 2019, Vol. 40
2019, Vol. 40



2. 中国联合工程有限公司, 杭州 310052
2. China United Engineering Corporation Limited, Hangzhou 310052, China
目前生物脱氮工艺主要依赖自养硝化和厌氧反硝化过程, 然而硝化和反硝化过程难以同步完成, 使得污水处理厂的工艺流程复杂、占地面积大、基建费用高.此外, 随着含氮化合物排放增加, 传统脱氮工艺逐步暴露出水力停留时间长、能耗高、抗冲击能力弱等不足.
近年来, 越来越多的异养硝化细菌被发现, 主要包括产碱菌属(Alcaligenes)[1]、假单胞菌属(Pseudomonas)[2]、芽孢杆菌(Bacillus)[3]和不动杆菌(Acinetobacter)[4]等.相比于自养硝化细菌, 异养硝化细菌具有生长速率快、活性高、增殖底物广泛以及环境适应性强等优点.同时, 有研究发现大部分异养硝化菌能够同步进行有机物降解、硝化和反硝化作用[3, 4], 使得脱氮反应在同一反应器中进行成为可能, 大大减少了占地面积与运行费用.基于异养硝化细菌独特的生物学特性, 有研究人员发现一些异养硝化细菌在生活污水、高氨氮养猪废水、含重金属工业废水处理等方面均具有良好的表现[1, 5, 6], 具有显著的工程应用价值.然而, 目前对于异养硝化技术的研究还处于起步阶段, 在生物学、动力学、脱氮机制等方面的认识仍不明确, 极大地限制了该技术在实际生产中的应用.因此, 加强高效异养硝化细菌脱氮特性及代谢机制方面的研究, 对丰富生物脱氮理论以及指导工程实践具有重要意义.
本文以异养硝化细菌Pseudomonas putida YH为研究对象, 开展其生理生化特性、脱氮性能和影响因子分析, 并通过构建Logistic和Compertz模型分别对YH的脱氮特性与菌体生长特性进行模拟, 以探究其氮素降解动力学和菌体生长动力学特性.
1 材料与方法 1.1 菌株来源异养硝化菌株Pseudomonas putida YH (GenBank ID: # KJ765710)从实验室SBR活性污泥反应器中分离纯化得到, 用甘油冷冻法保存在实验室, 使用前富集培养[7].
1.2 培养基异养硝化培养基(BM, g·L-1):(NH4)2SO4 0.47, 琥珀酸钠5.62, 维氏盐溶液50 mL, pH=7.0.
亚硝酸盐培养基(DM1, g·L-1):NaNO20.49, 琥珀酸钠5.62, 微量元素溶液2 mL, pH=7.0.
硝酸盐培养基(DM2, g·L-1):KNO3 0.72, 琥珀酸钠5.62, 微量元素溶液2 mL, pH=7.0.
维氏盐溶液(g·L-1):K2HPO4·3H2O 5.0, MgSO4·7H2O 2.5, NaCl 2.5, MnSO4·4H2O 0.05, FeSO4·7H2O 0.05.
微量元素溶液(g·L-1):Na2EDTA 50.0, ZnSO4·7H2O 2.2, CaCl2 5.5, MnCl2·4H2O 5.06, FeSO4 5.0, CuSO4·5H2O 1.57, CoCl2·6H2O 1.60.
1.3 菌株形态及生理生化分析将菌株YH在异养硝化培养基中活化后, 涂布于异养硝化固体培养基上, 30℃培养48 h后观察菌落特征; 将菌株YH进行革兰氏染色, 在光学显微镜下观察细胞形态; 分离菌株的个体形态通过扫描电镜观察; 菌株生理生化特性依据文献[8]进行分析.
