 2019, Vol. 40
2019, Vol. 40


2. 农村清洁工程重庆市工程研究中心, 重庆 400716
2. Chongqing Engineering Research Center of Rural Cleaning, Chongqing 400716, China
在处理污水的潜流人工湿地中, 因氧气扩散困难, DO长期处于较低浓度[1].在这种条件下, 植物受到缺氧胁迫[2].在此状况下, 湿地中的植物会根据所处的缺氧环境呈现出生理的适应性及形态结构的改变[3, 4].抵抗湿地缺氧的一个重要结构特征就是通气组织, 通气组织往往形成于植物根部中央[5], 并通过根系孔隙度表现出来.之前有研究证实, 湿地植物根部的通气组织占全部根系体积的60%以上[6~8].湿地植物这种经通气组织释放氧气至根际的行为称为径向氧损失, 又称为根系泌氧(radial oxygen loss, ROL)[9].植物所处环境会对ROL产生影响, 如污染物[10]、重金属[11, 12]、盐度[13], ROL还受到根际环境的氧化还原电位(oxidation-reduction potential, ORP)、pH、DO等[14, 15]的影响.根据Lai等[16]采用微电极对35种湿地植物ROL速率的测定结果发现, 不同植物ROL差异极显著(P=0.000).本文作者前期研究发现, 生物炭添加能够减少菖蒲(AC)体内超氧化物和过氧化物的积累, 显著降低AC体内丙二醛(malonaldehyde, MDA)的含量[17], 但生物炭对缓解缺氧胁迫的具体机制尚不清晰.因此, 本研究拟通过观察植物根系通气组织, 测定根孔隙度和根系ROL, 探究在生物炭缓解湿地植物缺氧(或厌氧)胁迫中, 植物根部组织的响应特征, 以期为生物炭对人工湿地的长期影响研究提供理论参考.
1 材料与方法 1.1 生物炭潜流人工湿地本研究的湿地反应器采用圆筒形聚乙烯容器, 每个容器表面积为0.1 m2, 深为35 cm.按照生物炭40%、30%、20%、10%和0%的体积添加比例分别构建微型系统.生物炭采用芦竹作为原材料于500℃条件自制, 长度为1~2 cm[18], 生物炭的比表面积为345.92 m2 ·g-1, 孔径为1.95 nm, 孔容为0.246 7 cm3 ·g-1, 构建方式按照文献[17]进行, 试验装置如图 1所示.湿地植物取自某生活污水处理厂, 植物经驯化扩培后, 选取长势好且根叶相似的AC, 分别栽入湿地中(对应湿地名称分别命名为AC-40, AC-30, AC-20, AC-10和AC-K), 栽种密度为30株·m-2.湿地微型系统置于温室中运行, 温度(25±2)℃, 光照强度(3 000±300)μmol ·(m2 ·s)-1, 光暗比12 h :12 h.

|
图 1 试验装置示意 Fig. 1 Schematic of the experimental microcosm |
人工湿地进水采用自来水配置, 配好的进水化学需氧量(COD)(414.12±10.67)mg ·L-1, NH4+-N(19.67±1.57)mg ·L-1, 硝态氮(NO3--N)(31.40±1.02)mg ·L-1和总磷(5.22±0.50)mg ·L-1, 进水pH为(7.5±0.3), DO为(2.0±0.5)mg ·L-1, ORP为(150±20)mV.湿地停留时间为2 d, 处理负荷设定为0.05 m3 ·(m2 ·d)-1, 有效进水量为10 L, 进水水位可由顶部水位阀门控制.经过150 d左右稳定运行后, 湿地出水pH为(7.2±0.2)~(7.3±0.1), 出水DO为(0.46±0.05)~(0.48±0.07)mg ·L-1, 出水ORP为(317±5)~(348±5)mV; 湿地主要污染物出水浓度COD为(39.73±8.77)~(50.51±11.77)mg ·L-1, NH4+-N为(8.04±2.54)~(12.71±3.21)mg ·L-1, TN为(9.85±1.61)~(14.53±2.55)mg ·L-1.
