 2020, Vol. 41
2020, Vol. 41



镉是一种对人体没有任何生理功能的有毒重金属.根据2014年发布的《全国土壤污染状况调查公报》, 我国土壤镉含量的点位超标率为7%, 是所有无机污染物的超标率中最高的[1].植物可食部位贡献了食物链和人体中大部分的镉, 植物中的镉又主要来源于根部对土壤镉的吸收[2].植物内没有镉的特异转运蛋白, 但镉可以通过一些金属营养元素的转运蛋白被植物非特异性地吸收[2].例如, 水稻OsIRT1(铁转运蛋白)的过表达可以显著提高水稻地上部和籽粒的镉含量[3];这种现象在钙、镁和锰等元素的转运蛋白上也有发现[4].
硒是人和动物必需的微量营养元素, 它也是人体多种必需蛋白质和酶的组成部分.虽然硒不是植物的必需元素[5], 但在适宜浓度时, 它可以降低植物对重金属的累积, 如对镉、砷、铅、铬和汞等[6~10].尤其是镉, 硒可以降低其在水稻[10]、小麦[11]和蔬菜[10, 12]等植物中的累积.硒有-2、0、+2、+4和+6这5种稳定价态.在自然环境中, 硒主要以硒酸盐和亚硒酸盐的形式存在;在不同的pH和pE条件下, 二者在无机环境中可以相互转化[13].硒酸盐主要通过高亲和力的硫酸盐转运蛋白被植物吸收, 而亚硒酸盐则通过正磷酸盐转运蛋白跨过质膜[5].因此, 电性相反的Cd2+和SeO32-/SeO42-在转运蛋白上不会发生吸收竞争;但是, 确实有研究表明, 一些阴离子可以显著改变细胞转运蛋白对金属离子的运输[14], 或是影响金属的吸收动力学和时间动态变化[15].
青菜是一种常见的芸薹属叶菜类蔬菜;它有较高的营养价值, 但也有较强的镉富集能力[16].Yang等[17]比较了6种蔬菜可食部位的镉含量, 发现青菜的镉含量在盆栽和大田试验中都是最高的.Liu等[18]也得到了相似的结果.同时, 不同品种青菜对镉的累积差异十分显著:Xia等[19]通过20种青菜的盆栽试验, 发现其地上部镉的含量范围是13.3~30.8 mg·kg-1, 最大差异为2.3倍.鉴于我国一些蔬菜产地的土壤镉污染情况以及蔬菜镉风险[20], 降低青菜可食部位镉含量对提高青菜品质、降低人体镉摄入风险和保证人体健康具有十分重要的意义.目前有关硒调控青菜中镉累积的研究并不透彻, 因此本研究选择了3个广泛种植的品种, 通过水培试验探究硒影响下的青菜对镉的基本吸收规律以及镉的吸收动力学, 旨在为降低青菜可食部位镉含量和人体镉摄入风险提供理论基础.
1 材料与方法 1.1 植物培养供试植物为青菜(Brassica rapa ssp. chinensis), 品种为上海青、苏州青根和杭州油冬儿.将青菜种子用去离子水洗净, 10% H2O2浸泡15 min消毒, 饱和CaSO4浸泡4 h吸水, 再将其用去离子水洗净、浮选.将饱满的种子插入蛭石0.5 cm深处发芽.播种14 d后, 选取长势一致的青菜幼苗移入容积为2.5 L的塑料盆里, 以营养液培养.营养液的组成如下(mmol·L-1):KNO3 1.0, MgSO4·7H2O 0.4, NH4H2PO4 0.5, Ca(NO3)2·4H2O 1.0, EDTA-Fe 0.03, H3BO3 3×10-3, MnSO4·H2O 10-3, ZnSO4·7H2O 10-3, CuSO4·5H2O 2×10-4, Na2MoO4 2×10-4;营养液的pH用KOH调节至5.8~6.0, 并使用1 mmol·L-1的2-(N-吗啉)乙磺酸一水合物[2-(N-morpholino) ethanesulfonic acid onohydrate, MES]缓冲[21].本试验在中国农业大学资环温室中进行, 控制生长条件为:温度25/20℃(昼/夜)、光照时间14 h·d-1、光照强度240~350 μmol·(m2·s)-1、相对湿度70%.整个试验中, 每3 d更换一次营养液.