1.4 亚硝酸盐还原酶(napA)和硝酸盐还原酶(nirK、nirS)的扩增菌株YH基因组DNA通过细菌DNA提取试剂盒进行提取(沃特森, 上海). PCR反应中所需的生物试剂由Takara生物技术有限公司提供, PCR引物由上海生工技术有限公司合成. PCR引物及其序列见表 1, 反应条件见表 2.反应采用25 μL反应体系:10×Buffer 2.5 μL, dNTP 2 μL, 正反引物各1 μL, Ex Taq酶0.2 μL, DNA模板2 μL, ddH2O 16.3 μL. PCR扩增产物通过1%琼脂糖凝胶电泳进行检测, 经核酸染料染色后, 通过凝胶成像系统成像拍照(G:Box EF, Syngene, USA).
|
|
表 1 PCR引物 Table 1 PCR primers |
|
|
表 2 PCR反应条件 Table 2 PCR reaction conditions |
1.5 菌株YH的异养硝化和好氧反硝化特性分析
以琥珀酸钠为碳源, 分别采用氨氮、亚硝酸盐和硝酸盐作为单一氮源, 氨氮、亚硝氮和硝氮初始浓度均为100 mg·L-1, C/N=10.在接种量为1%、30℃、160 mg·L-1条件下培养48 h, 定时测定其D600和氮浓度.
1.6 不同反应条件对菌株YH异养硝化性能的影响分别研究碳源、C/N、pH、温度、转速和氨氮初始浓度对氨氮去除效果的影响.碳源包括葡萄糖、乙酸钠、蔗糖、柠檬酸钠和琥珀酸钠; C/N分别为2、5、10和15; pH分别为5、6、7、8、9、10和11;温度分别为10、20、30和37℃; 摇床转速分别为80、120、160和200 r·min-1; 不同初始氨氮浓度分别取50、200、500和1 000 mg·L-1.反应过程中除改变单一影响因子, 其余反应条件固定不变, 分别为氨氮浓度100 mg·L-1、温度30℃、pH=7.0、C/N=10以及转速160 r·min-1.将处于对数生长期的菌液以1%接种量接种至经高压灭菌装有100 mL不同培养基的250 mL锥形瓶中, 摇床培养24 h, 定时取样测定其D600和氨氮浓度.
1.7 生长和氨氮降解动力学分析采用Logistic模型[9]拟合菌株YH异养硝化-好氧反硝化过程菌株生长特性.
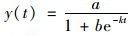
|
(1) |
式中, a、b、k均为Logistic生长方程常数; e为数学常数.
菌株YH的异养硝化-好氧反硝化氮素降解过程采用修饰过的Compertz模型[4]进行拟合.
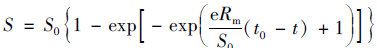
|
(2) |
式中, S为某一时刻氮素浓度, mg·L-1; S0为基质初始浓度, mg·L-1; Rm为最大转化速率, mg·(L·h)-1; t0为迟滞时间, h; t为反应时间, h; e为数学常数.
1.8 分析方法NH4+-N、NO2--N、NO3--N和TN浓度均采用标准方法测定[10]; 菌体生长吸光度(D600)采用吸光度法测定; 动力学拟合采用Oringin 9.1.
2 结果与讨论 2.1 菌株形态特征和生理生化特性菌株YH菌落呈淡黄色、直径约1 mm、不透明、圆形、凸起、表面光滑.革兰氏染色为阴性.通过电镜照片(图 1)可知, 菌株YH呈杆状, 无鞭毛, 大小为0.5 μm×(0.9~1.0) μm.生理生化检测结果如表 3所示.

|
图 1 菌株YH电镜扫描照片 Fig. 1 Scanning electron micrograph of strain YH |
|
|
表 3 菌株YH的生理生化特征1) Table 3 Chemical and physical characteristics of strain YH |
2.2 菌株YH异养硝化-好氧反硝化脱氮特性分析 2.2.1 异养硝化特性
菌株YH的氨氮降解过程和中间产物变化情况如图 2所示.经过约3 h的适应期后, 菌株开始快速生长, 在18 h达到稳定期, D600最高达到1.359, 生长过程中最大比生长速率为0.49 h-1.与Thiosphaera pantotropha[11] 0.28~0.45 h-1, Alcligenes faecalis No.4[12] 0.2 h-1等菌相当.采用Logistic模型对菌株YH异养硝化过程生长特性进行模拟, 如图 2(b)所示, 拟合所得R2大于0.99, 表明该模型能够充分表征菌株YH的生长特性.

