 2019, Vol. 40
2019, Vol. 40


2. 合肥工业大学土木与水利工程学院, 合肥 230009
2. College of Civil Engineering, Hefei University of Technology, Hefei 230009, China
磷是包括微生物在内的所有生命体中不可缺少的营养元素, 并主要以磷酸盐形式贮存于沉积物中, 因此沉积物在磷的生物地球化学循环中扮演重要角色[1, 2], 对调控水体磷浓度具有重要的环境生态意义.一些研究证实, 沉积物中微生物对磷的吸收、贮存和释放在磷循环中起着极其重要的作用[3~6].近年来, 针对水底固着或悬浮颗粒生物磷吸收的研究逐渐增多[7~9], 对推动微生物技术在水污染治理和水环境修复领域的应用发挥了积极作用.截至目前, 有关开放水体微生物磷吸收的作用机制, 特别是生物与非生物因素在磷吸收中的相对贡献情况依旧不是很清楚, 如何通过技术调控强化沉积物固磷和降低磷释放风险, 也还缺乏成熟的技术方法.投放经培养筛选的微生物菌种(如光合细菌PSB、高效微生物菌群EM等)可以在一定程度上改善和调控河流水质, 但却存在潜在生态风险, 而且污染治理成本较高, 大规模推广应用有一定难度.为此, 河流底栖固着或悬浮颗粒物上土著微生物、藻类等的磷吸收作用引起人们的兴趣[10~13], 其对外源营养物添加的响应受到人们的关注[14].但由于河流水体类型和环境水文条件的多样性, 相关研究还显得很不充分, 远不能满足河流水环境治理工作的需要.
巢湖是水体富营养化较为严重的湖泊[15], 目前正面临入湖氮磷负荷削减的艰巨任务[16].十五里河是合肥市西南城区与巢湖西半湖的重要联系纽带, 是巢湖西半湖外源氮、磷负荷输入的主要通道之一[17].十五里河上游原有的两个化肥生产企业已分别于2008年和2012年关停、搬迁, 但长期污水排放在河道底部富集的氮磷营养物质并未得到有效处置, 并通过内源释放形式不断影响河流水质, 导致河流氮、磷水质指标长期处于劣Ⅴ类水平[17], 既严重影响了合肥市城市形象, 也加大了巢湖富营养化治理的难度.目前, 如何有效改善十五里河水质状况、削减向下游巢湖传输的磷负荷, 成为困扰合肥市政府的一大难题.本研究选择十五里河为对象, 尝试从河流自身磷净化能力提升角度, 解析十五里河沉积物磷吸收潜力, 并对其对于不同外源碳的响应效应开展定量化分析, 以期为十五里河水环境治理和磷负荷削减提供依据.
1 研究区概况十五里河发源于合肥市大蜀山东南麓, 地处东经116°40′~117°52′, 北纬31°30′~32°37′, 全长35 km, 河流形态曲折变化明显, 为丘陵地区小河流水体.十五里河自西北向东南流经合肥市的蜀山区和包河区, 在义城镇附近汇入巢湖.河流上游人工开挖形成的天鹅湖水体是当地重要的景观水体, 建有溢流坝, 坝下主河道长22.64 km, 流域面积约111.25 km2.汇流区地形较为平缓, 丘陵岗地和河湖低洼平原为主要地貌类型; 气候温和湿润, 年均气温15~19℃, 平均湿度77%, 平均降雨量为1 000 mm.
十五里河上游地区为近十多年来新建的合肥市政务中心和高档居住小区, 该处附近原有的江淮化肥厂和红四方化工集团等两个化肥生产企业也已在近十年陆续停产、搬迁; 中游为城市在建区, 主要用于商品住宅和工业园区建设, 下游东南部为城郊结合部, 目前土地基本都划归巢湖北岸生态湿地保护范畴.十五里河是巢湖流域水质污染最严重的河流水体[18], 本研究期间河流水质情况见表 1.
|
|
表 1 河流水质统计结果 Table 1 Mean water quality indexes of the Shiwuli River |
2 材料与方法 2.1 采样点布设与样品预处理
根据河流形态、排污口或支流汇入情况以及周边土地利用特征, 沿十五里河干流的城乡梯度方向布设5个采样点位, 分别位于天鹅湖坝下(采样点1)、南门换乘中心附近的梳背桥下游(采样点2)、徽州大道的高王桥上游(采样点3)、包河大道包河桥上游(采样点4)和巢湖入湖口上游1 km的义城镇附近(采样点5).