1.2 试验设计暴露试验:青菜于营养液中培养23 d后(播种37 d后), 向基本营养液中额外添加镉和硒;每盆2株青菜, 作为1个重复.镉的处理浓度为10 μmol·L-1的Cd(NO3)2·4H2O;硒只以Na2SeO3的形式添加, 浓度设置分别为0、10、20和50 μmol·L-1.每个处理有3个生物学重复, 处理3 d后收获样品;处理期间不更换营养液.样品的后续采集和分析均按照每个处理2株青菜计算均值.
镉的吸收动力学:杭州油冬儿于营养液中培养26 d后(播种40 d后), 收获长势一致的植株并去除其地上部.将新鲜的根部放入200 mL以1 mmol·L-1的Ca(NO3)2和MES为缓冲(NaOH调节pH至5.8)的含硒、镉的吸收液中进行镉的吸收动力学试验.硒以硒酸盐和亚硒酸盐的形式添加, 设置3个硒水平, 0、5和20 μmol·L-1(Na2SeO3或Na2SeO4;分别记为SeIV和SeVI);设置7个镉水平, 1、10、20、40、80、160和320 μmol·L-1的Cd(NO3)2·4H2O.吸收试验在室温条件下进行, 每个处理重复3次, 吸收1 h后收获.
1.3 样品采集和元素含量测定将收获的植株分为地上部和根部(吸收动力学的仅为根部).根部在洗脱液中浸泡15 min(4℃);洗脱液的组成为1 mmol·L-1的CaSO4和MES(pH为5.8).将地上部和洗脱后的根部用去离子水洗净, 并称量其鲜重.将样品在烘箱内杀青30 min(105℃), 随后烘干48 h(75℃);称量干样的干重后将其粉碎.将0.25 g干样以HNO3密闭式微波消解法消解(MARS5;CEM, 美国);使用ICP-AES(Optima 7300 DV;PerkinElmer, 美国)测定消解液中镉、铁和锰的浓度(吸收动力学仅测定镉的浓度).整个测定过程中, 加入空白样品和标准物质[镉含量为(0.19±0.02) mg·kg-1;GSB-27大葱, 中国计量科学院]进行质量控制;所有元素的回收率为95%~105%.
1.4 数据处理所有试验数据以鲜重(fresh weight, FW)的均值±标准误差(standard errors, , SE;n=3)表示.按照式(1)~(4)分别计算青菜地上部和根部吸收的镉总量、每克根的镉吸收量、镉的转移系数和镉在地上部的分配比:
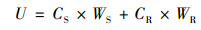
|
(1) |
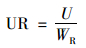
|
(2) |

|
(3) |
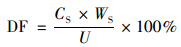
|
(4) |
式中, U、UR、TF和DF分别是青菜每株镉总量(μg)、每克根镉吸收量(μg·g-1)、转移系数和地上部分配比;CS和CR分别是青菜地上部和根部的镉含量(μg·g-1即mg·kg-1), WS和WR分别是青菜地上部和根部的生物量(g).
采用Sigmaplot 12.5拟合米氏方程来描述镉的吸收动力学[22]:
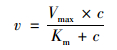
|
(5) |
式中, v表示单位质量根系吸收镉的速率[μg·(g·h)-1];Km为米氏常数, 是镉达到最大吸收速率的1/2时所对应的溶液镉浓度(μmol·L-1);Vmax表示根系所能达到的最大镉吸收速率[μg·(g·h)-1];c表示吸收液中镉的浓度(μmol·L-1).
使用SAS 9.0最小显著检验对所有数据进行方差分析(LSD;P<0.05), 使用Simaplot 12.5和Excel 2016作图.
2 结果与分析 2.1 生物量总体来说, 青菜的地上部和根部生物量受品种或亚硒酸盐的影响较小(图 1). 3个品种青菜的地上部鲜重基本在30~50 g之间, 而根部生物量则在3 g左右.尽管亚硒酸盐略微降低了上海青和苏州青根的地上部和根部生物量, 但所有差异均小于22%, 且均不显著.杭州油冬儿的生物量对硒水平较其他两个品种更敏感.和单独加镉相比, 10 μmol·L-1的亚硒酸盐使其地上部和根部生物量均提高了28%.但是, 随着硒处理水平的提高, 杭州油冬儿的鲜重逐渐降低, 和不加硒相比, 50 μmol·L-1亚硒酸盐使其根部和地上部的鲜重均下降了1/3左右;而和10 μmol·L-1亚硒酸盐相比, 此差异接近1/2, 且达到显著水平(P<0.05;图 1).