|
图 2 以氨氮为唯一氮源时的氮素和D600变化情况 Fig. 2 Changes in nitrogen compounds and D600 when ammonium was used as the sole nitrogen source |
同时, 反应过程氨氮最大去除率为99.1%, 平均氨氧化速率为8.35 mg·(L·h)-1, 与自养硝化细菌[7.0 mg·(L·h)-1]在同一数量级[13], 但高于Klebsiella pneumoniae CF-S9[14][4.3 mg·(L·h)-1]、Rhodococcus sp. CPZ24[15][3.1 mg·(L·h)-1]等异养硝化细菌.降解过程中仅有少量的硝化中间产物(亚硝氮和硝氮)生成.反应24 h后氨氮浓度逐渐增加(氨氮最终去除率为98.9%), 说明此时菌群进入内源呼吸期, 之前用于细胞合成的氮源又再次分解为氨氮回到水中.由于硝化产物含量较低且快速降解, TN去除趋势几乎与氨氮一致, 反应48 h菌株YH的TN去除率为98.5%.通过氮物质平衡分析, 菌株YH菌体生长合成的内源氮约占去除TN的53%, 其余部分通过好氧反硝化去除.
2.2.2 好氧反硝化特性由图 3可见, 当亚硝酸盐和硝酸盐作为单一氮源时, 菌株YH生长良好, D600均在18~24 h取得最大值1.386和1.276, 表明菌株YH能够利用亚硝氮和硝氮生长代谢, 这与菌株Acinetobacter sp. YN3[16]、Enterobacter cloacae HW-15[17]等一致.以亚硝酸盐为唯一氮源时, 菌株YH最大比生长速率和亚硝氮最大降解速率出现在6~9 h, 分别为0.27 h-1和8.78 mg·(L·h)-1.反应中有微量的氨氮和硝氮生成, 亚硝氮和总氮最终去除率分别为99.5%和98%.相比于菌株Bacillus sp. YX-6[18]等菌在亚硝氮浓度为20~40 mg·L-1时活性便受到抑制, 菌株YH能利用更高浓度的亚硝氮.以硝酸盐为唯一氮源时, 菌株YH最大比生长速率和最大硝氮降解速率出现在6~9 h, 分别为0.31 h-1和8.86 mg·(L·h)-1.硝氮和总氮最终去除率分别为99.8%和98%.硝氮降解初期存在亚硝氮累积现象, 最大累积量为9.33 mg·L-1, 表明菌株对数生长期, 高浓度硝氮较多被转化为亚硝氮, 而亚硝氮反硝化不及时致使亚硝氮累积.但不同于菌株YL[19]只是快速地将硝氮转化为亚硝氮而不完全去除, 菌株YH好氧反硝化所累积的亚硝氮最终会完全降解.氮平衡分析表明分离菌株菌体生长合成的内源氮占去除TN的51%~55%, 剩余损失TN则通过好氧反硝化作用去除.

|
图 3 以亚硝酸盐和硝酸盐为唯一氮源时菌株YH的好氧反硝化性能 Fig. 3 Aerobic denitrification capabilities of strain YH when nitrite and nitrate were used as the sole N-source, respectively |
如表 4所示, Compertz模型可以较好地描述菌株YH异养硝化-好氧反硝化过程中氮素的降解情况, 拟合所得最大降解速率Rm为氨氮>硝氮>亚硝氮, 迟滞时间t0为硝氮>亚硝氮>氨氮, 显著优于菌株Acinetobacter sp. JQ1004[氨氮:Rm=7.9 mg·(L·h)-1, t0=5.9 h; 硝氮Rm=4.1 mg·(L·h)-1, t0=12.7 h][4].以氨氮为唯一氮源时, 氮素最大降解速率最高, 迟滞期最短, 其原因可能是异养硝化-好氧反硝化过程中氨氧化酶活性高于好氧反硝化酶活性, 以及高浓度硝酸盐和亚硝酸盐对微生物活性有一定的抑制作用[20].