2017年7月(夏季)和2018年1月(冬季), 在每个采样点的0.5 m2范围内, 采集3个表层沉积物样, 并均匀混合代表该点位.将采集的沉积物迅速送回实验室冷藏, 并对每个采样点取一部分新鲜沉积物用于磷吸收潜力估算, 其余部分放置在阴凉处自然风干、研磨, 过80目筛后用于分析沉积物理化性质.
2.2 实验方案设计(1) 培养前沉积物磷含量的测定针对每个采样点, 按是否滴入抑制剂设置2种情景, 并分别取3个平行.首先, 向6个100 mL离心管中, 分别移入4 g左右的新鲜沉积物; 然后, 分别加入40 mL磷提取液(0.1mol·L-1 NaOH、0.1 mol·L-1 NaCl), 并向其中3个试管中另滴入3~4滴饱和氯仿溶液作为抑制剂.将离心管加塞后放置于25℃、200 r·min-1的振荡器中振荡16 h后, 再置于3 000 r·min-1离心机中离心5 min.取上清液2 mL, 将其注入50 mL比色管, 定容到标线刻度, 并滴加1 mL抗坏血酸(100 g·L-1)和2 mL钼酸盐溶液(26 g·L-1), 振荡摇匀, 静置15 min, 测定上清液磷酸盐浓度.对3个平行取均值, 再由溶液体积换算出新鲜沉积物的磷含量, 即SRPinitial.其中, 滴入氯仿的磷含量表示为SRPkillinitial, 未滴加氯仿的表示为SRPliveinitial.
(2) 培养后沉积物磷含量的测定针对每个采样点沉积物均设置8种实验方案, 另有空白实验作为对照, 共9种方案(表 2), 每种方案取3个平行.首先, 向每个100 mL离心管中, 分别移入4 g左右的新鲜沉积物.除空白实验仅加入40 mL蒸馏水外, 其它8种方案均加入20 mL培养液(1 mg·L-1 PO43--P、50 mg·L-1CaCl2溶液和30 mg·L-1 MgCl2溶液; 其中PO43--P溶液是由KH2PO4磷酸二氢钾配制).这里, 方案1和方案2加入了20 mL蒸馏水, 方案3和方案4加入了20 mL 100 mg·L-1乙酸钠(以C计), 方案5和方案6加入了20 mL 100 mg·L-1葡萄糖(以C计), 方案7和方案8加入了20 mL 100 mg·L-1乙酸钠和葡萄糖混合溶液(两者按1:1体积比混合, 以C计).其中, 方案2、4、6和8中均滴入了3~4滴饱和氯仿溶液.
|
|
表 2 实验方案设计1) Table 2 Design of the experimental scheme |
将所有离心管加塞并盖紧后置于30℃恒温培养箱中静置培养24 h.培养结束后, 再向各离心管中加入40 mL磷提取液(同上), 并盖塞振荡、离心, 测定上清液的磷酸盐浓度.对3个平行取平均值, 再根据溶液体积, 换算出沉积物可提取的磷含量SRPfinal(扣除了培养液引入的磷量).将滴入氯仿的磷含量表示为SRPkillfinal, 未滴加氯仿的表示为SRPlivefinal.将沉积物底质样过滤, 并将滤纸与沉积物一起置于烘箱中烘至恒重, 确定沉积物烘干后的质量dw(事先将滤纸置于烘箱中烘至恒重并记录质量).
