 2020, Vol. 41
2020, Vol. 41

2. 中国科学院上海高等研究院, 上海 201210;
3. 上海市农产品质量安全中心, 上海 200335
 , FENG Wen-wen1,2 , DONG Yong-hua3 , MA Zhong-liang1 , CAO Hui-jin1,2 , SUN Jun-song2 , ZHANG Bao-guo2
, FENG Wen-wen1,2 , DONG Yong-hua3 , MA Zhong-liang1 , CAO Hui-jin1,2 , SUN Jun-song2 , ZHANG Bao-guo2
2. Shanghai Advanced Research Institute, Chinese Academy of Sciences, Shanghai 201210, China;
3. Shanghai Agricultural Product Quality and Safety Center, Shanghai 200335, China
我国是畜禽养殖和果树种植大国, 近年来随着规模化、集约化的畜禽养殖数量和果树栽培面积的数量不断增加, 产生了大量的畜禽粪便和果树修剪枝条等农业有机固体废弃物[1, 2].这些农业有机废弃物如果不能及时有效加以处理, 就会成为生态环境污染的重要来源, 不仅造成了农业生态环境质量不断恶化, 危害人们的身心健康, 同时也导致大量附加值高的有用成分和养分资源流失, 造成严重的农业面源污染, 极大地削弱了我国现代农业可持续发展能力[3].因此, 切实有效地实现不同农业有机固体废弃物的减量化处理与资源化利用已经成为目前研究的重点和热点.
目前实现农业废弃物减量化、无害化和资源化的方式之一是进行好氧堆肥处理.好氧堆肥的本质是微生物分解和转化有机物的生化代谢过程, 因此了解堆肥过程中不同时期微生物丰度、多样性以及群落演替规律对于揭示堆肥过程中的有机物降解机制和优化堆肥处理工艺具有重要理论意义和实践价值.常用的分析堆肥过程中微生物相对丰度和微生物群落结构组成的经典技术有克隆测序技术、PCR-DGGE和Illumina MiSeq高通量测序技术[4], 其中Illumina MiSeq高通量测序技术可以更精确地分析堆肥过程中微生物相对丰度和微生物群落组成[5].本文以典型的畜禽养殖粪便猪粪和果树修剪枝条桃树枝为主要原料进行堆肥, 采用Illumina MiSeq高通量测序技术, 研究堆肥化过程中理化参数等环境因子和微生物相对丰度和微生物群落结构组成的变化, 同时对理化指标进行监测, 并分析理化指标与微生物之间的相关关系, 揭示其微生物多样性及其变化规律, 以期为堆肥处理工艺的优化提供理论依据, 整体提升我国农业废弃物资源化利用的理论和技术水平, 并为实现我们农业面源农田综合防治与修复技术目标提供理论支持.
1 材料与方法 1.1 堆肥实验样本取自于北京市某有机肥厂, 该厂采用槽式好氧堆肥工艺, 将猪粪与桃树枝以及腐熟有机肥按体积比(立方):5:2.5:1的比例混合后进行槽式堆肥.其中猪粪来自于北京某农场, 桃树枝来源于北京某果园枝条修剪废弃物, 腐熟有机肥取自该有机肥厂上一批堆肥结束后的样品, 腐熟有机肥的原料同为猪粪和桃树枝, 堆肥原料的理化性质见表 1.用铲车将猪粪和辅料均匀搅拌, 每隔2 d使用翻抛机进行翻堆, 进行为期53 d的好氧堆肥.调整水分含量为59.07%, 有机碳(TOC)含量为390.28g·kg-1, 总氮(TN)含量为21.34g·kg-1.分别于第0、2、5、24、30、33、40和50 d从反应堆顶部、中部和底部进行3点取样, 均匀混合, 作为代表性样品.样品编号分别为d0、d2、d5、d24、d30、d33、d40和d50.样品分成两份:一份于4℃保存, 用于测定理化参数;一份储存于-20℃, 用于微生物多样性分析.