|
|
表 4 菌株YH降解不同氮素的动力学参数与最终去除率 Table 4 Kinetic parameters and final removal efficiencies of strain YH in the biodegradation of different nitrogen sources |
2.2.3 好氧反硝化功能基因(napA、nirK)扩增
采用引物NAP1/NAP2对菌株YH的NAR进行基因扩增, 如图 4所示, 菌株YH在870 bp左右处得到清晰的目标条带, 与周质NAR的大亚基napA基因相对应, 因此可初步判断分离菌株含napA基因.利用引物nirK1F/nirK5R和nirS1F/nirS6R对菌株YH的DNA样品进行扩增, 菌株YH仅在514bp左右得到清晰的特异性条带, 表明基因nirK在菌株YH细胞中成功表达, 而nirS未能表达, 说明菌株YH含有铜离子的NIR, 但不存在细胞色素cd1的NIR, 此结果与一种细菌中只有一类NIR存在的结论相一致[21].本研究napA和nirK基因的成功表达, 进一步表明了NIR和NAR的存在, 在分子水平证明了菌株YH具有好氧反硝化的能力.基于菌株YH对不同氮源的利用转化、中间产物的生成规律以及好氧反硝化功能基因的检测分析, 推测其脱氮途径为:
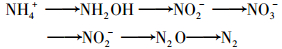
|

|
图 4 菌株YH的napA和nirK基因PCR扩增 Fig. 4 Amplification of napA and nirK in strain YH by PCR |
当采用葡萄糖和蔗糖作为碳源时, 菌株YH生长缓慢, D600最高值仅0.218和0.082;氨氮降解效果较差, 最终去除率分别为49.5%和25.5%(表 5).拟合所得Rm分别为1.16 mg·(L·h)-1和0.56 mg·(L·h)-1, 说明菌株YH难以利用葡萄糖、蔗糖等大分子糖类为碳源降解氨氮.在采用柠檬酸钠、乙酸钠和琥珀酸钠为碳源时, 菌体生长好且氨氮基本完全去除.与柠檬酸钠和乙酸钠相比, 琥珀酸钠的Rm提高至16.71mg·(L·h)-1, t0缩短为2.28 h, 是一种最理想的碳源.主要因为它是微生物将复杂有机物(如糖类、蛋白质和脂类)转化为简单小分子有机物时三羧酸循环过程中的中间产物, 能够直接用于生物合成.这与菌株Pseudomonas sp. N6[22]、Acinetobacter junii YB[23]对有机酸的利用率高于糖类的结论一致.
|
|
表 5 菌株YH在不同环境条件下的氨氮降解动力学参数与最终去除率 Table 5 Kinetic parameters and final removal efficiencies for the degradation of ammonium by strain YH under different environmental conditions |
2.3.2 C/N
低C/N条件下(C/N为2、5), 氨氮去除率低, 菌株生长情况差, 主要是由于碳源不足导致异养硝化效果不完全.在C/N=10时, 菌株YH生长良好, 菌体生长量最大为1.359, 氨氮最终去除率为98.9%.当C/N提高到15时, 菌体生长(D600最大为1.781)和氨氮去除率(99.9%)均有所提高.如表 5所示, C/N为10和15时, 迟滞时间分别为1.08 h和0.86 h, 氨氮最大转化速率分别为12.99 mg·(L·h)-1和15.09 mg·(L·h)-1.与Alcaligenes faecalis OKK17[24]等菌相比, 菌株YH能够适应更高的C/N环境, 而且C/N的越高, 去除速率越快.但C/N提高到15时, 氨氮降解速率只有略微增加, 而氨氮去除率并没有明显变化, C/N过高会导致资源的浪费, 综合考虑运行效果和经济效益, 确定菌株YH脱氮的最适宜C/N为10.
