 2019, Vol. 40
2019, Vol. 40

2. 南阳师范学院生命科学与技术学院, 南阳 473061
 , WANG Xiao-yu2 , CAI Hong2 , YAO Lun-guang1 , CAI Qian-di1 , WANG Yi-xue1 , CHEN Zhao-jin1
, WANG Xiao-yu2 , CAI Hong2 , YAO Lun-guang1 , CAI Qian-di1 , WANG Yi-xue1 , CHEN Zhao-jin1
2. School of Life Science and Technology, Nanyang Normal University, Nanyang 473061, China
随着工农业的快速发展以及耕地中农业投入品用量的不断增加, 大量的Cd和Pb进入土壤, 不仅会造成粮食减产和经济损失, 更严重威胁着粮食安全[1~3].据文献[4]显示我国受重金属污染土壤的总点位超标率为16.1%, 其中Cd的超标率最高, 达到7.0%.无机重金属污染物, 如:Cd、Pb和Cu等, 在土壤中移动性差、滞留时间长、不能被微生物降解, 严重恶化土壤环境质量, 并可经水和植物等介质进入人体, 最终严重影响人类健康[5~7].重金属超标土壤的农业利用是在我国耕地资源十分紧张, 粮食和食物安全形势十分严峻前提下的一种不得已的选择.在此前提下, 寻求边利用和边修复的有效途径具有重要的意义[8, 9].
植物促生细菌(plant growth-promoting bacteria, PGPB)是在植物根际系统定殖、能够明显促进植物生长的一类有益细菌的总称[10].近年来, 某些具有特殊功能的PGPB作为优良的重金属钝化剂被广泛研究和应用[11].重金属固定植物促生细菌不仅能够使重金属在土壤中沉淀或被吸附固定, 改变土壤重金属赋存状态, “钝化”其生物有效性, 从而降低作物对重金属的吸收, 还能促进作物生长, 提高作物品质, 改善土壤质量, 环境友好, 有益于可持续发展等优点[12, 13].虽然有学者已报道使用植物促生细菌降低向日葵、油菜等对重金属的吸收[14, 15], 但是目前关于具有促进小麦生长和阻控小麦吸收重金属的功能细菌的分离筛选仍较少.此外, 有关分离自重金属污染土壤植物促生细菌与Cd和Pb相互关系以及Cd和Pb对菌株生长、代谢的影响等研究较少.因此, 本研究从Cd和Pb污染农田中采集狗尾草根际土壤, 依据重金属抗性和产吲哚乙酸(indole-3-acetic acid, IAA)、铁载体和1-氨基环丙烷羧酸(1-Amino-1-cyclopropanecarboxylic acid, ACC)脱氨酶的能力进行供试菌株的初筛; 采用静置培养实验, 研究菌株对溶液中Cd和Pb质量浓度、pH值和NH4+质量浓度的影响; 通过小麦砂培实验, 筛选到既能促进小麦生长又能阻控小麦吸收Cd和Pb的功能菌株, 以期为实施重金属污染农田生态整治与安全高效利用策略提供菌种资源和技术支撑.
1 材料与方法 1.1 土壤样品采集及理化性质分析于2018年5月在河南省新乡市环宇集团锂离子电池厂附近农田(35°3′N, 113°78′E)使用五点采样法[16], 采用抖落法采集狗尾草根际土壤样品.参考文献[17]测定土壤碱解氮、速效磷、速效钾、pH值和有机质含量等土壤理化性质.狗尾草根际土壤样品采用微波消解仪(SINEO MDS-6G, 中国)进行消解, 然后使用电感耦合等离子发射光谱仪(Inductively coupled plasma atomic emission spectrometry, ICP-OES, SHIMADZ ICPE-9820, 日本)测定Cd和Pb的质量浓度.