|
结果以均值+标准误差(n=3)表示;Se0、Se10、Se20和Se50分别代表亚硒酸盐处理浓度为0~50 μmol·L-1, 下同;不同字母表示同一品种青菜地上部或根部鲜重的显著差异(LSD;P<0.05), 下同 图 1 不同浓度亚硒酸盐处理下青菜地上部和根部的鲜重 Fig. 1 Fresh weights of shoot and root of pak choi affected by different levels of selenite |
不同品种青菜地上部的镉含量对亚硒酸盐的响应差别明显[图 2(a)].与不加硒的相比, 3个浓度的亚硒酸盐均显著降低了杭州油冬儿地上部镉含量(P<0.05), 但不同亚硒酸盐水平的效果没有差异, 降幅都约为50%[图 2(a)].亚硒酸盐处理下苏州青根和上海青的地上部镉含量与不加硒的之间没有显著差异:10 μmol·L-1亚硒酸盐使它们地上部镉含量分别提高了15%和25%;但是, 当亚硒酸盐水平高于10 μmol·L-1时, 苏州青根和上海青的地上部镉累积开始受到抑制, 如50 μmol·L-1亚硒酸盐使它们的地上部镉含量分别降低了30%和51%[图 2(a)].不添加亚硒酸盐的杭州油冬儿地上部镉含量显著比上海青和苏州青根高100%和35%, 但是添加亚硒酸盐后, 它们的地上部镉含量差异缩小, 到硒50 μmol·L-1时, 杭州油冬儿比另外两个品种的地上部镉含量分别高36%和32%.

|
不同字母表示同一品种青菜地上部或根部镉含量的显著差异(LSD; P<0.05) 图 2 不同浓度亚硒酸盐处理下青菜地上部和根部的镉含量(鲜重) Fig. 2 Cd content in shoot and root of pak choi affected by different levels of selenite (FW) |
在不同亚硒酸盐水平下, 3个品种青菜的根部镉含量间的差异均不显著;尤其对于上海青和苏州青根, 其差异均在15%以内[图 2(b)].和地上部不同的是, 杭州油冬儿的根部镉含量对亚硒酸盐的浓度梯度却较为敏感:10 μmol·L-1和20 μmol·L-1亚硒酸盐分别使其根部镉含量降低42%和30%, 而50 μmol·L-1硒处理的根部镉含量甚至略微高于不加硒的(图 2).另外, 上海青的根部仍然累积了最少的镉, 但不加硒时, 根部镉含量的品种间差异远小于地上部.这3种青菜的根部镉含量在40~60 mg·kg-1之间, 而地上部的镉则约为其含量的1/10(图 2).
亚硒酸盐可以显著影响镉从青菜根部向地上部的转移(表 1).尤其在杭州油冬儿和苏州青根中, 它们的地上部镉分配比和转移系数均随着亚硒酸盐添加量升高而降低. 50 μmol·L-1亚硒酸盐处理下, 杭州油冬儿的分配比和转移系数相较不加硒的分别降低了29%和52%, 苏州青根的差异则分别为33%和54%, 差异均显著(P<0.05;表 1).对于上海青, 虽然亚硒酸盐没有影响其镉转移系数和地上部分配比, 但相比于单独加镉的, 50 μmol·L-1硒仍使这2个系数降低了15%和43%.
|
|
表 1 亚硒酸盐处理的青菜地上部镉分配比和转移系数1) Table 1 Distribution of Cd in pak choi shoots and translocation factors influenced by different levels of selenite |
总体来说, 杭州油冬儿中的铁、锰含量对亚硒酸盐处理更为敏感, 并存在比镉更明显的梯度关系(图 3和图 4).亚硒酸盐没有显著影响上海青和苏州青根的地上部铁含量;和不加硒的对比, 它们地上部铁含量的差异小于20%[图 3(a)].杭州油冬儿的地上部铁含量则随着亚硒酸盐浓度的升高而上升:和不加硒的相比, 10、20和50 μmol·L-1亚硒酸盐分别使其增加了7%、18%和39%, 且50 μmol·L-1时的差异显著(P<0.05;LSD).在根部, 3个品种青菜的根部铁含量也基本随亚硒酸盐添加量的增加而增加;与不加硒的相比, 50 μmol·L-1亚硒酸盐处理的上海青、苏州青根和杭州油冬儿的根部铁含量的增幅分别为36%、12%和50%[图 3(b)].