1.2 根系通气组织取新生的AC根系, 用超纯水冲洗干净, 然后用吸水纸将其表面水分彻底吸干.将处理干净的根系用封口袋密封, 保存于-20℃条件下备用.用刀片将备用根截断(于根系中间位置), 用环境扫描电镜(FEI, QuantaTM 650 FEG, 美国)对根系截断处进行扫描, 加速电压为10 kV, 放大倍数为400倍, 观察成像.
1.3 根孔隙度参照文献[19]的改进方法, 取新生的AC根系, 用超纯水冲洗干净, 然后用吸水纸将其表面水分彻底吸干.将每条根切成2~2.5 cm小段之后备用.将50 mL比重瓶装满超纯水后称重, 称取0.3 g左右的备用根, 置于装满超纯水的比重瓶中后, 称重.将装有备用根的比重瓶抽真空2 h, 之后取出备用根并置于干燥的研钵中研磨至糊状.将研磨好的根重新置于比重瓶中, 装满超纯水后称重.
根孔隙度计算公式如下:
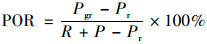
|
式中, POR:根孔隙度, %; Pgr:研磨后的根和装满水的比重瓶的总重量, g; Pr:未经研磨的根和装满水的比重瓶的总重量, g; R:根重, g; P:充满水的比重瓶重量, g.
1.4 ROL测定本试验利用溶解氧微电极(Unisense, MM336155, 丹麦)测定AC的ROL.在烧杯中放入事先灭菌冷却的1.2 L琼胶溶液(1/10改良Hoagland营养液中加入1 g ·L-1琼脂粉, 0.374 g ·L-1 KCl), 曝入高纯N2 15 min, 将刚从湿地拔出的AC洗净, 迅速放入溶液中, 将AC根部全部没入溶液中; 立即在溶液表面铺上一层1.0 cm厚的石蜡, 继续曝高纯N2 15 min.然后立即用溶氧微电极连续1 h跟踪检测溶液中溶解氧的变化, 以空白为对照, 单位时间内琼胶溶液中DO增加量即为ROL.考虑到光照温度对本试验的影响, 光照培养箱中[温度(25±2)℃, 光照强度(0、600、1 500、3 000和3 700)μmol ·(m2 ·s)-1, 植物在测定前预先遮光处理30 min, 处理过程中培养液采用1/10改良Hoagland营养液[20].
1.5 数据分析每组试验设置3个平行, 试验数据通过Origin 8.5整理作图, 并由PASW Statistics 18.0进行数据分析.试验数据表达均采用平均值加或减标准差.对象之间相互关系采用相关性分析, 并经Pearson检验(水平包括显著P<0.05和极显著P<0.01).对象之间的差异性分析采用One-way ANOVA(水平包括显著P<0.05和极显著P<0.01).
2 结果与讨论 2.1 根系通气组织不同生物炭湿地中, AC根系的通气组织成像如图 2所示.从中可以直观地看出, 随着生物炭添加量的增加, AC的根系通气组织逐渐增大, 且AC-40和AC-30的根系通气组织明显比其它AC的通气组织更大.

|
图 2 不同ACs的根系通气组织 Fig. 2 Root aerenchyma of different ACs |
经过150 d的运行, 不同生物炭湿地中AC的根孔隙度如图 3所示.相对于AC-K而言, 生物炭湿地植物根孔隙度均有所增高.这与上述各AC的根系通气组织成像图是一致的.由此可知, 生物炭的添加有利于AC根系通气组织的形成, 进而可以增强AC孔隙度.经过单因素方差分析(P﹥0.05)得出, 随着生物碳添加量的增加, AC的根孔隙度显著增加.根据相关性检验可知, AC根孔隙度与生物炭的添加量呈极显著正相关关系(R2=0.994, P<0.01). 40%的生物炭添加比例, 使AC根孔隙度从(10.87±0.71)%增大至(47.13±1.41)%.根孔隙度将直接决定ROL的速率, 高根孔隙度的湿地植物将会将更多的氧气通过植物体进行运输, 导致更多的ROL从根部释放到根际, 进而改变根际氧水平、pH和根部微区氧化还原环境, 对湿地植物适应淹水和胁迫环境具有重要意义[21].