2.3 样品分析测试沉积物样品中TN采用过硫酸钾消解法测定; TP采用钼锑抗比色法进行测定; Ex-P采用EDTA提取, 抗坏血酸法测定; Fe/Al-P按照SMT法测定; 有机质(OM)含量采用烧失量估算(105℃烘箱烘干12 h, 然后在550℃马弗炉中灼烧5 h); pH值采用pH计测定(水土比为5:1).至于培养前后沉积物上清液的磷酸盐浓度, 采用分光光度计(700 nm吸光度)测定.
2.4 磷吸收潜力估算沉积物磷的吸收潜力根据式(1)、(2)计算[19, 20]:
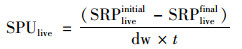
|
(1) |

|
(2) |
式中, SPUlive和SPUkill分别表示沉积物磷的总吸收潜力和非生物吸收潜力, μg·(g·h)-1; SRPliveinitial和SRPkillinitial分别表示培养前未滴入和滴入氯仿提取的磷含量, μg; SRPlivefinal和SRPkillfinal分别表示培养后未滴入和滴入氯仿提取的磷含量, μg; dw表示沉积物干质量, g; t表示培养时间, h.
由于SPUlive是生物与非生物吸收共同作用的结果, 而SPUkill反映了非生物吸收效应(也即物理化学吸附作用), 因此两者之差可以体现磷的生物吸收潜力, 不妨记作SPUbiotic, 则有:
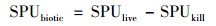
|
(3) |
采用Excel软件完成数据处理和绘制图表.采用Mann-Whitney U检验法分析研究对象的差异性, P<0.05表示样本差异性显著, P<0.01表示样本差异性极显著.
3 结果与讨论 3.1 沉积物基本理化特性5个采样点夏季和冬季表层沉积物的磷形态及其理化性质, 见表 3.可以看出, 十五里河沉积物氮磷污染相当严重, 氮磷含量显著高于一般河流、湖库水体.而与氮磷含量相比, 绝大多数采样点位沉积物的有机质含量却并不突出, 这可能与该河沉积物中的氮磷主要来自于上游的两个化肥厂污水大量排放造成的污染富集有关.由生态化学计量学原理, 微生物在对氮、磷、碳等生源物质的吸收利用上存在一定的化学计量关系[21, 22].由此, 大致可以推断, 十五里河沉积物中相对较不充分的有机质含量可能会对磷循环的生物地球化学过程产生一定的限制效应.
|
|
表 3 沉积物磷形态及其基本理化特性 Table 3 Phosphorus forms in sediment and its basic physical and chemical properties |
总体上, 5个采样点Ex-P和TN都表现为冬季含量明显高于夏季, 前3个采样点的Fe/Al-P和TP冬季含量高于夏季, 而IP则恰好相反. OM除采样点1外, 都表现为夏季含量高于冬季. 5个采样点的沉积物氮磷含量存在明显的差异性, 但并没有表现出显著的城乡梯度特征.
3.2 磷吸收潜力的外源碳影响 3.2.1 磷的总吸收不同外加碳情形下沉积物磷的总吸收潜力情况, 见图 1.可以看出, 十五里河沉积物磷的总吸收潜力存在一定的时空差异性.总体上, 夏季时采样点1与采样点5的SPUlive基本相当, 且从上游至下游, SPUlive逐步增大, 并于采样点4达到最大值, 然后迅速下降; 而冬季时采样点1的SPUlive明显高于其它4个采样点, 且采样点2至采样点4的SPUlive基本相当, 略高于采样点5.此外, 采样点2和采样点5的夏、冬季节差异并不明显, 采样点1表现出冬季显著大于夏季, 而采样点3和4则为夏季明显高于冬季.十五里河中下游地区人为活动较为显著, 表层沉积物的组成和性质易受人为扰动的影响.采样过程中发现, 冬季时的采样点1、夏季时的采样点4表层沉积物粒度都明显较同时期其他4个采样点更为细小一些.而且, 夏季时无论是TP还是生物可利用性磷(包括Ex-P和Fe/Al-P)含量, 采样点4都明显最高, 冬季时采样点1也表现出TP和生物可利用性磷(特别是Fe/Al-P)含量明显高于其他采样点的变化特征(表 3), 这可能是上述两个采样点SPUlive异常偏高的主要原因.