2.3.3 温度根据表 5的拟合结果, t0随温度的升高明显降低, 说明温度越高越有利于接种初期细菌适应环境.温度为30℃时, Rm取得最大值16.71 mg·(L·h)-1, 表明氨氮降解速率在30℃达到最佳.在30℃和37℃条件下, 氨氮去除率均在12 h达到最高, 分别为99.29%和99.85%.但在37℃时, D600最大为1.347低于30℃时的1.359, 且反应15 h后出现氨氮不断升高的现象(氨氮最终去除率为83.3%), 这可能因为温度升高, 生物活性物质变性, 细胞功能下降甚至死亡, 最终影响反应效果[25].综合比较而言, 菌株YH的最佳反应温度为30℃, 此结果与所报道的大多数异养硝化菌的最佳温度(30~37℃)一致[2, 6, 7].近些年研究发现一些嗜冷型异养硝化细菌, 这类细菌在低温条件下仍然具有较高的硝化活性.值得注意的是, 菌株YH在低温条件下具有良好的脱氮能力.根据拟合结果, 当培养温度为10℃时, t0=8.43 h, Rm=3.29 mg·(L·h)-1, YH需要更长的时间以适应低温环境, 但经过48h反应, 氨氮最终去除率仍高达99.2%.菌株YH低温环境下较强的适应能力将为寒冷地区污水生物脱氮提供高效菌源[26, 27].
2.3.4 转速转速对菌株YH异养硝化的影响结果如表 5所示, t0随着转速的提高而缓慢增加, 表明菌株YH需要更长的时间适应高DO环境.就氨氮处理效果而言, DO对于菌株YH的异养硝化效果影响较小, 在4种转速条件下, 氨氮的最终去除率均在99%左右.但是, 菌株YH的氨氮降解速率和菌体生长情况随转速的提高而明显改善, 氨氮最大转化速率由4.64 mg·(L·h)-1提高到25.84 mg·(L·h)-1, 160 r·min-1和200 r·min-1条件下D600最高分别为1.359和1.455.故转速在160~200 r·min-1较为适宜, 这与肖继波等[28]研究异养硝化菌Colloides sp. JZ1-1时所得结论一致.
|
|
表 6 菌株YH在不同初始氨氮浓度下的氨氮降解动力学参数与最终去除率 Table 6 Kinetic parameters and final removal efficiencies for the degradation of ammonium by strain YH under different initial ammonium concentrations |
2.3.5 pH
pH对于菌株YH氨氮去除效果影响较小, 但对其氨氮降解速率存在明显影响. pH在5~10范围内氨氮最终去除率均高于90%, 当pH提高到11时, 氨氮最终去除率略有下降, 相较于Pseudomonas sp. GL19[27]等异养硝化菌, 菌株YH的pH耐受范围更广, 去除率更高.从表 5可以看出, 弱碱性条件(pH为7~9)下Rm最高, 菌体生长也呈现出相同的趋势.已报道的诸多异养硝化菌均适宜在弱碱性条件下生长, 原因可能是弱碱性环境中较多的游离氨促进了氨氮加氧酶的氧化[29, 30]. Rm在pH=7时最大, 表明pH=7为菌株YH异养硝化的最佳pH.
2.3.6 氨氮浓度起始浓度为50 mg·L-1时, 氨氮能够在9 h内基本完全降解, 此时由于氮源不足, 菌体生长受到一定限制, 最高D600仅为0.963.起始浓度提高到100 mg·L-1和200 mg·L-1, 氨氮在30 h内分别降解至0.91 mg·L-1和0.61 mg·L-1, 菌体生长有所改善(D600最大分别为1.359和1.849).当起始浓度为500 mg·L-1和1 000 mg·L-1时, 菌体生长并未受限, 最高D600分别为1.917和1.853, 48 h内氨氮分别降至165.23 mg·L-1和406.70 mg·L-1, 氨氮最终去除率分别为66.7%和58.9%.起始浓度较低时, 氨氮去除较完全, 随着起始浓度的提高, 菌株需要的反应时间延长.拟合结果显示(表 6), 随着起始浓度由50 mg·L-1提高到200 mg·L-1, t0从1.07 h上升到4.03 h, 表明菌株适应期变长; Rm由13.97 mg·(L·h)-1增加至28.70 mg·(L·h)-1, 在上述起始浓度范围内, 菌株YH异养硝化速率同起始浓度成正相关关系.与Bacillus sp. LY[31]等菌株随起始浓度升高, 氨氮最大转化速率逐渐降低不同, 菌株YH随着起始浓度提高至1 000 mg·L-1, Rm明显提高, t0则相对缩短.菌株YH能够适应较宽范围的氨氮负荷, 在高氨氮负荷下仍具有较高的异养硝化能力, 表明菌株YH具有处理高浓度氨氮废水的潜能.