1.2 具有Cd和Pb抗性的植物促生细菌的筛选称取狗尾草根际土壤0.5 g放到含有50 mL无菌去离子水的摇瓶内, 150 r ·min-1, 振荡30 min, 然后取1 mL菌悬液梯度稀释成10-4和10-5, 涂布于含有400 mg ·L-1Cd和2 200 mg ·L-1Pb的LB固体平板上, 30℃培养3 d.从长出来的菌落中依据菌落大小、边缘形状和颜色等指标挑取菌落.用无菌牙签将菌株接种于含不同质量浓度Cd(400~1 000 mg ·L-1, 每隔50 mg ·L-1一个浓度梯度)和Pb(2 200~3 200 mg ·L-1, 每隔100 mg ·L-1一个浓度梯度)的LB固体培养基平板上, 30℃培养3 d, 若能长出单菌落, 转移到含有相应质量浓度重金属的LB液体培养基中, 培养18 h, 测定其D600.菌株产IAA的测定方法参考Jiang等[18]的方法, 产铁载体能力测定参考Rajkumar等[19]的方法, 产ACC脱氨酶能力测定方法参考Belimov等[20]的方法.
1.3 菌株吸附Cd和Pb能力测定配制含有Cd浓度为1 mg ·L-1、Pb浓度为4 mg ·L-1的LB液体培养基, 分装到三角瓶中, 每瓶50 mL.取活化后的菌悬液按1%比例接种到三角瓶中, 30℃、180 r ·min-1振荡培养48 h.培养结束后, 取5 mL发酵液、5 000 r ·min-1离心5 min, 然后取上清2 mL, 采用ICP-OES测定上清液中Cd和Pb的浓度.通过计算得出菌株对Cd和Pb的吸附率, 计算公式如下:
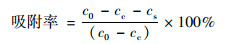
|
式中, c0表示溶液中重金属的初始浓度, mg ·L-1; cs表示供试菌株上清液中重金属浓度, mg ·L-1; cc表示对照样品上清液中重金属浓度, mg ·L-1.
1.4 菌株固定重金属静置培养实验参照1.3节的方法配制培养基.按1%的接种量将菌悬液接种于摇瓶中, 30℃静置培养, 设置不接菌作为对照.在第0、1、3、5和7 d取样.用分光光度计测定600 nm处的吸光度D600; 取7 mL发酵液, 12 000 r ·min-1离心5 min, 上清液用ICP-OES分析溶液中的Cd和Pb浓度; 用pH计(PHS-3CT)测定上清液的pH值; 参考Bai等[21]的方法, 采用靛酚蓝比色法测定溶液中NH4+的含量.
1.5 不同溶液pH对菌株固定吸附重金属的影响参照1.3节的方法配制培养基, 然后将培养基分成两份, 一份调节溶液pH为6.00, 另一份调节溶液pH为8.00.按1%的接种量将菌悬液接种于摇瓶中, 30℃, 180 r ·min-1振荡培养12 h, 同时设置不接菌处理作为对照.培养结束后, 取7 mL发酵液, 12 000 r ·min-1离心5 min, 上清液用ICP-OES分析溶液中Cd和Pb的质量浓度.
1.6 供试菌株种属鉴定和脲酶基因ureC扩增参考刘瑞等[22]的方法对菌株N3和H12进行种属鉴定, 然后利用MEGA6.0软件构建系统进化树.根据种属鉴定结果, 在NCBI(https://www.ncbi.nlm.nih.gov/)找到该种属的ureC基因序列, 利用CloneManager 8软件设计引物.菌株N3的ureC基因引物为ureC-3F:CGT TGC TGA CGC TGT TAA TC, ureC-3R: TTT GGC GCC GTT GAC GAT AC.菌株H12的ureC基因引物为ureC-12F:CTG GCG ATC TCC ACC TAT AC, ureC-12R: ACC GAG TGC CAC CCA GAT AG.然后以菌株DNA为模板进行PCR扩增, 最后送到生工生物工程(上海)股份有限公司进行序列测定.
1.7 小麦苗期砂培实验选取若干籽粒饱满的郑麦3号种子, 用75%乙醇进行消毒5 min, 再用无菌水漂洗3次, 然后置于水琼脂(0.4%)平板上, 30℃恒温培养, 待到种子长出两片小叶后, 挑取大小一致的小麦苗移栽于装有500 g无菌石英砂(间歇灭菌2次, 每次121℃, 40 min)的盆卜中进行培养, 盆卜上层装有石英砂, 下层则为霍格兰氏营养液.小麦幼苗长出5片叶子后, 添加重金属, 使Cd终浓度为1 mg ·L-1, Pb终浓度为4 mg ·L-1.采用菌悬液浸根2 h的方式对小麦进行接种, 同时以不接种处理作为对照.接菌完成后继续培养30 d.将培养完成后的小麦苗从水盆中取出, 根部用0.01 mol ·L-1EDTA-2Na溶液浸泡10 min, 以除去小麦根部表面吸附的重金属离子, 然后用剪刀将小麦分为地上部和根部, 置于烘箱80℃烘干, 测量小麦干重.将烘干的小麦样品用粉碎机磨碎, 准确称取0.5 g进行微波消解, 采用ICP-OES测定消解液中Pb和Cd的质量浓度.