|
图 1 各采样点沉积物磷的总吸收潜力 Fig. 1 Total potential for phosphorus uptake by sediments for each sampling site |
与空白样相比, 其它4个方案的SPUlive都有不同程度的增大, 意味着沉积物表现出了磷吸收特征.而且, 无论是夏季还是冬季, 各采样点沉积物磷的总吸收对外加碳的响应规律相似, 即添加乙酸钠的沉积物SPUlive最高, 葡萄糖次之, 乙酸钠和葡萄糖混合溶液的最低, 但高于仅添加磷培养液的方案1.由于外源碳添加与否对应的SPUlive数值相差不大, 表明外源碳输入对于沉积物磷的总吸收影响不大.
在方案1中, 夏季各采样点的SPUlive变化范围为1.950~8.489 μg·(g·h)-1, 均值为3.800 μg·(g·h)-1, 冬季变化范围为1.770~6.853 μg·(g·h)-1, 均值为3.691 μg·(g·h)-1.李如忠等[20]采用该方案对合肥城郊一条受人为扰动影响较大的溪流进行研究, 得到夏、秋季节SPUlive变化范围为1.27~9.10 μg·(g·h)-1, 均值为3.143 μg·(g·h)-1, 与本研究相近. Lottig等[19]解析了不同底质类型溪流沉积物的磷吸收特征, 发现沙质河床沉积物SPUlive平均值为23.0 μg·(g·h)-1, 砾石河床为14.5 μg·(g·h)-1, 岩石河床为12.5 μg·(g·h)-1, 高于十五里河沉积物.
采用Mann-Whitney U检验法对十五里河沉积物总吸收潜力进行差异性分析, 结果发现:采样点2和3、采样点2和4、采样点3和5、采样点4和5存在极显著差异性(P<0.01), 其它采样点之间差异性则不显著(P>0.082);不仅每个采样点位中各方案之间差异性不显著(P>0.151), 而且整个河流夏、冬两季的总吸收潜力差异性也不显著(P=0.594).
3.2.2 磷的非生物吸收不同外源碳添加情形的磷的非生物吸收潜力SPUkill情况见图 2.其中, 未添加碳源(即仅添加含磷培养液)的夏季SPUkill变化范围为1.190~7.378 μg·(g·h)-1, 均值为3.016 μg·(g·h)-1; 冬季变化范围为1.643~5.631 μg·(g·h)-1, 均值为3.368 μg·(g·h)-1, 也就是说, 冬季非生物吸收潜力略高于夏季.李如忠等[20]在合肥城郊人为扰动较大溪流的研究中, 得到夏、秋两季的SPUkill变化范围为1.19~7.44 μg·(g·h)-1, 均值为2.567 μg·(g·h)-1, 与本研究较为接近. Lottig等[19]得到沙质河床沉积物的SPUkill均值为24.0 μg·(g·h)-1, 砾石河床为9.8 μg·(g·h)-1, 岩石河床为5.0 μg·(g·h)-1, 稍高于十五里河沉积物.

|
图 2 各采样点位沉积物磷的非生物吸收潜力变化 Fig. 2 Variations in the abiotic uptake potential for phosphorus by sediments for each sampling site |
总体上, 沿城乡梯度方向5个采样点SPUkill变化情况与SPUlive大体相似, 即夏季时采样点4的SPUkill显著高于其他点位, 冬季时采样点1也明显高于其他采样点, 这可能与两个采样点表层沉积物的粒度更为细小, 以致对磷吸附能力更强有关[7].除采样点4的夏季SPUkill明显高于冬季外, 其他4个采样点几乎都较冬季低.而且, 无论是夏季还是冬季, 外源碳添加与否对SPUkill影响很小, 即添加碳源对非生物吸收潜力几乎没有明显的促进作用.