3 结论(1) 菌株YH可以利用氨氮、亚硝酸盐和硝酸盐作为唯一氮源生长代谢, 最大去除率分别为99.1%、99.5%、99.8%, 结合反硝化功能基因napA和nirK的PCR成功扩增, 进一步证明菌株YH具有好氧反硝化特性.
(2) YH菌体生长特性与Logistic模型相匹配(R2>0.99), 菌株YH对于不同氮素的降解特性则符合修饰过的Compertz模型(R2>0.99), 拟合所得氮素最大转化速率Rm为氨氮>硝氮>亚硝氮, 迟滞时间t0分别为硝氮>亚硝氮>氨氮.拟合参数的确定有助于进一步明确菌株YH的代谢特征.
(3) 菌株YH异养硝化最适宜的条件是碳源为琥珀酸钠、C/N=10、T=30℃、r为160~200 r·min-1以及pH=7, 最优条件下氨氧化平均速率和最大转化速率分别为8.35 mg·(L·h)-1和16.71 mg·(L·h)-1.
(4) 菌株YH能够适应较宽范围的氨氮负荷, 氨氮起始浓度提高至500 mg·L-1和1 000 mg·L-1, 氨氮最终去除率分别为66.7%和58.9%, Rm进一步提高且菌体生长未受明显抑制, 表明菌株YH具有处理高浓度氨氮废水的潜能.
| [1] | Joo H S, Hirai M, Shoda M. Piggery wastewater treatment using Alcaligenes faecalis strain No. 4 with heterotrophic nitrification and aerobic denitrification[J]. Water Research, 2006, 40(16): 3029-3036. DOI:10.1016/j.watres.2006.06.021 |
| [2] | Zhang J B, Wu P X, Hao B, et al. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001[J]. Bioresource Technology, 2011, 102(21): 9866-9869. DOI:10.1016/j.biortech.2011.07.118 |
| [3] | Rout P R, Bhunia P, Dash R R. Simultaneous removal of nitrogen and phosphorous from domestic wastewater using Bacillus cereus GS-5 strain exhibiting heterotrophic nitrification, aerobic denitrification and denitrifying phosphorous removal[J]. Bioresource Technology, 2017, 244: 484-495. DOI:10.1016/j.biortech.2017.07.186 |
| [4] |
王秀杰, 王维奇, 李军, 等. 异养硝化菌Acinetobacter sp.的分离鉴定及其脱氮特性[J]. 中国环境科学, 2017, 37(11): 4241-4250. Wang X J, Wang W Q, Li J, et al. Isolation and identification of a heterotrophic nitrifier, Acinetobacter sp., and its characteristics of nitrogen removal[J]. China Environmental Science, 2017, 37(11): 4241-4250. DOI:10.3969/j.issn.1000-6923.2017.11.029 |
| [5] | Sun Z Y, Lv Y T, Liu Y X, et al. Removal of nitrogen by heterotrophic nitrification-aerobic denitrification of a novel metal resistant bacterium Cupriavidus sp. S1[J]. Bioresource Technology, 2016, 220: 142-150. DOI:10.1016/j.biortech.2016.07.110 |
| [6] | Su J F, Shi J X, Huang T L, et al. Kinetic analysis of simultaneous denitrification and biomineralization of novel Acinetobacter sp. CN86[J]. Marine Pollution Bulletin, 2016, 109(1): 87-94. DOI:10.1016/j.marpolbul.2016.06.017 |
| [7] | Yang L, Ren Y X, Zhao S Q, et al. Isolation and characterization of three heterotrophic nitrifying-aerobic denitrifying bacteria from a sequencing batch reactor[J]. Annals of Microbiology, 2016, 66(2): 737-747. DOI:10.1007/s13213-015-1161-7 |
| [8] | 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001. |
| [9] | Chen J, Zhao B, An Q, et al. Kinetic characteristics and modelling of growth and substrate removal by Alcaligenes faecalis strain NR[J]. Bioprocess and Biosystems Engineering, 2016, 39(4): 593-601. DOI:10.1007/s00449-016-1541-9 |