1.8 数据处理采用SPSS 17.0软件进行数据统计分析, 用Microsoft Excel 2010软件绘图.
2 结果与分析 2.1 狗尾草根际土壤样品理化性质对所采集狗尾草根际土壤理化性质进行测定, 结果如表 1所示.狗尾草根际土壤pH值为7.03, 为中性土壤.总氮、总磷和总钾分别为5.3、2.5和5.46 g ·kg-1, 土壤有机质含量为22.71 g ·kg-1.土壤Cd含量为4.26 mg ·kg-1, Pb含量为323 mg ·kg-1, 根据《土壤环境质量标准》(GB 15618-1995), 狗尾草根际土壤Cd和Pb含量均达到三级污染标准, 为重度Cd和Pb污染土壤.
|
|
表 1 土壤样品主要理化参数1) Table 1 Main physical and chemical properties of the soil samples |
2.2 具有Cd和Pb抗性的植物促生细菌的分离筛选
从含有400 mg ·L-1 Cd和2 200 mg ·L-1Pb的LB固体平板上共分离到76株细菌, 然后测定了细菌的产IAA、铁载体和ACC脱氨酶的能力以及Cd和Pb的致死浓度.最终筛选到18株高产IAA、铁载体和ACC脱氨酶和耐受高浓度Cd和Pb的菌株, 结果如表 2所示.这18株菌株均能高产IAA, 介于18.49~69.14 mg ·L-1之间, 其中N3和H12产的IAA质量浓度最高, 分别为56.53 mg ·L-1和69.14 mg ·L-1.这18株菌株均能耐受高浓度的Cd (400~700 mg ·L-1)和Pb(2 400~2 700 mg ·L-1), 其中菌株N3的Cd致死浓度为700 mg ·L-1, Pb致死浓度为2 800 mg ·L-1; 菌株H12的Cd致死浓度为800 mg ·L-1, Pb致死浓度为2 800 mg ·L-1.
|
|
表 2 菌株的促生特性和重金属抗性1) Table 2 Growth promotion characteristics and heavy metal resistance of the isolated strains |
2.3 菌株吸附Cd和Pb能力比较
对初筛得到的18株具有促生能力的菌株进行Cd和Pb吸附能力的测定, 结果如图 1所示. 18株菌株对Cd的吸附率在18.6%~92.6%之间, 其中吸附率大于80%的菌株有2株, 分别为N3和H12. 18株菌株对Pb的吸附率在32.6%~91.5%之间, 其中吸附率大于80%的菌株有2株, 为N3和H12.由此可知, 菌株N3和H12具有很强的固定吸附Cd和Pb的能力.

|
图 1 菌株在溶液中对镉和铅的去除效果 Fig. 1 Efficiency of strains to remove Cd and Pb in liquid |
采用静置实验探究液体条件下供试菌株对Cd和Pb的吸附规律, 结果如图 2所示.随着培养时间的增加, 处理组N3和H12培养液的D600分别从0.02逐渐升高到2.02和2.2[图 2 (a)].对照组中Cd和Pb的质量浓度无显著差异, 均为0.88 mg ·L-1和3.65 mg ·L-1 [图 2(b)和2(c)], 但菌株N3和H12均能显著(P < 0.05)降低溶液中Cd(63.1%~73.8%)和Pb (69.1%~81.8%)的质量浓度[图 2(b)和2(c)].其中, 在培养第7 d, 处理组N3溶液中Cd和Pb的质量浓度分别为0.31 mg ·L-1和1.12 mg ·L-1, 处理组H12液中Cd和Pb的质量浓度分别为0.22 mg ·L-1和0.66 mg ·L-1.与对照相比, 处理组N3和H12溶液的pH值随着培养时间的增加而逐渐升高, 在第7 d, 处理组N3溶液中pH值达到8.02, 处理组H12溶液中pH值为8.41 [图 2 (d)].两株菌都能显著增加溶液中的NH4+质量浓度[图 2 (e)], 其中, 在培养第7 d, 处理组N3溶液NH4+的质量浓度为39.34 mg ·L-1, 处理组H12溶液中NH4+的质量浓度为46.21 mg ·L-1.