非生物吸收潜力的差异性分析结果表明, 采样点1和3、采样点1和4、采样点2和3、采样点3和5存在极显著性差异性(P<0.01), 其它采样点之间差异性不显著(P>0.074);各采样点各种方案之间差异性也不显著(P>0.257);夏、冬两季的非生物吸收潜力存在显著差异性(P=0.026).
3.2.3 磷的生物吸收5个采样点位沉积物磷的生物吸收潜力情况见图 3.显然, 夏季时采样点3的SPUbiotic明显高于其它采样点, 冬季时采样点1相对较高, 且在添加乙酸钠、葡萄糖时与采样点2接近.总体上, 夏季磷的生物吸收潜力高于冬季, 特别是中下游采样点.就未添加外源碳而言, 除采样点1和采样点5的夏季生物吸收潜力略低于冬季外, 其它采样点都表现为夏季高于冬季, 特别是位于城市在建区的采样点3和采样点4. 5个采样点的夏季SPUbiotic变化范围为0.072~1.710 μg·(g·h)-1, 均值为0.784 μg·(g·h)-1; 冬季变化范围为0.126~0.951 μg·(g·h)-1, 均值为0.323 μg·(g·h)-1, 与文献[20]对合肥城郊溪流沉积物SPUbiotic的估算结果相近.

|
图 3 各采样点位沉积物磷的生物吸收潜力变化情况 Fig. 3 Variations in the biotic uptake potential of phosphorus by sediments for each sampling site |
显然, 在外加碳源的情形下, 5个采样点的生物吸收潜力都表现出了相似的变化特征, 即添加乙酸钠的SPUbiotic最高, 添加葡萄糖次之, 投加混合溶液的SPUbiotic最低.尽管添加碳源基本都较不添加碳源的生物吸收潜力更高, 但乙酸钠和葡萄糖混合溶液对沉积物的生物吸收潜力提升幅度并不大, 而且夏、冬季都如此.
采样点1和采样点5之间存在极显著差异性(P=0.005), 采样点3和采样点5、采样点4和采样点5之间则都存在显著差异性(P<0.05), 其它采样点之间的差异性不明显(P>0.083). 5个采样点中, 未添加碳源和添加乙酸钠情形之间存在极显著差异性(P=0.010), 添加乙酸钠和添加混合碳源情形之间存在显著差异性(P=0.019), 而其它各组方案之间差异性则不显著(P>0.059).总体上, 夏、冬两季节之间磷的生物吸收潜力接近显著差异性水平(P=0.055).
不妨采用(SPUbioticC+P-SPUbioticP)/SPUbioticP×100%定量刻画磷的生物吸收潜力对于外加碳源的响应效果, 其中SPUbioticC+P表示添加了磷培养液和外源碳的生物吸收潜力, μg·(g·h)-1; SPUbioticP表示仅添加了磷培养液的生物吸收潜力, μg·(g·h)-1. 5个采样点沉积物磷的生物吸收潜力对外源碳的响应情况, 见表 4.显然, 生物吸收潜力对乙酸钠的响应相对最为显著, 其次是葡萄糖, 而对于混合碳源的响应最弱.而且, 大多数采样点冬季响应水平高于夏季.可以说, 十五里河大多数采样点位沉积物中磷的生物吸收潜力对外源碳的响应效应, 特别是对乙酸钠的偏好性, 表明该河沉积物磷的生物吸收存在一定程度的碳限制性.