| [10] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002: 254-279. |
| [11] | Robertson L A, Kuenen J G. Heterotrophic nitrification in Thiosphaera pantotropha:oxygen uptake and enzyme studies[J]. Journal of General Microbiology, 1988, 134(4): 857-863. |
| [12] | Joo H S, Hirai M, Shoda M. Characteristics of ammonium removal by heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis No. 4[J]. Journal of Bioscience and Bioengineering, 2005, 100(2): 184-191. DOI:10.1263/jbb.100.184 |
| [13] | 张自杰. 排水工程[M]. (第四版). 北京: 中国建筑工业出版社, 1999: 311-312. |
| [14] | Padhi S K, Tripathy S, Sen R, et al. Characterisation of heterotrophic nitrifying and aerobic denitrifying Klebsiella pneumoniae CF-S9 strain for bioremediation of wastewater[J]. International Biodeterioration & Biodegradation, 2013, 78: 67-73. |
| [15] | Chen P Z, Li J, Li Q X, et al. Simultaneous heterotrophic nitrification and aerobic denitrification by bacterium Rhodococcus sp. CPZ24[J]. Bioresource Technology, 2012, 116: 266-270. DOI:10.1016/j.biortech.2012.02.050 |
| [16] |
颜薇芝, 张汉强, 余从田, 等. 1株异养硝化好氧反硝化不动杆菌的分离及脱氮性能[J]. 环境工程学报, 2017, 11(7): 4419-4428. Yan W Z, Zhang H Q, Yu C T, et al. Isolation of Acinetobacter sp. YN3 and its heterotrophic nitrification-aerobic denitrification characters[J]. Chinese Journal of Environmental Engineering, 2017, 11(7): 4419-4428. |
| [17] | Wan W J, He D L, Xue Z J. Removal of nitrogen and phosphorus by heterotrophic nitrification-aerobic denitrification of a denitrifying phosphorus-accumulating bacterium Enterobacter cloacae HW-15[J]. Ecological Engineering, 2017, 99: 199-208. DOI:10.1016/j.ecoleng.2016.11.030 |
| [18] | Song Z F, An J, Fu G H, et al. Isolation and characterization of an aerobic denitrifying Bacillus sp. YX-6 from shrimp culture ponds[J]. Aquaculture, 2011, 319(1-2): 188-193. DOI:10.1016/j.aquaculture.2011.06.018 |
| [19] |
梁贤, 任勇翔, 杨垒, 等. 异养硝化-好氧反硝化菌YL的脱氮特性[J]. 环境科学, 2015, 36(5): 1749-1755. Liang X, Ren Y X, Yang L, et al. Characteristics of nitrogen removal by a heterotrophic nitrification-aerobic denitrification bacterium YL[J]. Environmental Science, 2015, 36(5): 1749-1755. |
| [20] | Yang L, Ren Y X, Liang X, et al. Nitrogen removal characteristics of a heterotrophic nitrifier Acinetobacter junii YB and its potential application for the treatment of high-strength nitrogenous wastewater[J]. Bioresource Technology, 2015, 193: 227-233. DOI:10.1016/j.biortech.2015.05.075 |
| [21] | Braker G, Zhou J Z, Wu L Y, et al. Nitrite reductase genes (nirK and nirS) as functional markers to investigate diversity of denitrifying bacteria in pacific northwest marine sediment communities[J]. Applied and Environmental Microbiology, 2000, 66(5): 2096-2104. DOI:10.1128/AEM.66.5.2096-2104.2000 |