|
图 2 菌株N3和H12在溶液中生长变化及对溶液中Cd和Pb质量浓度、pH值和NH4+质量浓度的影响 Fig. 2 Changes in cell number (indicated by D600), Cd and Pb concentrations, pH, and NH4+ concentration in the culture solution in the presence of strains N3 and H12 |
采用人为设置不同溶液pH的方法探究pH对菌株固定吸附重金属的影响, 结果如图 3所示.在不接菌条件下, 溶液中Cd和Pb的质量浓度在溶液pH分别为6.00和8.00的条件下无显著变化.在接菌条件下, 溶液pH为8的溶液中Cd(0.54~0.61 mg ·L-1)和Pb(2.53~2.84 mg ·L-1)的质量浓度均显著小于溶液pH为6的溶液中Cd(0.73~0.78 mg ·L-1)和Pb(2.97~3.13 mg ·L-1)的质量浓度.

|
同一重金属条件下不同小写字母表示处理之间有显著差异(P < 0.05) 图 3 不同溶液pH对菌株固定吸附重金属的影响 Fig. 3 Effect of different pH solution on the immobilization Cd and Pb by strains |
以供试菌株N3和H12的总DNA为模板, 进行16S rDNA PCR扩增和测序, 在https://www.ezbiocloud.net/identify网站上进行序列比对, 利用MEGA6.0软件构建系统进化树, 结果如图 4所示.菌株N3为巨大芽孢杆菌(Bacillus megaterium), 登录号为MH715353, 菌株H12为液质沙雷氏菌(Serratia liquefaciens), 登录号为MH715354.由如图 5可知, 菌株N3和H12中均含有ureC基因, 这说明这两株供试菌株均含有脲酶基因, 能够分泌脲酶.

|
图 4 基于菌株16S rDNA序列构建的系统发育树 Fig. 4 Phylogenetic tree constructed by the Neighbor Joining method on the basis of 16S rDNA sequences of the strains |

|
图 5 菌株N3和H12的脲酶基因ureC扩增电泳图 Fig. 5 Validation electrophoregram of the ureC gene of strains N3 and H12 |
通过小麦苗期砂培实验, 研究供试菌株N3和H12对小麦生长和重金属吸收的影响, 结果如图 6和7所示.与不接菌对照相比, 接菌N3和H12处理显著促进小麦地上部和根部的干重(图 6).其中, 菌株N3使小麦地上部干重提高了65.3%, 根部干重提高了47.2%.菌株H12使小麦地上部干重提高了153%, 根部干重提高了97.4%.与不接菌对照相比, 菌株N3和H12均能够显著减少小麦地上部和根部Cd和Pb的含量(图 7).其中, 菌株N3使小麦根部Cd含量降低了49.2%, Pb含量降低了69.2%;地上部Cd含量降低了56.4%, Pb含量降低了27.4%.菌株H12使小麦根部Cd含量降低了68.3%, Pb含量降低了82.6%;地上部Cd含量降低了81.4%, Pb含量降低了84.2%.以上结果表明, 菌株N3和H12既能促进小麦生长又能阻控小麦富集Cd和Pb.