|
不同字母表示青菜地上部或根部铁含量的显著差异(LSD; P<0.05) 图 3 不同浓度亚硒酸盐处理下青菜地上部和根部的铁含量(鲜重) Fig. 3 Fe content in shoot and root of pak choi affected by different levels of selenite (FW) |
亚硒酸盐对上海青和杭州油冬儿地上部锰累积的促进更为明显, 而对苏州青根的影响却小于5%[图 4(a)].上海青和杭州油冬儿的地上部锰含量均随着亚硒酸盐添加量的增加而增加:和不加硒的相比, 杭州油冬儿在3个梯度硒处理下的增幅分别为4%、15%和31%, 50 μmol·L-1硒处理的差异显著(P<0.05;LSD).此外, 青菜根部锰含量的差异更小:10μmol·L-1和50 μmol·L-1的亚硒酸盐使上海青的根部锰含量分别降低了15%和增加了42%;其余两个品种青菜的根部锰含量的变化均在20%以内[图 4(b)].

|
不同字母表示青菜地上部或根部锰含量的显著差异(LSD; P<0.05) 图 4 不同浓度亚硒酸盐处理下青菜地上部和根部的锰含量(鲜重) Fig. 4 Mn contents in the shoot and root of pak choi as affected by different levels of selenite (FW) |
无论是否添加硒, 米氏方程拟合杭州油冬儿镉的吸收动力学的相关系数R都大于0.9878(表 2).未添加硒(Se0)时, 杭州油冬儿的吸收曲线在镉320 μmol·L-1时达到平台, 根部吸收速率接近最大(图 5).添加亚硒酸盐和硒酸盐都显著提高了杭州油冬儿对镉的吸收速率. 5μmol·L-1和20 μmol·L-1亚硒酸盐均使杭州油冬儿的Vmax提高了超过1倍, 但Km仅分别被提高了61%和45%(表 2). 5和20 μmol·L-1硒酸盐处理下的Vmax分别与等水平亚硒酸盐处理下的相近, 但硒酸盐处理的Km更低(即镉在更低浓度时即可达到最大吸收速率).两种无机硒处理下的镉吸收速率相差不到10%(图 5), 拟合参数十分接近(表 2), 同种硒处理的镉吸收速率在两个水平之间的差异也小于10%.
|
|
表 2 杭州油冬儿镉的吸收动力学参数1) Table 2 Michaelis-Menten kinetics paramters for Cd of Hangzhouyoudonger |

|
镉的吸收速率是1g鲜重青菜根部在1h内所吸收镉的质量 图 5 不同浓度亚硒酸盐和硒酸盐影响下杭州油冬儿镉的吸收动力学 Fig. 5 Uptake kinetics of Cd in Hangzhouyoudonger shoot influenced by different levels of selenite or selenate |
芸薹属的青菜对镉有较强的累积能力[17].本研究中, 青菜表现出了很强的镉富集能力:10 μmol·L-1镉(约1.13 mg·L-1)处理下的青菜地上部的镉含量均高于3 mg·kg-1, 根部的含量甚至全部高于35 mg·kg-1(图 2).生物量是反映环境胁迫的重要指标.本研究中, 3个浓度亚硒酸盐(>10 μmol·L-1)处理下青菜的生物量间无显著差异(图 1).Zembala等[11]的研究发现, 2 μmol·L-1硒酸钠显著提高了甘蓝型青菜和小麦地上部的生物量.Feng等[23]则指出, 硒对生物量的效应依赖于硒水平;他们发现1.27 μmol·L-1和12.7 μmol·L-1亚硒酸盐使水稻根部的生物量分别提高了18%和44%;当硒水平达到63.5 μmol·L-1时, 地上部和根部的生物量却被降低了一半以上, 因为硒在较高浓度的时候会表现出毒性[23].