| [22] |
姜磊, 徐成斌, 马溪平, 等. 1株好氧反硝化菌的分离鉴定和反硝化特性研究[J]. 环境科学与技术, 2013, 36(3): 12-15. Jiang L, Xu C B, Ma X P, et al. Identification and denitrification characteristics of an aerobic denitirfier[J]. Environmental Science & Technology, 2013, 36(3): 12-15. |
| [23] | Ren Y X, Yang L, Liang X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB[J]. Bioresource Technology, 2014, 171: 1-9. DOI:10.1016/j.biortech.2014.08.058 |
| [24] | Nishio T, Yoshikura T, Mishima H, et al. Conditions for nitrification and denitrification by an immobilized heterotrophic nitrifying bacterium Alcaligenes faecalis OKK17[J]. Journal of Fermentation and Bioengineering, 1998, 86(4): 351-356. DOI:10.1016/S0922-338X(99)89003-5 |
| [25] |
杨秀玲, 杨宗政, 庞金钊, 等. 具有硝化作用的异养菌脱除氨氮的性能研究[J]. 天津科技大学学报, 2005, 20(1): 24-26, 48. Yang X L, Yang Z Z, Pang J Z, et al. Study on removal of NH3-N using nitrification and heterotrophic bacteria[J]. Journal of Tianjin University of Science & Technology, 2005, 20(1): 24-26, 48. DOI:10.3969/j.issn.1672-6510.2005.01.007 |
| [26] | Yao S, Ni J R, Ma T, et al. Heterotrophic nitrification and aerobic denitrification at low temperature by a newly isolated bacterium, Acinetobacter sp. HA2[J]. Bioresource Technology, 2013, 139: 80-86. DOI:10.1016/j.biortech.2013.03.189 |
| [27] |
王兆阳, 陈国耀, 姜珂, 等. 1株耐冷兼性嗜碱好氧反硝化菌的分离鉴定及反硝化特性[J]. 环境科学, 2014, 35(6): 2341-2348. Wang Z Y, Chen G Y, Jiang K, et al. Identification and denitrification characteristics of a psychrotolerant facultative basophilic aerobic denitrifier[J]. Environmental Science, 2014, 35(6): 2341-2348. |
| [28] |
肖继波, 江惠霞, 褚淑祎. 新异养硝化菌Colloides sp. JZ1-1的鉴定及其硝化性能[J]. 应用生态学报, 2012, 23: 1635-1640. Xiao J B, Jiang H X, Chu S Y. Identification of a new heterotrophic nitrobacterium strain Colloides sp. JZI-1 and its nitrifying capability[J]. Chinese Journal of Applied Ecology, 2012, 23(6): 1635-1640. |
| [29] | Zhang Q L, Liu Y, Ai G M, et al. The characteristics of a novel heterotrophic nitrification-aerobic denitrification bacterium, Bacillus methylotrophicus strain L7[J]. Bioresource Technology, 2012, 108: 35-44. DOI:10.1016/j.biortech.2011.12.139 |
| [30] | Ye J, Zhao B, An Q, et al. Nitrogen removal by Providencia rettgeri strain YL with heterotrophic nitrification and aerobic denitrification[J]. Environmental Technology, 2016, 37(17): 2206-2213. DOI:10.1080/09593330.2016.1146338 |
| [31] |
何霞, 赵彬, 吕剑, 等. 异养硝化细菌Bacillus sp. LY脱氮性能研究[J]. 环境科学, 2007, 28(6): 1404-1408. He X, Zhao B, Lv J, et al. Nitrogen removal by Bacillus sp. LY with heterotrophic nitrification ability[J]. Environmental Science, 2007, 28(6): 1404-1408. DOI:10.3321/j.issn:0250-3301.2007.06.042 |