|
图 3 不同ACs的根孔隙度 Fig. 3 Root porosity of different ACs |
不同生物炭湿地AC的琼胶溶液中, O2浓度随时间的变化趋势如图 4所示.在试验周期内, 随着时间的增加, 溶液中的DO浓度逐渐升高, 同时, 还发现不同光照条件下, DO变化规律不同, 说明光照强度对ROL具有影响.值得注意的是, 在空白对照试验中, DO随时间也有增大趋势, 从起始的0.02 mg ·L-1增大至0.06 mg ·L-1, 增大幅度较小.尽管本试验采用石蜡进行液封, 还是会有少量空气穿透进入琼胶溶液.因此, 在计算ROL时将减掉这一部分自然复氧, 以保证结果的可靠性.通过对琼胶溶液中DO随时间变化做线性回归, 得到DO变化速率, 即ROL.湿地植物在不同光照条件下的拟合方程, 决定系数(R2), ROL如表 1所示.

|
图 4 DO浓度随时间变化 Fig. 4 DO concentrations changing over time for different ACs |
|
|
表 1 在不同光照条件下的拟合方程、决定系数和计算ROL Table 1 Fitting equations under different light conditions, determination coefficients and calculated ROLs for different ACs |
除了高生物炭投加比例(40%和30%)在低光照强度下ROL不具有显著差异外, 不同湿地植物的ROL在0、600、1 500和3 700 μmol ·(m2 ·s)-1这4种光照强度下均表现出随生物炭投加量的增大呈显著增加的趋势(图 5).扣除空白对照, 本试验各组湿地中ROL(以鲜重计)平均值AC-40为48.77 nmol ·(h ·g)-1, AC-30为41.27 nmol ·(h ·g)-1, AC-20为32.46 nmol ·(h ·g)-1, AC-10为25.67 nmol ·(h ·g)-1, AC-K为15.69 nmol ·(h ·g)-1.根据光强对5种不同湿地中AC的ROL影响进行单因素方差分析发现, 有光条件下AC的ROL能力显著高于黑暗条件.在3 000 μmol ·(m2 ·s)-1光强条件下, ROL达到最大且显著大于其余光强.虽然在光强<3 000μmol ·(m2 ·s)-1时, 植物的ROL整体呈现随光强增加而增大的趋势, 但当光强>3 000 μmol ·(m2 ·s)-1时, 植物泌氧能力显著下降.这是由于光照对植物生长的影响与温度相似, 随着光强(或温度)的升高均能呈现出先增大后降低的趋势, 存在一个最适光强(或温度).当光强(或温度)超过最适, 则植物体内的叶绿体和线粒体等敏感细胞器会受到损害, 反而不利于植物生长[22].

|
柱状图上不同的字母揭示不同处理的差异性, 其中大写字母揭示组间差异, 小写字母揭示组内差异 图 5 不同光照条件下湿地AC的ROL Fig. 5 ROLs of ACs under different light intensities in the CWs |
植物体内气体的流动需要高压和低压形成压力差, 当植物叶片部分的高压与根际附近的氧低压构成压差时, 根系将表现为较强的泌氧能力[23~25].根据文献[17], 随着生物炭投加量的增加, 植物鲜重和叶片长度增大, 光合色素和可溶性蛋白含量增多, 都有利于植物的光合代谢产生O2.随着叶片中氧分压增大, 利于O2通过通气组织传输至地下部分, 且以ROL的形式扩散至根际[26].
根据光强试验发现, 3 000 μmol ·(m2 ·s)-1应该是AC的较为适宜生长的光照强度, 在该光强下, 植物表现出代谢旺盛, 泌氧能力增大的特点.在该光照强度下, 虽然生物炭湿地中ACs的ROL仍显著大于AC-K, 但由于植物对该光强响应明显, 减弱生物炭的影响, 表现出在生物炭湿地中, 40%和20%的生物炭投加比例差异不显著, 30%和10%的生物炭投加比例差异亦不显著.相关性分析也获得类似的结果, 在0、600和3 700 μmol ·(m2 ·s)-1光强条件下, ROL与生物炭的投加量呈极显著正相关(R12=0.974, R22=0.959, R32=0.966, P<0.01), 而在3 000 μmol ·(m2 ·s)-1光强条件下, ROL与生物炭的投加量呈正相关, 但关系不显著(R2=0.707, P>0.05).