虽然硒不是植物的必需元素[5], 但它却可以降低植物对镉的累积和转移.本研究发现, 亚硒酸盐抑制了青菜对镉的累积, 尤其可以降低其地上部(可食部位)的镉含量.亚硒酸盐的此效果在其它植物中也有报道, 如水稻[24]、小麦[11]和蔬菜[10, 12]等.Sun等[25]的研究发现, 3 μmol·L-1亚硒酸盐使50 μmol·L-1镉处理的黄瓜叶片和根部的镉含量分别显著降低了21%和32%.Wan等[15]的研究发现, 5 μmol·L-1亚硒酸盐使等物质的量浓度镉处理的水稻的镉吸收量降低了42%.周健等[26]则发现, 盆栽条件下, 1 mg·kg-1的亚硒酸盐使5 mg·kg-1镉处理的地上部镉含量显著下降了20%左右, 而硒酸盐却促进了镉的累积.
青菜根部的镉含量远高于地上部的(图 2), 这说明青菜会自发地将镉固定在根中, 防止镉进一步毒害地上部.该现象与大多数植物的相同[10, 15].本研究中, 亚硒酸盐使青菜镉的地上部分配比和转移系数均有不同程度地下降(表 1).同样的结果在其它植物上也有发现:5 μmol·L-1硒酸盐和亚硒酸盐分别使水稻中镉的地上部分配比降低了近50%[15].植物体内大部分的镉与含硫有机物中的硫结合[27].硒与硫均为氧族元素, 它们化学性质相似, 所以硒可能通过取代硫而与镉结合, 与镉结合形成难移动的复合物并促进镉在液泡[28]和细胞壁中[29]的沉积, 从而抑制镉在植物内的移动.
除了受环境因素的影响外, 植物对镉的吸收还受到植物本身基因型(品种)的影响.Yang等[17]在含2 mg·kg-1镉的土壤中种植了6种不同蔬菜, 发现它们可食部位的镉含量范围是0.55~10.30 mg·kg-1, 最大相差接近20倍.Zhang等[30]在含镉0.13~1.25 mg·kg-1的土壤上种植了28种莴苣, 发现其地上部镉含量间的最大差异约为5倍.Xia等[19]通过20种青菜的盆栽试验, 发现其地上部镉的含量范围是13.3~30.8 mg·kg-1, 最大差异为2.3倍.在亚硒酸盐的调控下, 3种青菜在镉累积和转移都有非常不同的响应(图 2).不同品种的植物的部分基因表达可能存在显著差别.例如, 水稻高累积品种Anjana dhan和低累积品种Sasanishiki的OsNRAMP1的氨基酸序列完全一致, 但该转运蛋白基因在前者根部的表达量是后者的6倍[31].因此, 尽管镉的吸收和运输过程在青菜中基本相同, 但该过程在硒处理下的表达程度决定了硒在宏观上对不同品种镉累积、转移表现出不同的效果.
在本研究中, 尽管适宜浓度的硒可以降低青菜地上部的镉含量, 但是对于一些和镉共用转运蛋白的必需金属元素如铁、锰等, 硒却在总体上提高了它们在青菜地上部的累积(图 3和图 4).Liu等[32]的研究发现, 50 μmol·L-1的CdCl2使水稻地上部锰和锌的含量降低了2/3以上, 而3 μmol·L-1的亚硒酸钠处理和其相比, 使两种元素的含量分别显著提高了304%和39%.相似的结果在甘蓝型青菜中也有发现[11].而这些矿质元素又是一些抗氧化酶和叶绿素的重要组成部分, 因此此现象可能会进一步影响植物生理代谢从而缓解镉的毒性[33].