|
不同小写字母表示处理之间有显著差异(P < 0.05), 下同 图 6 供试菌株对小麦各组织干重的影响 Fig. 6 Effect of strains on the tissue dry weight of wheat |

|
图 7 供试菌株对小麦各组织镉和铅含量的影响 Fig. 7 Effect of the strains on the Cd and Pb concentrations in wheat |
据调查, 我国许多地区的农田土壤都受到重金属的污染, 尤其是Cd和Pb的污染十分严重[23].在这些土壤中种植农作物, 会造成农产品中重金属含量超标, 人类的健康受到严重的威胁.向土壤中施加羟基磷灰石、沸石、飞灰等化学钝化剂可以将重金属由生物有效性高的可交换态和碳酸盐结合态向生物有效性低的有机结合态和残渣态转化, 降低其生物有效性, 从而阻控作物吸收重金属[24, 25], 但是如果大量使用这些钝化修复剂, 会产生水体富营养化、重金属二次污染、妨碍微量营养元素吸收、影响土壤理化性质等不利影响, 并直接影响钝化修复剂的持续应用[26].近年来, 重金属固定植物促生细菌作为优良的重金属钝化剂被广泛研究和应用[16, 27].植物促生细菌对重金属的钝化作用可使重金属在土壤中沉淀或被吸附固定, 改变土壤重金属赋存状态, “钝化”其生物有效性, 从而降低作物对重金属的吸收[28]. Xu等[29]报道指出Bacillus subtilis 38能够耐受高浓度的Cd, 在Cd污染土壤中接种Bacillus subtilis 38, 能够阻控拟南芥对Cd的吸收, 显著降低了拟南芥中Cd的含量.本文在Cd污染地区植物根际土壤分离筛选获得了两株具有重金属固定能力的植物促生细菌苏云金芽孢杆菌N3和液质沙雷氏菌H12, 均能够显著降低小麦根和地上部Cd(49%~68%)和Pb(27%~84%)的含量.相比无机/有机钝化剂, 植物促生细菌最大的优势是不仅能够阻控作物吸收重金属, 还能够促进作物生长[1, 15].重金属污染土壤中根际植物促生细菌与植物根系以及土壤形成特殊根际微环境, 通过产生植物生长激素促进植物生长, 改变根际微环境中重金属元素生物有效性, 调控植物对重金属的吸收和富集效率[15, 30]. Egamberdieva等[31]报道细菌产生的吲哚乙酸能够促进盐胁迫下小麦幼苗根和茎的伸长.本文筛选的供试菌株N3和H12都能分泌大量的IAA、铁载体和ACC脱氨酶, 显著提高了小麦地上部(65%~153%)和根部的干量(47%~97%).
3.2 溶液条件下供试菌株固定钝化Cd和Pb规律研究植物促生细菌能够通过吸附重金属离子降低其生物有效性或降低溶液中重金属的浓度[32]. pH值是影响重金属在环境中分布、转化以及生物有效态含量高低的最关键因素[33].溶液pH升高时, 即溶液中的H+显著减小.由于细菌的细胞壁上含有许多阴离子官能团, 当溶液中的H+减少时, 与阴离子官能团结合的H+也会减少.由于重金属带有正电荷, 便会结合到阴离子官能团上, 从而增加细菌细胞壁对重金属的吸附量, 降低溶液中重金属的含量[34]. Xu等[35]的研究表明, Enterobacter cloacae TU对Cd的吸附率高达97%, 吸附的Cd主要位于胞外, 其产生的胞外聚合物中的O—H、N—H和S—H是Cd结合的主要位点.本文筛选得到的两株植物促生细菌N3和H12在Cd和Pb浓度分为1 mg ·L-1和4 mg ·L-1的LB液体培养基中都能吸附大量的Cd和Pb, 对Cd和Pb的吸附率都达到80%以上.
本研究发现, 随着培养时间的增加, 菌株N3和H12都能显著降低溶液中Cd和Pb浓度的同时, 均能显著提高溶液的pH值, 在培养第7 d, 菌株H12处理组溶液中pH值为8.48, 菌株N3处理组溶液中pH值为8.08.这说明两株供试菌株在生长过程中通过提高溶液的pH值, 来固定Cd和Pb, 从而缓解了Cd和Pb的胁迫.进一步测定发现功能菌株N3和H12均含有脲酶基因, 能够分泌脲酶.而细菌脲酶可以催化尿素生成NH4+, 而NH4+质量浓度的增加则会提高周围环境的pH值[35].细菌脲酶由结构基因簇、辅助基因簇和调节基因簇等三大基因簇组成, 大多数细菌脲酶的结构亚基基因包括ureA、ureB和ureC, 其中ureC可以被选作脲酶活性分析的靶基因, 因为它是编码尿素酶功能亚基的最大的基因, 并包含若干个高度保守的区域, 所以ureC比较适合作为分子标记来设计引物, 进行PCR扩增[36]. Fisher等[37]报道土壤中ureC基因丰度的提高可以提高土壤中的pH值. Yang等[38]报道指出含有N循环基因(ureC、gdh和amoA)的细菌种群与土壤中的N的新陈代谢密切相关, 且能够提高土壤中的pH值和NH4+含量.通过对菌株N3和H12进行PCR扩增, 以及凝胶电泳观察和序列比对, 确定两株功能菌株均含有脲酶基因, 在生长过程中均能分泌大量的脲酶, 提高NH4+的质量浓度, 进而提高了溶液的pH值.