|
|
表 1 堆肥各原料的理化性质 Table 1 Physicochemical properties of raw compost materials |
1.2 理化参数测定
用DTSW-2型数字电子温度计(泰安德图自动化仪器有限公司)测定堆体顶端、中部、底部这3个位置的温度.用水分测定仪SH1 0A(上海菁海仪器有限公司)检测堆肥样品含水率.新鲜样品和蒸馏水按比例1:10(质量比)混合后用pH计PB-10(Sartorius德国)和DDS-307A型电导率仪(上海仪电科学仪器股份有限公司)分别测定堆肥样品的pH和EC.将样品研磨并过筛(40目筛), 按照国家标准[6](HJ 615-2011), 采用重铬酸钾-分光光度法测定TOC;TN含量参照国家标准[7](HJ 714-2014)采用凯氏法测定.NH4+-N含量参考国家标准[8](HJ 535-2009), 使用纳氏试剂分光光度法进行测定.种子发芽率采用Sun[9]的方法进行测定.种子发芽率根据以下公式计算:

|
将堆肥样品送上海美吉生物医药科技有限公司进行16S rDNA和18S rDNA高通量测序, 细菌的通用引物序列为:338F(5′-ACTCCTACGGGAGGCAG CAG-3′)和806R(5′- GGACTACHVGGGTWTCTAAT-3′);真菌的通用引物序列为SSU0817F(5′-TTAGC ATGGAATAATRRAATAGGA-3′)和1196R(5′-TCTG GACCTGGTGAGTTTCC-3′), 测序区域分别为V3-V4区和V5-V7区, 测序平台为IIIumina MiSeq.利用Usearch(vsesion 7.0)软件, 对序列进行聚类, 并将相似性大于0.97的序列定义为一个操作分类单元(OTU), 得到OTU表, 并进行生物信息统计分析.利用mothur (version v.1.30.1)进行微生物群落的α多样性分析, 计算参数为Chao1、Shannon和Simpson.Chao1指数, 由Chao于1984年提出, 用于计算微生物群落中含OTU数量[10].Shannon指数用于提供物种丰富度[11].利用Qiime平台进行物种分类, 生成不同分类水平上的物种丰度表.利用R语言软件中的vegan包进行群落组成分析.采用R语言软件进行主坐标分析(PCoA), 研究微生物群落结构相似性分析.采用R语言vegan包进行冗余分析(RDA), 研究微生物群落和环境因子之间的关系.多样性指数计算公式如下:
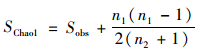
|
(1) |
式中, SChao1表示估计的OTU数;Sobs表示实际测得OTU数;n1表示只含一条序列的OTU数;n2表示只含两条序列的OTU数.
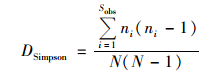
|
(2) |
式中, Sobs表示实际测得OTU数;ni表示第i个OTU所含序列数;N表示所有序列数.
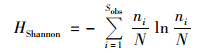
|
(3) |
式中, Sobs表示实际测得OTU数;ni表示第i个OTU所含序列数;N表示所有序列数.
1.4 数据分析应用统计软件Excel和Origin进行理化指标统计并绘制图表.
2 结果与讨论 2.1 堆体温度、pH和EC的变化温度是评价好氧堆肥过程的重要指标.由图 1可知第2 d堆肥迅速进入高温期(>50℃), 这是由于好氧微生物快速降解有机物时产生了大量的热[12].第28 d达到最高温60.4℃, 并且高温期持续至第32 d(高温持续时间>7 d), 杀死大量的病原菌, 达到堆肥的卫生标准[13].在堆肥高温阶段, 随着易分解物质的消耗, 微生物代谢强度降低, 导致温度下降, 但在转化成堆后, 高温好氧微生物再次繁殖, 开始缓慢消耗可分解的纤维素大分子, 从而温度上升[14].从第33 d起, 堆体进入降温阶段.

|
图 1 堆肥过程中温度的变化 Fig. 1 Temperature changes during composting |
pH反映堆肥过程中微生物所处的酸碱环境, 过高或过低的pH都会影响微生物生长和有机物的分解, 影响堆肥的进程.由图 2可知, 堆体初始pH为7.69, 随着堆肥进行pH不断上升, pH于30 d达到最大值8.87, 随后pH变化趋于平缓, 堆肥结束时pH为8.81.在堆肥初期, 一些易于分解的含氮有机物在微生物作用下发生氨化作用, 导致pH升高[15].随着堆肥过程进行, 由于微生物氨化作用减弱, 导致堆肥后期pH基本稳定不变[16].

