去除率更高, 小球藻、墨西哥衣藻、斜生栅藻和微芒藻等对恩诺沙星的去除率最高分别可达到26%、25%、23%和18%, 主要为微藻的生物积累、生物吸附和酶系统催化等方式去除或降解抗生素.不过, 另有研究认为水解和光解通常也是污染物降解的主要途径[50].在本研究中, 添加胶网藻的组中5 mg·L-1恩诺沙星的去除率在12 d时即可达到23.34%, 是相同浓度下对照组(无藻组)的3.21倍, 低于前期所研究的墨西哥衣藻和普通小球藻, 但高于微芒藻和斜生栅藻等[40].胶网藻的存在能够促进恩诺沙星的去除, 并且在恩诺沙星浓度低于50 mg·L-1的水平内, 去除率随胁迫浓度的降低而升高.而各对照组由自然光解或水解对恩诺沙星的去除率仅为4.88% ~7.27%, 处于较低水平.除生物积累和酶系统催化降解外, 微藻在受胁迫的情况下可产生大量的活性氧自由基, 可氧化降解抗生素等污染物, 同时, 微藻也具有内源性分解代谢系统以及污染物转化能力[25].此外, 藻类胞外聚合物(EPS)主要成分为蛋白质和多糖, 不仅可以起到保护细胞, 使其凝聚为团体以抵御捕食等作用, 还可以通过结合成糖蛋白的形式捕捉水体中的污染物从而达到吸附去除的目的[43];恩诺沙星的暴露可刺激藻细胞不断分泌多糖类、活性氧等物质, 以降解污染物[39];同时, 微藻也可通过细胞壁所富有的蛋白质和多糖类物质结合形成的糖蛋白吸附污染物, 以产生去除水体抗生素、重金属等物质的效果[51].另外, EPS成分中也含有氨基酸、腐殖酸等多种氧化性物质[34], 它们可参与水体光降解过程, 起到促进有机污染物产生光降解的作用[14, 52, 53], 不过在本研究中, EPS在胶网藻对恩诺沙星的毒性响应以及在胶网藻对恩诺沙星的降解过程中具体如何发挥作用, 其机制还需要进一步深入研究.
4 结论(1) 恩诺沙星对胶网藻的生长及色素积累均有显著抑制作用(P < 0.05), 并且恩诺沙星可刺激胶网藻释放EPS.
(2) 当恩诺沙星浓度为5 mg·L-1时, 能够促进胶网藻的光合作用, 高于5 mg·L-1则对其具有抑制作用;暴露培养6 d后, 胶网藻可逐渐恢复光合作用活力.
(3) 第12 d时, 添加胶网藻的各处理组恩诺沙星去除率分别为对照组(无藻组)的3.21、3.01、2.69和2.83倍, 与自然条件相比, 胶网藻对于水体中恩诺沙星残留物的去除具有显著的促进作用(P < 0.05).