|
|
表 4 外源碳添加对磷的生物吸收潜力的影响水平/% Table 4 Influence of carbon additions on the biotic uptake potential of phosphorus/% |
3.3 生物非生物吸收贡献率
不同外加碳源情形下生物与非生物因素的磷吸收贡献情况见图 4.在未添加外源碳的情况下, 5个采样点夏季的生物与非生物吸收贡献率范围分别为3.41%~39.77%(均值为21.55%)和60.23%~96.59%(均值为78.45%), 冬季分别为4.58%~14.45%(均值为7.48%)和85.55%~95.42%(均值为92.52%), 即十五里河沉积物磷吸收主要来自非生物因素贡献, 尤其是冬季.但采样点1和采样点3的夏季生物贡献率仍高达40%左右, 生物因素影响较为显著.李如忠等[20]在对合肥城郊溪流研究中, 得到生物与非生物吸收贡献率分别为3.98%~35.21%、64.79%~96.02%, 与本研究非常接近. Aldridge等[23]针对岩石附生生物群落的实验研究发现, 磷的生物吸收贡献率不足30%, 而非生物吸收贡献率达70%以上. Stutter等[6]在对以藻类和细菌为主体的水底生物膜研究中, 得到磷的生物吸收率为37.5%、非生物吸收率为62.5%.但也有学者发现生物吸收贡献率占据主导地位, 如Lottig等[19]发现在沉积物粒度较粗的溪流磷滞留中, 约50%以上的磷吸收都来自于生物作用过程.

|
图 4 不同季节各采样点磷的生物与非生物吸收贡献率变化 Fig. 4 Changes in the contribution rates of biotic and abiotic uptake of phosphorus at each sampling site in different seasons |
在添加了乙酸钠后, 5个采样点的夏季和冬季沉积物磷的生物吸收贡献率, 分别达20.31%~55.09%(均值为42.40%)和16.48%~42.83%(均值为23.99%); 添加葡萄糖后, 夏季和冬季的生物贡献率分别为14.05%~48.01%(均值为30.09%)和12.86%~35.33%(均值为19.06%); 而以乙酸钠和葡萄糖混合液作为外源碳时, 得到的夏季和冬季生物贡献率分别仅为11.12%~46.48%(均值为26.83%)和8.30%~13.77%(均值为11.84%).事实上, Khoshmanesh等[24]在对湿地沉积物的研究中也发现, 当以葡萄糖为碳源, 生物吸收贡献率可以由最初的9%快速上升到40%;而当以乙酸酯为碳源, 生物吸收贡献率则可以进一步达到45%, 即也表现出乙酸酯的作用效果高于葡萄糖. Khoshmanesh等[24]认为, 这一现象的产生可能是因为葡萄糖被大部分用于细胞生长, 产生大量的CO2, 使水体pH值降低, 导致沉积物颗粒对磷吸附的点位减少, 致使磷的生物吸收能力减弱; 而乙酸钠与葡萄糖混合溶液并没有为聚磷菌(PAO)的生长提供有利条件, 此时可能是其它细菌主导了沉积物磷的释放.
此外, 生物对磷的吸收与磷赋存形态有很大的关系.一般地, Ex-P、Fe/Al-P等易于被微生物吸收利用[25, 26], 因此沉积物中生物有效性磷含量及其在总磷中的占比情况, 可能影响磷的生物/非生物吸收贡献率.总的来说, 在十五里河城乡梯度方向, 5个采样点位都表现为添加乙酸钠的沉积物磷的生物吸收贡献率最高, 添加葡萄糖次之, 而添加乙酸钠与葡萄糖混合溶液带来的生物吸收贡献率最低.
4 结论(1) 十五里河沉积物氮磷污染严重, 但有机质含量相对并不突出, 且沉积物氮、磷和有机质含量并未表现出显著的城乡梯度特征.
(2) 十五里河表层沉积物磷的生物吸收潜力显著低于非生物吸收, 其中夏季生物和非生物吸收潜力均值分别为0.784 μg·(g·h)-1和3.016 μg·(g·h)-1, 冬季生物和非生物吸收潜力均值分别为0.323 μg·(g·h)-1和3.368 μg·(g·h)-1.