|
图 2 堆肥过程中pH和EC的变化 Fig. 2 Changes in pH and EC during composting |
电导率反映了堆肥过程中可溶性盐的变化.由图 2可以看出, EC值总体表现为先下降后升高的趋势, 这可能因为在堆肥初期至高温阶段, 由于有机质的快速分解, 微生物活动消耗堆体的有机质以及利用水溶性盐物质满足自身生长需求, 导致EC在高温堆肥过程中先减少[17].随后, 由于堆肥过程中难降解物质的分解, 导致堆体的盐类物质产生使EC又逐渐上升[18], 至堆肥结束, 堆体的EC达到最大值4.5 mS·cm-1, 符合中国固体废弃物处理标准(EC<9 mS·cm-1), 基本满足农田使用对EC要求[19].
2.2 堆肥NH4+-N、TN和TOC的变化从图 3可以看出, NH4+-N含量整体呈现先上升后下降的趋势.堆肥初期, NH4+-N含量1.71 mg·g-1升高到5.62 mg·g-1(5 d), 之后NH4+-N开始下降至堆肥结束, 堆体末期的NH4+-N含量为1.92 mg·g-1, 因为在堆肥初期, 微生物分解代谢堆肥中残留的有机氮类物质产生铵态氮, 导致NH4+-N含量升高;而从堆肥的第10 d至30 d, 由于高温和pH升高导致NH4+-N以氨气的形式散失, 从而NH4+-N含量逐渐下降[20, 21].与第30 d相比, 第33 d的NH4+-N含量有所上升, 分析可能与氨化细菌的活动有关[22].此后, 由于堆肥进入降温阶段, NH4+-N含量下降减缓.

|
图 3 堆肥过程中铵态氮(NH4+-N)、总有机碳(TOC)和总氮(TN)的变化 Fig. 3 Changes in NH4+-N, TOC, and TN during composting |
堆肥过程中TN含量的变化如图 3所示, 堆肥初期TN快速下降, 主要由于初期堆体温度升高较快, 微生物大量消耗有机氮类物质以致产生高浓度的NH4+-N加强NH3的挥发[23], 导致堆肥TN下降.随后由于堆肥有机质大量微生物降解导致堆肥浓缩效应使得堆肥TN逐渐缓慢呈增加趋势[24].第33 d, TN含量为19.82 g·kg-1, 相比之前有所下降, 可能是由于高温期结束, 堆体的嗜温微生物活性增强, 分解有机物产生了氨气.第0 d时TN含量为21.87 g·kg-1, 堆肥结束TN含量为20.58 g·kg-1, 堆体氮含量损失了5.90%.TOC含量随堆肥的进行呈波动性变化.TOC含量下降可能是由于堆肥微生物分解碳水化合物并以CO2的形式挥发[25, 26], 以及类似总氮由于堆肥浓缩效应, 所以导致堆肥总有机碳初期先下降, 然后逐渐增加, 总体下降的变化趋势(图 3).
2.3 种子发芽指数(GI)变化GI是一种反映堆肥植物毒性和腐熟度的重要生物学指标[27], GI是植物对于堆肥低毒性(植物根长)和高毒性(发芽率)的综合反映.如图 4所示, 堆肥初期(0~5 d)的GI几乎为0, 这与堆肥物料本身具有严重的植物毒性有关[28].在堆肥中后期(24~40 d), GI出现了短暂的下降趋势, 这与Liu等[14]的研究结果相似, 原因可能是堆肥产生了低分子量的酸或挥发性脂肪酸[29].随着堆肥进行, 堆肥中有毒有害物质被逐渐降解, GI逐渐上升, 堆肥结束时, 种子发芽指数为73%.

|
图 4 堆肥过程中种子发芽指数(GI)的变化 Fig. 4 Changes in GI during composting |
微生物群落相似性可以通过主坐标分析(principal co-ordinates analysis, PCoA)[30].堆肥过程中不同样品中细菌群落的相似性如图 5(a)所示, 结果表明堆肥初期的样品d0、d2和d5相距较近, 说明堆肥前5 d细菌群落有高度的相似性;而堆肥高温期的样品(d24、d30、d33)相互间距离远, 说明堆肥高温期细菌群落变化明显, 可能是堆肥温度影响细菌的生长繁殖.而在堆肥末期的样品(d40、d50)相聚较近, 说明样品中的细菌群落有高度相似性, 降温阶段和腐熟期的细菌群落结构有着高度稳定性.