| [1] | Schwarzenbach R P, Escher B I, Fenner K, et al. The challenge of micropollutants in aquatic systems[J]. Science, 2006, 313(5790): 1072-1077. DOI:10.1126/science.1127291 |
| [2] | Barber L B, Keefe S H, Brown G K, et al. Persistence and potential effects of complex organic contaminant mixtures in wastewater-impacted streams[J]. Environmental Science & Technology, 2013, 47(5): 2177-2188. |
| [3] |
杨弯弯, 武秋贤, 吴亦潇, 等. 恩诺沙星和硫氰酸红霉素对铜绿微囊藻的毒性研究[J]. 中国环境科学, 2013, 33(10): 1829-1834. Yang W W, Wu Q X, Wu Y X, et al. Toxicity of enrofloxacin and erythromycin thiocyanate on Microcystis aeruginosa[J]. China Environmental Science, 2013, 33(10): 1829-1834. |
| [4] | Gonzãlez-Pleiter M, Gonzalo S, Rodea-Palomares I, et al. Toxicity of five antibiotics and their mixtures towards photosynthetic aquatic organisms:implications for environmental risk assessment[J]. Water Research, 2013, 47(6): 2050-2064. DOI:10.1016/j.watres.2013.01.020 |
| [5] |
姜蕾, 陈书怡, 杨蓉, 等. 长江三角洲地区典型废水中抗生素的初步分析[J]. 环境化学, 2008, 27(3): 371-374. Jiang L, Chen S Y, Yang R, et al. Occurrence of antibiotics in the aquatic environment of the Changjiang Delta, China[J]. Environmental Chemistry, 2008, 27(3): 371-374. DOI:10.3321/j.issn:0254-6108.2008.03.022 |
| [6] | Yi R H, Wang Q J, Mo C H, et al. Determination of four fluoroquinolone antibiotics in tap water in Guangzhou and Macao[J]. Environmental Pollution, 2010, 158(7): 2350-2358. DOI:10.1016/j.envpol.2010.03.019 |
| [7] |
朱婷婷, 宋战锋, 尹魁浩, 等. 南方某水库水体中抗生素生态与健康风险研究[J]. 生态毒理学报, 2015, 10(5): 124-131. Zhu T T, Song Z F, Yin K H, et al. Assessments of ecological and health risk induced by antibiotics in source water of a reservoir in a Southern City[J]. Asian Journal of Ecotoxicology, 2015, 10(5): 124-131. |
| [8] | Lin A Y C, Tsai Y T. Occurrence of pharmaceuticals in Taiwan's surface waters:impact of waste streams from hospitals and pharmaceutical production facilities[J]. Science of the Total Environment, 2009, 407(12): 3793-3802. DOI:10.1016/j.scitotenv.2009.03.009 |
| [9] | Liu J Z, Danneels B, Vanormelingen P, et al. Nutrient removal from horticultural wastewater by benthic filamentous algae Klebsormidium sp., Stigeoclonium spp. and their communities:from laboratory flask to outdoor algal turf scrubber (ATS)[J]. Water Research, 2016, 92: 61-68. DOI:10.1016/j.watres.2016.01.049 |
| [10] |
朱凤娇.恩诺沙星在中华绒螯蟹模拟养殖系统中的分布及其耐药性在转录水平的研究[D].上海: 上海海洋大学, 2017. Zhu F J. Distribution of enrofloxacin in simulated aquaculture system of Chinese mitten crab(Eriocheir sinensis) and study it resistance at the transcription level[D]. Shanghai: Shanghai Ocean University, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10264-1017854314.htm |
| [11] |
宋红波, 吴光红, 沈美芳, 等. 恩诺沙星在水产品中残留的风险评估[J]. 渔业现代化, 2008, 35(5): 39-42. Song H B, Wu G H, Shen M F, et al. Risk evaluation of enrofloxacin in aquatic products[J]. Fishery Modernization, 2008, 35(5): 39-42. DOI:10.3969/j.issn.1007-9580.2008.05.010 |
| [12] | Vancutsem P M, Babish J G, Schwark W S. The fluoroquinolone antimicrobials:structure, antimicrobial activity, pharmacokinetics, clinical use in domestic animals and toxicity[J]. Cornell Veterinarian, 1990, 80(2): 173-186. |
| [13] |