镉和硒在土壤环境中以及本试验营养液pH(5.8~6.0)的条件下, 分别以CdOH+和HSeO3-/HSeO42-为主要形态[34].SeO32-/SeO42-分别通过植物根表质膜的正磷酸盐转运蛋白和高亲和力的硫酸盐转运蛋白被吸收[5].Cd2+主要由一些二价金属元素转运蛋白运输[2].因此, 电性不同的无机硒和镉离子在根表不存在转运蛋白上的竞争.尽管如此, 亚硒酸盐和硒酸盐仍然显著提高了两种青菜根部对镉的吸收速率(图 5和表 2).Wan等[15]也发现, 在1 h内, 亚硒酸盐提高了水稻根部对镉的吸收速率.植物对金属离子的吸收在很大程度上受环境中阴离子的影响[14].Cl-和SO42-等阴离子有促进重金属离子吸收的效果[35]. Smolders等[36]认为, 当排除土壤环境的作用后, SO42-可以和自由镉离子Cd2+在营养液中形成CdSO40, 起到缓冲的作用[37], Cl-则通过与Cd2+结合产生CdCln2-n而产生同样的作用.植物对这两种配合物的吸收效率高于Cd2+, 所以Cl-和SO42-表现出提高镉吸收速率的效果.此外, 共质体含有丰富的电荷, 这些电荷对镉的吸收产生了较大的阻力[38].这些阴离子恰好可以削弱镉的吸收限制.值得注意的是, 两种形态的硒对青菜的镉吸收速率的促进幅度基本一致(表 2);因此, 硒酸盐和亚硒酸盐促进镉吸收速率的效果应该更多地来源于它们所带的负电荷(以充当离子缓冲剂), 而不是硒元素的效果;这也从侧面说明硒对镉累积的调控效果主要来源于植物内部的过程, 而非根表的吸收竞争.另外, 通过米氏方程拟合得到, 杭州油冬儿的Vmax在没有硒的情况下也可超过200 μg·(g·h)-1(根中, 下同), 而在有硒的情况下甚至能超过800 μg·(g·h)-1, 远高于水稻的水平[36], 这说明青菜根部具有很强的镉吸收能力.
4 结论(1) 不同品种青菜地上部对不同浓度的亚硒酸盐的响应不同:10~50 μmol·L-1亚硒酸盐均使杭州油冬儿地上部镉含量显著降低50%左右;苏州青根和上海青的地上部镉含量随硒处理的水平提高而降低, 但降幅不显著.
(2) 不同浓度的亚硒酸盐均可以抑制苏州青根和杭州油冬儿中的镉从根部向地上部的转移, 且随着硒处理浓度的升高, 抑制效果更加明显.亚硒酸盐对上海青中镉的转移的影响不明显.
(3) 亚硒酸盐总体提高了青菜中与镉共用转运蛋白的铁、锰的含量, 尤其是在杭州油冬儿中.
(4) 硒酸盐和亚硒酸盐均显著提高了青菜对镉的吸收速率, 并且提高了米氏方程拟合得到的最大吸收速率.但是, 二者也提高了米氏常数, 即镉需要在更高的浓度才能达到最大吸收速率.
| [1] | 环境保护部和国土资源部.全国土壤污染状况调查公报[DB/OL]. http://www.ndrc.gov.cn/fzgggz/ncjj/zhdt/201404/t20140418_607888.html, 2014-04-17. |
| [2] | Clemens S, Aarts M G M, Thomine S, et al. Plant science: the key to preventing slow cadmium poisoning[J]. Trends in Plant Science, 2013, 18(2): 92-99. |
| [3] | Nakanishi H, Ogawa I, Ishimaru Y, et al. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice[J]. Soil Science and Plant Nutrition, 2006, 52(4): 464-469. DOI:10.1111/j.1747-0765.2006.00055.x |
| [4] | Sasaki A, Yamaji N, Yokosho K, et al. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice[J]. The Plant Cell, 2012, 24(5): 2155-2167. DOI:10.1105/tpc.112.096925 |
| [5] | Terry N, Zayed A M, De Souza M P, et al. Selenium in higher plants[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 2000, 51: 401-432. DOI:10.1146/annurev.arplant.51.1.401 |
| [6] | Mroczek-Zdyrska M, Wójcik M. The influence of selenium on root growth and oxidative stress induced by lead in Vicia faba L. minor plants[J]. Biological Trace Element Research, 2012, 147(1-3): 320-328. DOI:10.1007/s12011-011-9292-6 |
| [7] | Hu Y, Duan G L, Huang Y Z, et al. Interactive effects of different inorganic As and Se species on their uptake and translocation by rice (Oryza sativa L.) seedlings[J]. Environmental Science and Pollution Research, 2014, 21(5): 3955-3962. DOI:10.1007/s11356-013-2321-6 |
| [8] | Qing X J, Zhao X H, Hu C X, et al. Selenium alleviates chromium toxicity by preventing oxidative stress in cabbage (Brassica campestris L. ssp. Pekinensis) leaves[J]. Ecotoxicology and Environmental Safety, 2015, 114: 179-189. DOI:10.1016/j.ecoenv.2015.01.026 |
| [9] | Zhang H, Feng X B, Zhu J M, et al. Selenium in soil inhibits mercury uptake and translocation in rice (Oryza sativa L.)[J]. Environmental Science & Technology, 2012, 46(18): 10040-10046. |
| [10] | Wu Z C, Wang F H, Liu S, et al. Comparative responses to silicon and selenium in relation to cadmium uptake, compartmentation in roots, and xylem transport in flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis) under cadmium stress[J]. Environmental and Experimental Botany, 2016, 131: 173-180. DOI:10.1016/j.envexpbot.2016.07.012 |