(3) 外源碳添加不仅可以提升十五里河沉积物磷的生物吸收潜力, 也可以提高生物因素在沉积物磷吸收中的重要性, 并以乙酸钠添加带来的影响效果最显著, 其次是葡萄糖, 乙酸钠与葡萄糖混合溶液的作用效果相对较弱.
| [1] | Maitra N, Manna S K, Samanta S, et al. Ecological significance and phosphorus release potential of phosphate solubilizing bacteria in freshwater ecosystems[J]. Hydrobiologia, 2015, 745(1): 69-83. DOI:10.1007/s10750-014-2094-z |
| [2] |
刘佳, 雷丹, 李琼, 等. 黄柏河流域梯级水库沉积物磷形态特征及磷释放通量分析[J]. 环境科学, 2018, 39(4): 1608-1615. Liu J, Lei D, Li Q, et al. Characteristics of phosphorus fractions and phosphate diffusion fluxes of sediments in cascade reservoirs of the Huangbai River[J]. Environmental Science, 2018, 39(4): 1608-1615. |
| [3] | Khoshmanesh A, Hart B T, Duncan A, et al. Luxury uptake of phosphorus by sediment bacteria[J]. Water Research, 2002, 36(3): 774-778. |
| [4] | Jaisi D P, How Y, Stout L M, et al. Modeling of biotic and abiotic processes affecting phosphate oxygen isotope ratios in a mineral-water-biota system[J]. Water Research, 2017, 126: 262-273. DOI:10.1016/j.watres.2017.09.025 |
| [5] | Jaisi D P, Kukkadapu R K, Stout L M, et al. Biotic and abiotic pathways of phosphorus cycling in minerals and sediments:Insights from oxygen isotope ratios in phosphate[J]. Environmental Science & Technology, 2011, 45(15): 6254-6261. |
| [6] | Sutter M I, Demars B O L, Langan S J. River phosphorus cycling:Separating biotic and abiotic uptake during short-term changes in sewage effluent loading[J]. Water Research, 2010, 44(15): 4425-4436. DOI:10.1016/j.watres.2010.06.014 |
| [7] | Meng J, Yao Q Z, Yu Z G. Particulate phosphorus speciation and phosphate adsorption characteristics associated with sediment grain size[J]. Ecological Engineering, 2014, 70: 140-145. DOI:10.1016/j.ecoleng.2014.05.007 |
| [8] | Mangadze T, Bere T, Mwedzi T. Epilithic diatom flora in contrasting land-use settings in tropical streams, Manyame Catchment, Zimbabwe[J]. Hydrobiologia, 2015, 753(1): 163-173. DOI:10.1007/s10750-015-2203-7 |
| [9] | Wan J J, Liu X M, Wu C X, et al. Nutrient capture and recycling by periphyton attached to modified agrowaste carriers[J]. Environmental Science and Pollution Research, 2016, 23(8): 8035-8043. DOI:10.1007/s11356-015-5988-z |
| [10] | Scinto L J, Reddy K R. Biotic and abiotic uptake of phosphorus by periphyton in a subtropical freshwater wetland[J]. Aquatic Botany, 2003, 77(3): 203-222. DOI:10.1016/S0304-3770(03)00106-2 |
| [11] | McCormick P V, Shuford Ⅲ R B E, Chimney M J. Periphyton as a potential phosphorus sink in the everglades nutrient removal project[J]. Ecological Engineering, 2006, 27(4): 279-289. DOI:10.1016/j.ecoleng.2006.05.018 |
| [12] | Wolfe Ⅲ J E, Lind O T. Phosphorus uptake and turnover by periphyton in the presence of suspended clays[J]. Limnology, 2010, 11(1): 31-37. DOI:10.1007/s10201-009-0287-3 |
| [13] | Lu H Y, Wan J J, Li J Y, et al. Periphytic biofilm:A buffer for phosphorus precipitation and release between sediments and water[J]. Chemosphere, 2016, 144: 2058-2064. DOI:10.1016/j.chemosphere.2015.10.129 |
| [14] | McDowell R W, Elkin K R, Kleinman P J A. Temperature and nitrogen effects on phosphorus uptake by agricultural stream-bed sediments[J]. Journal of Environmental Quality, 2017, 46(2): 295-301. |