4 结论(1) 本研究从Cd和Pb污染农田中分离筛选了两株重金属固定植物促生细菌巨大芽孢杆菌N3和液质沙雷氏菌H12, 均能够耐受高浓度的Cd和Pb, 分泌IAA、铁载体和ACC脱氨酶.
(2) 菌株N3和H12含有脲酶合成基因ureC, 能分泌脲酶, 显著提高溶液的pH值和NH4+的质量浓度, 从而降低溶液中Cd和Pb的质量浓度.
(3) 砂培条件下, 菌株N3和H12不仅能够显著增加小麦根部和地上部的干重, 还能够显著降低小麦根部和地上部中Cd和Pb的含量.
| [1] | Chen L, He L Y, Wang Q, et al. Synergistic effects of plant growth-promoting Neorhizobium huautlense T1-17 and immobilizers on the growth and heavy metal accumulation of edible tissues of hot pepper[J]. Journal of Hazardous Materials, 2016, 312: 123-131. DOI:10.1016/j.jhazmat.2016.03.042 |
| [2] | Wu C, Shi L Z, Xue S G, et al. Effect of sulfur-iron modified biochar on the available cadmium and bacterial community structure in contaminated soils[J]. Science of the Total Environment, 2019, 647: 1158-1168. DOI:10.1016/j.scitotenv.2018.08.087 |
| [3] |
靳琪, 高红, 岳波, 等. 村镇生活垃圾重金属含量及其来源分析[J]. 环境科学, 2018, 39(9): 4385-4392. Jin Q, Gao H, Yue B, et al. Heavy metal content of rural living solid waste and related source and distribution analysis[J]. Environmental Science, 2018, 39(9): 4385-4392. |
| [4] |
陈能场, 郑煜基, 何晓峰, 等. 《全国土壤污染状况调查公报》探析[J]. 农业环境科学学报, 2017, 36(9): 1689-1692. Chen N C, Zheng Y J, He X F, et al. Analysis of the Report on the national general survey of soil contamination[J]. Journal of Agro-Environment Science, 2017, 36(9): 1689-1692. |
| [5] | Mackie K A, Marhan S, Ditterich F, et al. The effects of biochar and compost amendments on copper immobilization and soil microorganisms in a temperate vineyard[J]. Agriculture, Ecosystems & Environment, 2015, 201: 58-69. |
| [6] | Wan Y N, Camara A Y, Yu Y, et al. Cadmium dynamics in soil pore water and uptake by rice:Influences of soil-applied selenite with different water managements[J]. Environmental Pollution, 2018, 240: 523-533. DOI:10.1016/j.envpol.2018.04.044 |
| [7] |
赵军超, 王权, 任秀娜, 等. 钙基膨润土辅助对堆肥及土壤Cu、Zn形态转化和白菜吸收的影响[J]. 环境科学, 2018, 39(4): 1926-1933. Zhao J C, Wang Q, Ren X N, et al. Effect of Ca-bentonite on Cu and Zn forms in compost and soil, and their absorption by Chinese cabbage[J]. Environmental Science, 2018, 39(4): 1926-1933. |
| [8] |
曾希柏, 徐建明, 黄巧云, 等. 中国农田重金属问题的若干思考[J]. 土壤学报, 2013, 50(1): 186-194. Zeng X B, Xu J M, Huang Q Y, et al. Some deliberations on the issues of heavy metals in farmlands of China[J]. Acta Pedologica Sinica, 2013, 50(1): 186-1954. |
| [9] |
韩蕾, 陈娟, 杜平, 等. 不同钝化剂对Cd污染农田土壤生态安全的影响[J]. 环境科学研究, 2018, 31(7): 1289-1295. Han L, Chen J, Du P, et al. Assessing the ecological security of the cadmium contaminated farmland treated with different amendments[J]. Research of Environmental Science, 2018, 31(7): 1289-1295. |
| [10] |
余斐, 苏艳, 李吉跃, 等. 植物根际促生菌促生机理研究[J]. 林业与环境科学, 2017, 33(2): 107-112. Yu F, Su Y, Li J Y, et al. Plant growth-promoting rhizobacteria promoting mechanism research[J]. Forestry and Environmental Science, 2017, 33(2): 107-112. DOI:10.3969/j.issn.1006-4427.2017.02.020 |
| [11] | Sharma R K, Archana G. Cadmium minimization in food crops by cadmium resistant plant growth promoting rhizobacteria[J]. Applied Soil Ecology, 2016, 107: 66-78. DOI:10.1016/j.apsoil.2016.05.009 |