|
图 5 堆肥过程中各时间段细菌和真菌的PCoA分析 Fig. 5 PCoA analysis of bacteria and fungi at various time points during composting |
堆肥过程中不同样品中真菌群落的相似性如图 5(b)所示, 堆肥初期的样品(d0、d2、d5)、高温期的样品(d24、d30、d33)和腐熟期的样品(d50)同细菌群落的相似性类似, 第0、2、5 d的样品相距较近, 说明堆肥前5 d真菌群落有高度的相似性, 而与后期高温阶段的样品(d24、d30和d33)相距明显远, 说明堆肥初期和高温期的真菌微生物发生了明显的群落演替现象.堆肥末期(50 d)的真菌组成与前两个时期发生了明显变化.
2.4.2 微生物群落多样性分析Chao1指数反映样品中群落的丰富度, Shannon指数反映的是群落的多样性, 受样品群落中物种丰富度和物种均匀度的影响.如表 2所示, 随着堆肥的进行, 0~30 d堆体的细菌丰富度降低;堆肥30 d后, 随着堆温下降, 细菌丰富度增加, 在第40 d丰富度最大.相比建堆初期, d5、d30、d33、d40和d50样品的细菌多样性升高.真菌群落的丰富度变化总体呈先上升后下降的趋势;与0 d相比, 高温阶段的真菌群落多样性上升, 第50 d多样性下降.这表明:①温度的变化会影响微生物群落多样性;②不同好氧堆肥时期的微生物群落多样性不同, 这与王秀红等[31]的研究结果一致.
|
|
表 2 堆肥过程中不同时间点细菌群落和真菌群落的相对丰度和多样性指标 Table 2 Indices of relative bacterial and fungal abundances and diversity at different times during composting |
2.5 堆肥过程中细菌群落结构的变化
通过高通量测序技术可获得环境中微生物群落在数量和结构方面的信息, 分析细菌群落结构的动态变化, 结果如图 6(a)所示, 在细菌门分类水平上, 厚壁菌门(Firmicutes)和放线菌门(Actinobacteria)在整个堆肥过程中占主导地位, 为整个堆肥过程中的优势门类, 其相对丰度所占比例分别为79.31%~95.09%和2.98%~19.70%, 在一些报道中, 厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)均为堆肥过程中的细菌优势门类的一部分[12, 18, 32];其中, others代表相对丰度小于0.01的物种集合.在不同的堆肥时期, 细菌群落在各细菌门类上有不同的相对丰度:第0 d时, 厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)和变形菌门(Proteobacteria)所占比例分别为87.28%、9.64%和1.38%.随着堆肥的进行, 厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)的相对丰度在第24 d均达到最大值分别为95.09%和19.77%.厚壁菌门(Firmicutes)可以形成耐热孢子以抵抗高温, 同时厚壁菌门(Firmicutes)在高温期是最优势菌门, 其可以在高温下(>55℃)存活, 并能参与各种代谢活动[32].另外放线菌门(Actinobacteria)的相对丰度呈上升趋势, 其可以通过分泌各种抗生素抑制病原微生物[33].软壁菌门(Tenericutes)在堆肥第33 d时出现, 相对丰度为1.11%.

|
图 6 堆肥过程中细菌群落组成变化 Fig. 6 Changes in bacterial community composition during composting |
在细菌属水平上, 如图 6(b)所示, 堆肥初期, 主要优势菌属为Clostridium_sensu_stricto_1, Terrisporobacter和Bacillus.从第24 d一直到堆肥结束, 优势菌群为norank_f_Bacillaceae, Bacillus, Oceanbacillus和Pseudogracilibacillus, 这说明堆肥过程中细菌群落组发生了明显变化.Clostridium_sensu_stricto_1在堆肥过程中呈逐渐减少, 而norank_f_Bacillaceae逐渐成为堆肥过程中的主导菌属, 并且从堆肥初期至堆肥结束, Bacillus呈现增加, 然后维持相对稳定.Oceanbacillus菌属的变化情况类似Bacillus.并且其他菌属在堆肥过程中总体逐渐减少.而自动物肠道的微生物(Clostridium_sensu_stricto_1和Terrisporobacter菌属)经过堆肥后相对丰度下降, 这与黄雅楠等[34]的研究结果一致.