| [15] | Li S S, Yuan Z W, Bi J, et al. Anthropogenic phosphorus flow analysis of Hefei City, China[J]. Science of the Total Environment, 2010, 408(23): 5715-5722. DOI:10.1016/j.scitotenv.2010.08.052 |
| [16] |
李如忠, 秦如彬, 高苏蒂, 等. 城乡交错带典型溪流沟渠氮素污染特征及生态风险分析[J]. 华北水利水电大学学报(自然科学版), 2017, 38(3): 27-34. Li R Z, Qin R B, Gao S D, et al. Investigation on the characteristics and ecological risk of nitrogen pollution in the streams across the ditches in a urban-rural fringe[J]. Journal of North China University of Water Resources and Electric Power (Natural Science Edition), 2017, 38(3): 27-34. DOI:10.3969/j.issn.1002-5634.2017.03.004 |
| [17] |
李如忠, 阙凤翔, 熊鸿斌, 等. 巢湖十五里河河床地貌单元沉积物硝化速率及污染特征[J]. 环境科学, 2019, 40(1): 211-218. Li R Z, Que F X, Xiong H B, et al. Nitrification rates and pollution characteristics of sediments with different geomorphic features in the Shiwuli Stream, Chaohu Lake basin[J]. Environmental Science, 2019, 40(1): 211-218. |
| [18] |
李如忠, 李峰, 周爱佳, 等. 巢湖十五里河沉积物氮磷形态分布及生物有效性[J]. 环境科学, 2012, 33(5): 1503-1510. Li R Z, Li F, Zhou A J, et al. Distribution and bioavailability of nitrogen and phosphorus species in the sediments from Shiwuli Stream in Lake Chaohu[J]. Environmental Science, 2012, 33(5): 1503-1510. |
| [19] | Lottig N R, Stanley E H. Benthic sediment influence on dissolved phosphorus concentrations in a headwater stream[J]. Biogeochemistry, 2007, 84(3): 297-309. DOI:10.1007/s10533-007-9116-0 |
| [20] |
李如忠, 叶舟, 高苏蒂, 等. 人为扰动背景下城郊溪流底质磷的生物-非生物吸收潜力分析[J]. 环境科学, 2017, 38(8): 3235-3242. Li R Z, Ye Z, Gao S D, et al. Biotic and abiotic uptake of phosphorus in benthic sediments of suburban streams under intense human disturbance scenario[J]. Environmental Science, 2017, 38(8): 3235-3242. |
| [21] | Gibson C A, O'reilly C M. Organic matter stoichiometry influences nitrogen and phosphorus uptake in a headwater stream[J]. Freshwater Science, 2012, 31(2): 395-407. DOI:10.1899/11-033.1 |
| [22] | Schade J D, Macneill K, Thomas S A, et al. The stoichiometry of nitrogen and phosphorus spiralling in heterotrophic and autotrophic streams[J]. Freshwater Biology, 2011, 56(3): 424-436. DOI:10.1111/fwb.2011.56.issue-3 |
| [23] | Aldridge K T, Brookes J D, Ganf G G. Changes in abiotic and biotic phosphorus uptake across a gradient of stream condition[J]. River Research and Applications, 2010, 26(5): 636-649. |
| [24] | Khoshmanesh A, Hart B T, Duncan A, et al. Biotic uptake and release of phosphorus by a wetland sediment[J]. Environmental Technology, 1999, 20(1): 85-91. DOI:10.1080/09593332008616796 |
| [25] |
裴婷婷, 李如忠, 高苏蒂, 等. 合肥城郊典型农田溪流水系统沉积物磷形态及释放风险分析[J]. 环境科学, 2016, 37(2): 548-557. Pei T T, Li R Z, Gao S D, et al. Phosphorus fractions and release risk in surface sediments of an agricultural headwater stream system in Hefei Suburban, China[J]. Environmental Science, 2016, 37(2): 548-557. |
| [26] |
丁瑶, 欧阳莉莉, 石清, 等. 特大城市河流表层沉积物磷形态分布及有效性:以成都市为例[J]. 环境科学, 2019, 40(1): 219-227. Ding Y, Ouyang L L, Shi Q, et al. Distribution and bioavability of phosphorus in surface sediments in megalopolis:A case study of Chengdu[J]. Environmental Science, 2019, 40(1): 219-227. |