徐冬梅, 王艳花, 饶桂维. 四环素类抗生素对淡水绿藻的毒性作用[J]. 环境科学, 2013, 34(9): 3386-3390. Xu D M, Wang Y H, Rao G W. Cellular response of freshwater green algae to the toxicity of tetracycline antibiotics[J]. Environmental Science, 2013, 34(9): 3386-3390. |
| [14] | Xiong J Q, Kurade M B, Kim J R, et al. Ciprofloxacin toxicity and its co-metabolic removal by a freshwater microalga Chlamydomonas mexicana[J]. Journal of Hazardous Materials, 2017, 323: 212-219. DOI:10.1016/j.jhazmat.2016.04.073 |
| [15] | Sathasivam R, Ebenezer V, Guo R Y, et al. Physiological and biochemical responses of the freshwater green algae Closterium ehrenbergii to the common disinfectant chlorine[J]. Ecotoxicology and Environmental Safety, 2016, 133: 501-508. DOI:10.1016/j.ecoenv.2016.08.004 |
| [16] |
孔玄庆, 郭军, 金晨钟, 等. 农药与藻类互作的研究进展[J]. 现代农业科技, 2016(5): 222-224, 228. Kong X Q, Guo J, Jin C Z, et al. Research progress on interaction between pesticides and algae[J]. Modern Agricultural Science and Technology, 2016(5): 222-224, 228. DOI:10.3969/j.issn.1007-5739.2016.05.129 |
| [17] | Qin H W, Chen L F, Lu N, et al. Toxic effects of enrofloxacin on Scenedesmus obliquus[J]. Frontiers of Environmental Science & Engineering, 2012, 6(1): 107-116. |
| [18] | Yao L L, Wang Y X, Tong L, et al. Occurrence and risk assessment of antibiotics in surface water and groundwater from different depths of aquifers:a case study at Jianghan Plain, central China[J]. Ecotoxicology and Environmental Safety, 2017, 135: 236-242. DOI:10.1016/j.ecoenv.2016.10.006 |
| [19] | Guo R X, Chen J Q. Phytoplankton toxicity of the antibiotic chlortetracycline and its UV light degradation products[J]. Chemosphere, 2012, 87(11): 1254-1259. DOI:10.1016/j.chemosphere.2012.01.031 |
| [20] | Lai H T, Hou J H, Su C I, et al. Effects of chloramphenicol, florfenicol, and thiamphenicol on growth of algae Chlorella pyrenoidosa, Isochrysis galbana, and Tetraselmis chui[J]. Ecotoxicology and Environmental Safety, 2009, 72(2): 329-334. DOI:10.1016/j.ecoenv.2008.03.005 |
| [21] | Xiong J Q, Kurade M B, Jeon B H, et al. Biodegradation of levofloxacin by an acclimated freshwater microalga, Chlorella vulgaris[J]. Chemical Engineering Journal, 2017, 313: 1251-1257. DOI:10.1016/j.cej.2016.11.017 |
| [22] | Matamoros V, Uggetti E, García J, et al. Assessment of the mechanisms involved in the removal of emerging contaminants by microalgae from wastewater:a laboratory scale study[J]. Journal of Hazardous Materials, 2015, 301: 197-205. |
| [23] |
杜迎翔, 冯云庆, 项钟润, 等. 蛋白核小球藻去除2种头孢类抗生素的研究[J]. 环境科学与技术, 2015, 38(10): 105-111. Du Y X, Feng Y Q, Xiang Z R, et al. Removal of two cephalosporins in Chlorella pyrenoidosa[J]. Environmental Science & Technology, 2015, 38(10): 105-111. |
| [24] | Xiong J Q, Kurade M B, Patil D V, et al. Biodegradation and metabolic fate of levofloxacin via a freshwater green alga, Scenedesmus obliquus in synthetic saline wastewater[J]. Algal Research, 2017, 25: 54-61. DOI:10.1016/j.algal.2017.04.012 |
| [25] |
朱小燕.淡水藻引发喹诺酮类抗生素的降解行为研究[D].南京: 东南大学, 2012. Zhu X Y. Study on degradation behavior of quinolone antibacterial induced by freshwater algae[D]. Nanjing: Southeast University, 2012. |
| [26] |