2.6 堆肥过程中真菌群落结构的变化如图 7(a), 在真菌门水平上, 堆肥初期和高温期后主要真菌门类有Ascomycota和Basidiomycota, 并且在整个堆肥过程中Ascomycota始终为优势门类, 主要是因为Ascomycota能够分泌多种纤维素、半纤维素降解酶, 能高效利用堆肥中的营养元素[35].在整个堆肥过程中, Basidiomycota相对丰度呈现先上升后下降的趋势, 在第40 d时其相对丰度达到最大值13.71%.

|
图 7 堆肥过程中真菌群落结构变化 Fig. 7 Changes in fungal community composition during composting |
从真菌属水平上看, 堆肥过程中各时间点的真菌群落变化如图 7(b)所示, 堆肥过程中真菌优势类群分别为unclassified_f_Trichocomaceae、norank_c_Sordariomycetes、Scopulariopsis和norank_o_Sordariales, 其中unclassified_f_Trichocomaceae比例在堆肥过程中基本呈现总体减少的趋势, Scopulariopsis没有明显的变化, 但是在堆肥过程中norank_c_Sordariomycetes的比例逐渐增加, 在堆肥末期成为优势类群, 据相关研究报道, norank_c_Sordariomycetes是分解堆肥中有机质的主要真菌类群, 可能由于Sordariomycetes具有耐高温和抗旱的微生物结构特征和孢子[36, 37].另外, norank_o_Sordariales的比例在堆肥过程中逐渐增加, 高温中期(24 d)达到最大随后略减少比例维持稳定, 研究表明在堆肥的嗜热阶段Sordariales有较高的相对丰度[38].
2.7 环境因子与微生物群落结构的相关关系采用冗余分析(RDA)分析堆肥体理化参数(温度、铵态氮、pH、TOC和TN)与主要菌属之间的关系.两个排序轴分别解释种群与环境积累变化率为85.10%、4.73%和69.14%、11.61%, 理化参数分别共同解释89.83%和80.75%的样本信息, 说明堆肥过程中理化参数与细菌和真菌群落结构变化有关.如图 8所示, 环境因子对细菌和真菌群落结构影响相关性排序均为pH>铵态氮>温度>TOC>TN, 该结果表明一方面堆肥过程中的微生物会造成有机酸的积累[39], 另一方面pH的变化会导致堆肥中钙和镁的变化, 从而间接影响微生物群落[40].含碳物质与含氮物质与细菌群落结构的相关性表明, 堆肥中的C、N含量会随着微生物代谢产物的不断转化而发生变化[32];另外氮源的质量和数量也可以控制细菌的群落结构变化[41].如图 8所示, 虽然温度不是与微生物群落结构变化相关性最大的环境因子, 但其仍可通过影响微生物的生物活性从而导致微生物群落结构的改变[42].在堆肥初期(第2 d和第5 d), 细菌群落结构较为相似;高温阶段和降温阶段, 细菌群落结构变化剧烈.此外, 在整个堆肥过程中, 真菌群落结构变化明显, 其中这可能与温度影响了真菌生长繁殖有关[43];Duan等[44]的研究结果表明, 温度在堆肥的高温期和腐熟阶段均会影响真菌的群落结构.从图 8(a)可以看出, 在细菌群落结构中, TOC、TN、铵态氮与Clostridium_sensu_stricto_1、Terrisporobacter呈正相关, 与Bacillus和norank_f_Bacillaceae呈负相关, 而Bacillus和norank_f_Bacillaceae与pH呈正相关.如图 8(b)中显示:norank_c_Sordariomycetes、norank_o_Sordariales和norank_c_Agaricomycetes与TN、pH呈正相关, 与TOC、铵态氮呈负相关, 说明norank_c_Sordariomycetes、norank_o_Sordariales和norank_c_Agaricomycetes可能与铵态氮的挥发有关.

|
图 8 堆肥过程中细菌和真群落结构与理化参数之间的冗余分析(属水平) Fig. 8 Redundancy analysis (RDA) between bacterial and fungal community structure and physical and chemical parameters during composting (genus level) |
(1) 堆肥高温期持续了30 d(>7 d), 达到粪便处理卫生标准;堆肥结束时EC为4.5 mS·cm-1, 符合中国固体废弃物处理标准(EC<9 mS·cm-1).利用好氧堆肥工艺, 以腐熟有机肥为外源添加物, 实现了对畜禽粪便和桃树枝堆肥的无害化和资源化.