| [12] | Lin X Y, Mou R X, Cao Z Y, et al. Characterization of cadmium-resistant bacteria and their potential for reducing accumulation of cadmium in rice grains[J]. Science of the Total Environment, 2016, 569-570: 97-104. DOI:10.1016/j.scitotenv.2016.06.121 |
| [13] | Ahmad M, Ok Y S, Rajapaksha A U, et al. Lead and copper immobilization in a shooting range soil using soybean stover- and pine needle-derived biochars:Chemical, microbial and spectroscopic assessments[J]. Journal of Hazardous Materials, 2016, 301: 179-186. DOI:10.1016/j.jhazmat.2015.08.029 |
| [14] | Marques A P G C, Moreira H, Franco A R, et al. Inoculating Helianthus annuus (sunflower) grown in zinc and cadmium contaminated soils with plant growth promoting bacteria-Effects on phytoremediation strategies[J]. Chemosphere, 2013, 92(1): 74-83. DOI:10.1016/j.chemosphere.2013.02.055 |
| [15] | Han H, Wang Q, He L Y, et al. Increased biomass and reduced rapeseed Cd accumulation of oilseed rape in the presence of Cd-immobilizing and polyamine-producing bacteria[J]. Journal of Hazardous Materials, 2018, 353: 280-289. DOI:10.1016/j.jhazmat.2018.04.024 |
| [16] | Chang B V, Yu C H, Yuan S Y. Degradation of nonylphenol by anaerobic microorganisms from river sediment[J]. Chemosphere, 2004, 55(4): 493-500. DOI:10.1016/j.chemosphere.2004.01.004 |
| [17] | 鲍士旦. 土壤农化分析[M]. (第三版).北京: 中国农业出版社, 2000: 86-115. |
| [18] | Jiang C Y, Sheng X F, Qian M, et al. Isolation and characterization of a heavy metal-resistant Burkholderia sp. from heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal-polluted soil[J]. Chemosphere, 2008, 72(2): 157-164. DOI:10.1016/j.chemosphere.2008.02.006 |
| [19] | Rajkumar M, Nagendran R, Lee K J, et al. Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard[J]. Chemosphere, 2006, 62(5): 741-748. DOI:10.1016/j.chemosphere.2005.04.117 |
| [20] | Belimov A A, Hontzeas N, Safronova V I, et al. Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.)[J]. Soil Biology and Biochemistry, 2005, 37(2): 241-250. DOI:10.1016/j.soilbio.2004.07.033 |
| [21] | Bai J H, Wang Q G, Gao H F, et al. Spatial and temporal distribution patterns of nitrogen in marsh soils from an inland alkaline wetland-A case study of Fulaowenpao wetland, China[J]. Acta Ecologica Sinica, 2010, 30(4): 210-215. DOI:10.1016/j.chnaes.2010.06.004 |
| [22] |
刘瑞, 于章龙, 薛冲, 等. 市售豆芽携带细菌种属鉴定及酸性电解水的杀菌效果[J]. 食品科学, 2017, 38(17): 168-173. Liu R, Yu Z L, Xue C, et al. Identification of bacterial species and microbial inactivation by acidic electrolyzed water on commercial bean sprouts[J]. Food Science, 2017, 38(17): 168-173. DOI:10.7506/spkx1002-6630-201717028 |
| [23] |
龚梦丹, 朱维琴, 顾燕青, 等. 杭州蔬菜基地重金属污染及风险评价[J]. 环境科学, 2016, 37(6): 2329-2337. Gong M D, Zhu W Q, Gu Y Q, et al. Evaluation on heavy metal pollution and its risk in soils from vegetable bases of Hangzhou[J]. Environmental Science, 2016, 37(6): 2329-2337. |
| [24] | Wang D F, Zhang G L, Zhou L L, et al. Immobilizing arsenic and copper ions in manure using a nanocomposite[J]. Journal of Agricultural and Food Chemistry, 2017, 65(41): 8999-9005. DOI:10.1021/acs.jafc.7b02370 |
| [25] |