朱小燕, 傅大放, 邓琳, 等. 小球藻引发水中环丙沙星的光降解效能研究[J]. 中国环境科学, 2013, 33(4): 663-668. Zhu X Y, Fu D F, Deng L, et al. Efficiency of ciprofloxacin photodegradation induced by chlorella in aqueous solutions[J]. China Environmental Science, 2013, 33(4): 663-668. DOI:10.3969/j.issn.1000-6923.2013.04.012 |
| [27] |
姜晶, 李亮, 李海鹏, 等. 蛋白核小球藻对Pb(Ⅱ)和Cd(Ⅱ)的生物吸附及其影响因素[J]. 生态学报, 2012, 32(7): 1995-2003. Jiang J, Li L, Li H P, et al. Biosorption of lead (Ⅱ) and cadmium (Ⅱ) from aqueous solution by Chlorella pyrenoidsa and its influential factors[J]. Acta Ecologica Sinica, 2012, 32(7): 1995-2003. |
| [28] | Peng Z E, Wu F, Deng N S. Photodegradation of bisphenol A in simulated lake water containing algae, humic acid and ferric ions[J]. Environmental Pollution, 2006, 144(3): 840-846. DOI:10.1016/j.envpol.2006.02.006 |
| [29] | Gao Q T, Tam N F Y. Growth, photosynthesis and antioxidant responses of two microalgal species, Chlorella vulgaris and Selenastrum capricornutum, to nonylphenol stress[J]. Chemosphere, 2011, 82(3): 346-354. DOI:10.1016/j.chemosphere.2010.10.010 |
| [30] |
王秀海, 赵震宇, 刘平怀, 等. 胶网藻化学成分及其体外抗氧化、抗菌活性研究[J]. 食品工业, 2018, 39(7): 206-211. Wang X H, Zhao Z Y, Liu P H, et al. The chemical components of Dictyosphaerium sp. and their antioxidant, antibacterial activities in vitro[J]. The Food Industry, 2018, 39(7): 206-211. |
| [31] | Xiong J Q, Kurade M B, Abou-Shanab R A I, et al. Biodegradation of carbamazepine using freshwater microalgae Chlamydomonas mexicana and Scenedesmus obliquus and the determination of its metabolic fate[J]. Bioresource Technology, 2016, 205: 183-190. DOI:10.1016/j.biortech.2016.01.038 |
| [32] | Roháček K, Barták M. Technique of the modulated chlorophyll fluorescence:basic concepts, useful parameters, and some applications[J]. Photosynthetica, 1999, 37(3): 339-363. |
| [33] | Zhang W, Jeppesen E, Wang M M, et al. Allelopathic effect boosts Chrysosporum ovalisporum dominance in summer at the expense of Microcystis panniformis in a shallow coastal water body[J]. Environmental Science and Pollution Research, 2017, 24(5): 4666-4675. DOI:10.1007/s11356-016-8149-0 |
| [34] |
陈毛珍, 谢梅娟, 巫伟, 等. 微囊藻胶鞘多糖和水溶性胞外多糖的化学特性[J]. 湖泊科学, 2016, 28(3): 609-615. Chen M Z, Xie M J, Wu W, et al. Chemical characteristics of capsular polysaccharide and water-soluble released exopolysaccharide from Microcystis[J]. Journal of Lake Sciences, 2016, 28(3): 609-615. |
| [35] | 王学奎. 植物生理生化实验原理和技术[M]. ((第二版)). 北京: 高等教育出版社, 2006. |
| [36] | 管超, 毛杰. 抗生素对水生态环境毒性效应研究概述[J]. 资源节约与环保, 2014(2): 136. |
| [37] | Chen J Q, Guo R X. Access the toxic effect of the antibiotic cefradine and its UV light degradation products on two freshwater algae[J]. Journal of Hazardous Materials, 2012, 209-210: 520-523. DOI:10.1016/j.jhazmat.2012.01.041 |
| [38] | Wen Y Z, Chen H, Shen C S, et al. Enantioselectivity tuning of chiral herbicide dichlorprop by copper:Roles of reactive oxygen species[J]. Environmental Science & Technology, 2011, 45(11): 4778-4784. |
| [39] |
万禁禁.淡水微藻对几种抗生素胁迫的响应研究[D].泉州: 华侨大学, 2014. Wan J J. Study the response of freshwater microalgae to several antibiotics stress[D]. Quanzhou: Huaqiao University, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10385-1014079605.htm |