(2) 利用高通量测序技术对堆肥过程中微生物的变化进行监测, 结果显示, 厚壁菌门和放线菌门是堆肥中的细菌优势门类;子囊菌门为真菌的优势门类;芽孢杆菌属是堆肥中后期的优势菌属.
(3) 对堆肥的理化指标进行冗余分析, 环境因子对细菌和真菌群落结构影响相关性排序均为pH>铵态氮>温度>TOC>TN, 其中pH对微生物群落组成影响最大.norank_c_Sordariomycetes、norank_o_Sordariales和norank_c_Agaricomycetes可能与铵态氮的挥发有关, 这为堆肥工艺的调节提供了理论参考.
| [1] | Yang X P, Li Q, Tang Z, et al. Heavy metal concentrations and arsenic speciation in animal manure composts in China[J]. Waste Management, 2017, 64: 333-339. DOI:10.1016/j.wasman.2017.03.015 |
| [2] | ten Hoeve M, Bruun S, Jensen L S, et al. Life cycle assessment of garden waste management options including long-term emissions after land application[J]. Waste Management, 2019, 86: 54-66. DOI:10.1016/j.wasman.2019.01.005 |
| [3] |
孙笑蕾, 胡正义, 刘莉, 等. 基于肥水资源化的河网区镇域农业面源污染控制系统的构建:以太湖地区新建镇为例[J]. 生态与农村环境学报, 2019, 35(5): 582-592. Sun X L, Hu Z Y, Liu L, et al. Pollutant reduction systems for controlling agricultural non-point-source pollution in town district of river network area based on reuse of wastewater and nutrient: A case study in Xinjian town, Taihu Lake[J]. Journal of Ecology and Rural Environment, 2019, 35(5): 582-592. |
| [4] | Lu C Y, Gu J, Wang X J, et al. Effects of coal gasification slag on antibiotic resistance genes and the bacterial community during swine manure composting[J]. Bioresource Technology, 2018, 268: 20-27. DOI:10.1016/j.biortech.2018.07.086 |
| [5] | Tian X P, Yang T, He J Z, et al. Fungal community and cellulose-degrading genes in the composting process of Chinese medicinal herbal residues[J]. Bioresource Technology, 2017, 241: 374-383. DOI:10.1016/j.biortech.2017.05.116 |
| [6] | HJ 615-2011, 土壤有机碳的测定重铬酸钾氧化-分光光度法[S]. |
| [7] | HJ 717-2014, 土壤质量全氮的测定凯氏法[S]. |
| [8] | HJ 535-2009, 水质氨氮的测定纳氏试剂分光光度法[S]. |
| [9] | Sun Z Y, Zhang J, Zhong X Z, et al. Production of nitrate-rich compost from the solid fraction of dairy manure by a lab-scale composting system[J]. Waste Management, 2016, 51: 55-64. DOI:10.1016/j.wasman.2016.03.002 |
| [10] | Chao A N. Non-parametric estimation of the number of the classes in a population[J]. Scandinavian Journal of Statistics, 1984, 11(4): 265-270. |
| [11] | Shannon C E, Weaver W. The mathematical theory of communication[M]. Urbana: The University of Illinois Press, 1963. |
| [12] | Wei H W, Wang L H, Hassan M, et al. Succession of the functional microbial communities and the metabolic functions in maize straw composting process[J]. Bioresource Technology, 2018, 256: 333-341. DOI:10.1016/j.biortech.2018.02.050 |
| [13] | Wu J, He S Z, Liang Y, et al. Effect of phosphate additive on the nitrogen transformation during pig manure composting[J]. Environmental Science and Pollution Research, 2017, 24(21): 17760-17768. DOI:10.1007/s11356-017-9285-x |
| [14] | Liu L, Wang S Q, Guo X P, et al. Succession and diversity of microorganisms and their association with physicochemical properties during green waste thermophilic composting[J]. Waste management, 2018, 73: 101-112. DOI:10.1016/j.wasman.2017.12.026 |
| [15] | Liu N, Zhou J L, Han L J, et al. Characterization of lignocellulosic compositions' degradation during chicken manure composting with added biochar by phospholipid fatty acid (PLFA) and correlation analysis[J]. Science of the Total Environment, 2017, 586: 1003-1011. DOI:10.1016/j.scitotenv.2017.02.081 |
| [16] | Liu N, Hou T, Yin H J, et al. Effects of amoxicillin on nitrogen transformation and bacterial community succession during aerobic composting[J]. Journal of Hazardous Materials, 2018, 362: 258-265. |