刘德玲, 尹光彩, 陈志良, 等. 硅酸钙和生物腐殖肥复配对葱生长和镉吸收的影响[J]. 环境科学, 2018, 39(6): 2927-2935. Liu D L, Yin G C, Chen Z L, et al. Effect of calcium silicate-biological humus fertilizer composite on uptake of Cd by shallots from contaminated agricultural soil[J]. Environmental Science, 2018, 39(6): 2927-2935. |
| [26] |
王欣, 尹带霞, 张凤, 等. 生物炭对土壤肥力与环境质量的影响机制与风险解析[J]. 农业工程学报, 2015, 31(4): 248-257. Wang X, Yin D X, Zhang F, et al. Analysis of effect mechanism and risk of biochar on soil fertility and environmental quality[J]. Transactions of the Chinese Society of Agricultural Engineering, 2015, 31(4): 248-257. DOI:10.3969/j.issn.1002-6819.2015.04.035 |
| [27] | Ma Y, Prasad M N V, Rajkumar M, et al. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils[J]. Biotechnology Advances, 2011, 29(2): 248-258. DOI:10.1016/j.biotechadv.2010.12.001 |
| [28] | Etesami H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues:Mechanisms and future prospects[J]. Ecotoxicology and Environmental Safety, 2018, 147: 175-191. DOI:10.1016/j.ecoenv.2017.08.032 |
| [29] | Xu Q R, Pan W, Zhang R R, et al. Inoculation with Bacillus subtilis and Azospirillum brasilense produces abscisic acid that reduces Irt1-mediated Cadmium uptake of roots[J]. Journal of Agricultural and Food Chemistry, 2018, 66(20): 5229-5236. DOI:10.1021/acs.jafc.8b00598 |
| [30] | Babu A G, Shea P J, Sudhakar D, et al. Potential use of Pseudomonas koreensis AGB-1 in association with Miscanthus sinensis to remediate heavy metal(loid)-contaminated mining site soil[J]. Journal of Environmental Management, 2015, 151: 160-166. |
| [31] | Egamberdieva D. Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat[J]. Acta Physiologiae Plantarum, 2009, 31(4): 861-864. DOI:10.1007/s11738-009-0297-0 |
| [32] | Bai J, Yang X H, Du R Y, et al. Biosorption mechanisms involved in immobilization of soil Pb by Bacillus subtilis DBM in a multi-metal-contaminated soil[J]. Journal of Environmental Sciences, 2014, 26(10): 2056-2064. DOI:10.1016/j.jes.2014.07.015 |
| [33] |
陈远其, 张煜, 陈国梁. 石灰对土壤重金属污染修复研究进展[J]. 生态环境学报, 2016, 25(8): 1419-1424. Chen Y Q, Zhang Y, Chen G L. Remediation of heavy metal contaminated soils by lime:a review[J]. Ecology and Environmental Sciences, 2016, 25(8): 1419-1424. |
| [34] | Huang J H, Yuan F, Zeng G M, et al. Influence of pH on heavy metal speciation and removal from wastewater using micellar-enhanced ultrafiltration[J]. Chemosphere, 2017, 173: 199-206. DOI:10.1016/j.chemosphere.2016.12.137 |
| [35] | Xu C R, He S B, Liu Y M, et al. Bioadsorption and biostabilization of cadmium by Enterobacter cloacae TU[J]. Chemosphere, 2017, 173: 622-629. DOI:10.1016/j.chemosphere.2017.01.005 |
| [36] | Gresham T L T, Sheridan P P, Watwood M E, et al. Design and validation of ureC-based primers for groundwater detection of urea-hydrolyzing bacteria[J]. Geomicrobiology Journal, 2007, 24(3-4): 353-364. DOI:10.1080/01490450701459283 |
| [37] | Fisher K A, Yarwood S A, James B R. Soil urease activity and bacterial ureC gene copy numbers:effect of pH[J]. Geoderma, 2017, 285: 1-8. DOI:10.1016/j.geoderma.2016.09.012 |
| [38] | Yang Y F, Gao Y, Wang S P, et al. The microbial gene diversity along an elevation gradient of the Tibetan grassland[J]. The ISME Journal, 2014, 8(2): 430-440. DOI:10.1038/ismej.2013.146 |