| [40] | Xiong J Q, Kurade M B, Jeon B H. Ecotoxicological effects of enrofloxacin and its removal by monoculture of microalgal species and their consortium[J]. Environmental Pollution, 2017, 226: 486-493. DOI:10.1016/j.envpol.2017.04.044 |
| [41] | Andrieu M, Rico A, Phu T M, et al. Ecological risk assessment of the antibiotic enrofloxacin applied to Pangasius catfish farms in the Mekong Delta, Vietnam[J]. Chemosphere, 2015, 119: 407-414. DOI:10.1016/j.chemosphere.2014.06.062 |
| [42] | Ebert I, Bachmann J, Kühnen U, et al. Toxicity of the fluoroquinolone antibiotics enrofloxacin and ciprofloxacin to photoautotrophic aquatic organisms[J]. Environmental Toxicology and Chemistry, 2011, 30(12): 2786-2792. DOI:10.1002/etc.678 |
| [43] |
支田田, 程丽华, 徐新华, 等. 藻类去除水体中重金属的机理及应用[J]. 化学进展, 2011, 23(8): 1782-1794. Zhi T T, Cheng L H, Xu X H, et al. Advances on heavy metals removal from aqueous solution by algae[J]. Progress in Chemistry, 2011, 23(8): 1782-1794. |
| [44] | Nie X P, Wang X, Chen J F, et al. Response of the freshwater alga Chlorella vulgaris to trichloroisocyanuric acid and ciprofloxacin[J]. Environmental Toxicology and Chemistry, 2008, 27(1): 168-173. |
| [45] | Jahns P, Holzwarth A R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem Ⅱ[J]. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 2012, 1817(1): 182-193. |
| [46] |
刘滨扬.红霉素、环丙沙星和磺胺甲噁唑对羊角月牙藻的毒性效应及其作用机理[D].广州: 暨南大学, 2011. Liu B Y. Toxic effects and its mechanism of erythromycin, ciprofloxacin and sulfamethoxazole to Selenastrum capricornutum[D]. Guangzhou: Ji'nan University, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10559-1011128731.htm |
| [47] | Pan X L, Deng C N, Zhang D Y, et al. Toxic effects of amoxicillin on the photosystem Ⅱ of Synechocystis sp. characterized by a variety of in vivo chlorophyll fluorescence tests[J]. Aquatic Toxicology, 2008, 89(4): 207-213. DOI:10.1016/j.aquatox.2008.06.018 |
| [48] | Liu W H, Ming Y, Huang Z W, et al. Impacts of florfenicol on marine diatom Skeletonema costatum through photosynthesis inhibition and oxidative damages[J]. Plant Physiology and Biochemistry, 2012, 60: 165-170. DOI:10.1016/j.plaphy.2012.08.009 |
| [49] | Matamoros V, Gutierrez R, Ferrer I, et al. Capability of microalgae-based wastewater treatment systems to remove emerging organic contaminants:a pilot-scale study[J]. Journal of Hazardous Materials, 2015, 288: 34-42. DOI:10.1016/j.jhazmat.2015.02.002 |
| [50] | Andreozzi R, Raffaele M, Nicklas P. Pharmaceuticals in STP effluents and their solar photodegradation in aquatic environment[J]. Chemosphere, 2013, 50(10): 1319-1330. |
| [51] | 王奇, 葛姝洁, 王传花, 等. 水华期间藻类分层胞外聚合物与重金属的相互作用机制研究[J]. 环境保护与循环经济, 2017, 37(3): 35-40. DOI:10.3969/j.issn.1674-1021.2017.03.009 |
| [52] | De Godos I, Muñoz R, Guieysse B. Tetracycline removal during wastewater treatment in high-rate algal ponds[J]. Journal of Hazardous Materials, 2012, 229-230: 446-449. DOI:10.1016/j.jhazmat.2012.05.106 |
| [53] | Zhang J W, Fu D F, Wu J L. Photodegradation of Norfloxacin in aqueous solution containing algae[J]. Journal of Environmental Sciences, 2012, 24(4): 743-749. DOI:10.1016/S1001-0742(11)60814-0 |