| [17] | Arab G, Razaviarani V, Sheng Z Y, et al. Benefits to decomposition rates when using digestate as compost co-feedstock: Part Ⅱ-Focus on microbial community dynamics[J]. Waste Management, 2017, 68: 74-84. DOI:10.1016/j.wasman.2017.07.018 |
| [18] | Zhou J M. Effect of turning frequency on co-composting pig manure and fungus residue[J]. Journal of the Air & Waste Management Association, 2017, 67(3): 313-321. |
| [19] | 聂永丰. 三废处理工程技术手册:固体废物卷[M]. 北京: 化学工业出版社, 2000. |
| [20] | Zhang L H, Zhang J C, Zeng G M, et al. Multivariate relationships between microbial communities and environmental variables during co-composting of sewage sludge and agricultural waste in the presence of PVP-AgNPs[J]. Bioresource Technology, 2018, 261: 10-18. DOI:10.1016/j.biortech.2018.03.089 |
| [21] | Wang Q, Wang Z, Awasthi M K, et al. Evaluation of medical stone amendment for the reduction of nitrogen loss and bioavailability of heavy metals during pig manure composting[J]. Bioresource Technology, 2016, 220: 297-304. DOI:10.1016/j.biortech.2016.08.081 |
| [22] | Qian X Y, Shen G X, Wang Z Q, et al. Co-composting of livestock manure with rice straw: characterization and establishment of maturity evaluation system[J]. Waste Management, 2014, 34(2): 530-535. DOI:10.1016/j.wasman.2013.10.007 |
| [23] | Zhang L H, Zeng G M, Dong H R, et al. The impact of silver nanoparticles on the co-composting of sewage sludge and agricultural waste: evolutions of organic matter and nitrogen[J]. Bioresource Technology, 2017, 230: 132-139. DOI:10.1016/j.biortech.2017.01.032 |
| [24] | Mao H, Lv Z Y, Sun H D, et al. Improvement of biochar and bacterial powder addition on gaseous emission and bacterial community in pig manure compost[J]. Bioresource Technology, 2018, 258: 195-202. DOI:10.1016/j.biortech.2018.02.082 |
| [25] | Zhou J M, Wang L T, Wang H M, et al. Effects of different ratios of pig manure to fungus residue on physicochemical parameters during composting[J]. Journal of the Air & Waste Management Association, 2016, 66(5): 499-507. |
| [26] | Huang C, Zeng G M, Huang D L, et al. Effect of Phanerochaete chrysosporium inoculation on bacterial community and metal stabilization in lead-contaminated agricultural waste composting[J]. Bioresource Technology, 2017, 243: 294-303. DOI:10.1016/j.biortech.2017.06.124 |
| [27] | Zhang D F, Luo W H, Li Y, et al. Performance of co-composting sewage sludge and organic fraction of municipal solid waste at different proportions[J]. Bioresource Technology, 2018, 250: 853-859. DOI:10.1016/j.biortech.2017.08.136 |
| [28] | Gong X Q, Cai L L, Li S Y, et al. Bamboo biochar amendment improves the growth and reproduction of Eisenia fetida and the quality of green waste vermicompost[J]. Ecotoxicology and Environmental Safety, 2018, 156: 197-204. DOI:10.1016/j.ecoenv.2018.03.023 |
| [29] | Paradelo R, Moldes A B, Barral M T. Evolution of organic matter during the mesophilic composting of lignocellulosic winery wastes[J]. Journal of Environmental Management, 2013, 116: 18-26. |
| [30] | Hu T, Wang X J, Zhen L S, et al. Effects of inoculating with lignocellulose-degrading consortium on cellulose-degrading genes and fungal community during co-composting of spent mushroom substrate with swine manure[J]. Bioresource Technology, 2019, 291: 121876. DOI:10.1016/j.biortech.2019.121876 |
| [31] |