| [11] | Zembala M, Filek M, Walas S, et al. Effect of selenium on macro- and microelement distribution and physiological parameters of rape and wheat seedlings exposed to cadmium stress[J]. Plant and Soil, 2010, 329(1-2): 457-468. DOI:10.1007/s11104-009-0171-2 |
| [12] | He P P, Lv X Z, Wang G Y. Effects of Se and Zn supplementation on the antagonism against Pb and Cd in vegetables[J]. Environment International, 2004, 30(2): 167-172. DOI:10.1016/S0160-4120(03)00167-3 |
| [13] | Elrashidi M A, Adriano D C, Workman S M, et al. Chemical-equilibria of selenium in soils - a theoretical development[J]. Soil Science, 1987, 144(2): 141-152. DOI:10.1097/00010694-198708000-00008 |
| [14] | Nelson N. Metal ion transporters and homeostasis[J]. The EMBO Journal, 1999, 18(16): 4361-4371. DOI:10.1093/emboj/18.16.4361 |
| [15] | Wan Y N, Yu Y, Wang Q, et al. Cadmium uptake dynamics and translocation in rice seedling: Influence of different forms of selenium[J]. Ecotoxicology and Environmental Safety, 2016, 133: 127-134. DOI:10.1016/j.ecoenv.2016.07.001 |
| [16] | Wang C L, Liu Y G, Zeng G M, et al. Mechanism of exogenous selenium alleviates cadmium induced toxicity in Bechmeria nivea (L.) Gaud (Ramie)[J]. Transactions of Nonferrous Metals Society of China, 2014, 24(12): 3964-3970. DOI:10.1016/S1003-6326(14)63557-3 |
| [17] | Yang Y, Zhang F S, Li H F, et al. Accumulation of cadmium in the edible parts of six vegetable species grown in Cd-contaminated soils[J]. Journal of Environmental Management, 2009, 90(2): 1117-1122. DOI:10.1016/j.jenvman.2008.05.004 |
| [18] | Liu L, Hu L L, Tang J J, et al. Food safety assessment of planting patterns of four vegetable-type crops grown in soil contaminated by electronic waste activities[J]. Journal of Environmental Management, 2012, 93(1): 22-30. |
| [19] | Xia S L, Deng R B, Zhang Z, et al. Variations in the accumulation and translocation of cadmium among pak choi cultivars as related to root morphology[J]. Environmental Science and Pollution Research, 2016, 23(10): 9832-9842. DOI:10.1007/s11356-016-6210-7 |
| [20] |
沈彤, 刘明月, 贾来, 等. 长沙地区蔬菜重金属污染初探[J]. 湖南农业大学学报(自然科学版), 2005, 31(1): 87-90. Shen T, Liu M Y, Jia L, et al. On the heavy metal pollution of vegetables in Changsha region[J]. Journal of Hunan Agricultural University (Natural Sciences), 2005, 31(1): 87-90. |
| [21] | Yu Y, Yuan S L, Zhuang J, et al. Effect of selenium on the uptake kinetics and accumulation of and oxidative stress induced by cadmium in Brassica chinensis[J]. Ecotoxicology and Environmental Safety, 2018, 162: 571-580. DOI:10.1016/j.ecoenv.2018.07.041 |
| [22] | Wilkinson G N. Statistical estimations in enzyme kinetics[J]. Biochemical Journal, 1961, 80(2): 324-332. |
| [23] | Feng R W, Wei C Y, Tu S X, et al. A dual role of se on cd toxicity: Evidences from the uptake of Cd and some essential elements and the growth responses in paddy rice[J]. Biological Trace Element Research, 2013, 151(1): 113-121. DOI:10.1007/s12011-012-9532-4 |
| [24] | Lin L, Zhou W H, Dai H X, et al. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice[J]. Journal of Hazardous Materials, 2012, 235-236: 343-351. DOI:10.1016/j.jhazmat.2012.08.012 |