王秀红, 李欣欣, 史向远, 等. 好氧堆肥微生物代谢多样性及其细菌群落结构[J]. 环境科学研究, 2018, 31(8): 1457-1463. Wang X H, Li X X, Shi X Y, et al. Microbial metabolism diversity and bacterial flora structure during aerobic composting[J]. Research of Environmental Sciences, 2018, 31(8): 1457-1463. |
| [32] | Zhou G X, Xu X F, Qiu X W, et al. Biochar influences the succession of microbial communities and the metabolic functions during rice straw composting with pig manure[J]. Bioresource Technology, 2019, 272: 10-18. DOI:10.1016/j.biortech.2018.09.135 |
| [33] |
杨萍萍, 尹华, 彭辉, 等. 外接菌种对污泥堆肥效能及堆体细菌群落的影响[J]. 环境科学, 2017, 38(8): 3536-3543. Yang P P, Yin H, Peng H, et al. Effects of exogenous microorganism inoculation on efficiency and bacterial community structure of sludge composting[J]. Environmental Science, 2017, 38(8): 3536-3543. |
| [34] |
黄雅楠, 王晓慧, 曹琦, 等. 高通量测序技术分析猪粪堆肥过程中微生物群落结构变化[J]. 微生物学杂志, 2018, 38(5): 21-26. Huang Y N, Wang X H, Cao Q, et al. Analysis of microbial community changes in pig excrement during compost process based on high-throughput sequencing technology[J]. Journal of Microbiology, 2018, 38(5): 21-26. DOI:10.3969/j.issn.1005-7021.2018.05.004 |
| [35] | Singh S, Madlala A M, Prior B A. Thermomyces lanuginosus: properties of strains and their hemicellulases[J]. FEMS Microbiology Reviews, 2003, 27(1): 3-16. DOI:10.1016/S0168-6445(03)00018-4 |
| [36] | Mbareche H, Veillette M, Bonifait L, et al. A next generation sequencing approach with a suitable bioinformatics workflow to study fungal diversity in bioaerosols released from two different types of composting plants[J]. Science of the Total Environment, 2017, 601-602: 1306-1314. DOI:10.1016/j.scitotenv.2017.05.235 |
| [37] | Gu W J, Lu Y S, Tan Z Y, et al. Fungi diversity from different depths and times in chicken manure waste static aerobic composting[J]. Bioresource Technology, 2017, 239: 447-453. DOI:10.1016/j.biortech.2017.04.047 |
| [38] | Morgenstern I, Powlowski J, Ishmael N, et al. A molecular phylogeny of thermophilic fungi[J]. Fungal Biology, 2012, 116(4): 489-502. DOI:10.1016/j.funbio.2012.01.010 |
| [39] | López-González J A, Suárez-Estrella F, Vargas-García M C, et al. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity[J]. Bioresource Technology, 2015, 175: 406-416. DOI:10.1016/j.biortech.2014.10.123 |
| [40] | Lucas R W, Klaminder J, Futter M N, et al. A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams[J]. Forest Ecology and Management, 2011, 262(2): 95-104. DOI:10.1016/j.foreco.2011.03.018 |
| [41] | Wallenstein M D, McMahon S, Schimel J. Bacterial and fungal community structure in Arctic tundra tussock and shrub soils[J]. FEMS Microbiology Ecology, 2007, 59(2): 428-435. DOI:10.1111/j.1574-6941.2006.00260.x |
| [42] | Wang J Q, Liu Z P, Xia J S, et al. Effect of microbial inoculation on physicochemical properties and bacterial community structure of citrus peel composting[J]. Bioresource Technology, 2019, 291: 121843. DOI:10.1016/j.biortech.2019.121843 |
| [43] |
张文浩, 门梦琪, 许本姝, 等. 牛粪稻秸新型静态堆肥中真菌群落组成的动态特征[J]. 农业环境科学学报, 2018, 37(9): 2029-2036. Zhang W H, Men M Q, Xu B S, et al. Dynamic characteristics of the composition of the fungal community in a novel static composting system of dairy manure and rice straw[J]. Journal of Agro-Environment Science, 2018, 37(9): 2029-2036. |
| [44] | Duan Y M, Awasthi S K, Chen H Y, et al. Evaluating the impact of bamboo biochar on the fungal community succession during chicken manure composting[J]. Bioresource Technology, 2019, 272: 308-314. DOI:10.1016/j.biortech.2018.10.045 |