| [25] | Sun H Y, Dai H X, Wang X Y, et al. Physiological and proteomic analysis of selenium-mediated tolerance to Cd stress in cucumber (Cucumis sativus L.)[J]. Ecotoxicology and Environmental Safety, 2016, 133: 114-126. DOI:10.1016/j.ecoenv.2016.07.003 |
| [26] |
周健, 郝苗, 刘永红, 等. 不同价态硒缓解小油菜镉胁迫的生理机制[J]. 植物营养与肥料学报, 2017, 32(2): 444-450. Zhou J, Hao M, Liu Y H, et al. Physiological mechanism of different valences selenium in relieving cadmium stress of rape[J]. Journal of Plant Nutrition and Fertilizer, 2017, 32(2): 444-450. |
| [27] | DalCorso G, Farinati S, Maistri S, et al. How plants cope with cadmium: Staking all on metabolism and gene expression[J]. Journal of Integrative Plant Biology, 2008, 50(10): 1268-1280. DOI:10.1111/j.1744-7909.2008.00737.x |
| [28] | Mendoza-Cózatl D G, Jobe T O, Hauser F, et al. Long-distance transport, vacuolar sequestration, tolerance, and transcriptional responses induced by cadmium and arsenic[J]. Current Opinion in Plant Biology, 2011, 14(5): 554-562. DOI:10.1016/j.pbi.2011.07.004 |
| [29] | Zhu X F, Wang Z W, Dong F, et al. Exogenous auxin alleviates cadmium toxicity in Arabidopsis thaliana by stimulating synthesis of hemicellulose 1 and increasing the cadmium fixation capacity of root cell walls[J]. Journal of Hazardous Materials, 2013, 263: 398-403. DOI:10.1016/j.jhazmat.2013.09.018 |
| [30] | Zhang K, Yuan J G, Kong W, et al. Genotype variations in cadmium and lead accumulations of leafy lettuce (Lactuca sativa L.) and screening for pollution-safe cultivars for food safety[J]. Environmental Science: Processes & Impacts, 2013, 15(6): 1245-1255. |
| [31] | Takahashi R, Ishimaru Y, Senoura T, et al. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice[J]. Journal of Experimental Botany, 2011, 62(14): 4843-4850. DOI:10.1093/jxb/err136 |
| [32] | Liu H W, Wang H Y, Ma Y B, et al. Role of transpiration and metabolism in translocation and accumulation of cadmium in tobacco plants (Nicotiana tabacum L.)[J]. Chemosphere, 2016, 144: 1960-1965. DOI:10.1016/j.chemosphere.2015.10.093 |
| [33] | Filek M, Keskinen R, Hartikainen H, et al. The protective role of selenium in rape seedlings subjected to cadmium stress[J]. Journal of Plant Physiology, 2008, 165(8): 833-844. DOI:10.1016/j.jplph.2007.06.006 |
| [34] | Sandrin T R, Maier R M. Effect of pH on cadmium toxicity, speciation, and accumulation during naphthalene biodegradation[J]. Environmental Toxicology & Chemistry, 2002, 21(10): 2075-2079. |
| [35] | Ozkutlu F, Ozturk L, Erdem H, et al. Leaf-applied sodium chloride promotes cadmium accumulation in durum wheat grain[J]. Plant and Soil, 2007, 290(1-2): 323-331. DOI:10.1007/s11104-006-9164-6 |
| [36] | Smolders E, McLaughlin M J. Chloride increases cadmium uptake in Swiss chard in a resin-buffered nutrient solution[J]. Soil Science Society of America Journal, 1996, 60(5): 1443-1447. DOI:10.2136/sssaj1996.03615995006000050022x |
| [37] | McLaughlin M J, Andrew S J, Smart M K, et al. Effects of sulfate on cadmium uptake by Swiss chard: I. Effects of complexation and calcium competition in nutrient solutions[J]. Plant and Soil, 1998, 202(2): 211-216. DOI:10.1023/A:1004333529886 |
| [38] | Grignon C, Sentenac H. pH and ionic conditions in the apoplast[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1991, 42: 103-128. DOI:10.1146/annurev.pp.42.060191.000535 |