3 结论(1) 通过在传统潜流人工湿地中添加生物炭, 有利于菖蒲形成根系通气组织, 增大菖蒲的根孔隙度.
(2) 在湿地中添加生物炭将间接促进植物的光合代谢产生O2, 利于O2通过通气组织传输至地下部分, 并以根系泌氧的形式扩散至根际.
(3) 在光照强度小于3 000 μmol ·(m2 ·s)-1时, 菖蒲的根系泌氧量整体呈现随生物炭投加量增加而增大的趋势. 3 000 μmol ·(m2 ·s)-1为菖蒲的较为适宜生长的光照强度, 在该光强下, 植物泌氧能力较强, 减弱了生物炭投加比例对植物ROL的影响.
| [1] |
汪健, 李怀正, 甄葆崇, 等. 间歇曝气对垂直潜流人工湿地脱氮效果的影响[J]. 环境科学, 2016, 37(3): 980-987. Wang J, Li H Z, Zhen B C, et al. Effect of intermittent aeration on nitrogen removal efficiency in vertical subsurface flow constructed wetland[J]. Environmental Science, 2016, 37(3): 980-987. |
| [2] | Loreti E, van Veen H, Perata P. Plant responses to flooding stress[J]. Current Opinion in Plant Biology, 2016, 33: 64-71. DOI:10.1016/j.pbi.2016.06.005 |
| [3] | Vretare V, Weisner S E B. Influence of pressurized ventilation on performance of an emergent macrophyte (Phragmites australis)[J]. Journal of Ecology, 2000, 88(6): 978-987. DOI:10.1046/j.1365-2745.2000.00505.x |
| [4] | Srivastava P, Dwivedi S, Kumar N, et al. Performance assessment of aeration and radial oxygen loss assisted cathode based integrated constructed wetland-microbial fuel cell systems[J]. Bioresource Technology, 2017, 244: 1178-1182. DOI:10.1016/j.biortech.2017.08.026 |
| [5] | Armstrong J, Jones R E, Armstrong W. Rhizome phyllosphere oxygenation in Phragmites and other species in relation to redox potential, convective gas flow, submergence and aeration pathways[J]. New Phytologist, 2006, 172(4): 719-731. DOI:10.1111/nph.2006.172.issue-4 |
| [6] | Bedford B L, Bouldin D R, Beliveau B D. Net oxygen and carbon-dioxide balances in solutions bathing roots of wetland plants[J]. Journal of Ecology, 1991, 79(4): 943-959. DOI:10.2307/2261090 |
| [7] | Chen H J, Qualls R G, Miller G C. Adaptive responses of Lepidium latifolium to soil flooding:biomass allocation, adventitious rooting, aerenchyma formation and ethylene production[J]. Environmental and Experimental Botany, 2002, 48(2): 119-128. DOI:10.1016/S0098-8472(02)00018-7 |
| [8] | Li H, Ye Z H, Wei Z J, et al. Root porosity and radial oxygen loss related to arsenic tolerance and uptake in wetland plants[J]. Environmental Pollution, 2011, 159(1): 30-37. DOI:10.1016/j.envpol.2010.09.031 |
| [9] | Lemoine D G, Mermillod-Blondin F, Barrat-Segretain M H, et al. The ability of aquatic macrophytes to increase root porosity and radial oxygen loss determines their resistance to sediment anoxia[J]. Aquatic Ecology, 2012, 46(2): 191-200. DOI:10.1007/s10452-012-9391-2 |
| [10] | Han C, Ren J H, Wang Z D, et al. Characterization of phosphorus availability in response to radial oxygen losses in the rhizosphere of Vallisneria spiralis[J]. Chemosphere, 2018, 208: 740-748. DOI:10.1016/j.chemosphere.2018.05.180 |
| [11] | Deng H, Ye Z H, Wong M H. Lead, zinc and iron (Fe2+) tolerances in wetland plants and relation to root anatomy and spatial pattern of ROL[J]. Environmental and Experimental Botany, 2009, 65(2-3): 353-362. DOI:10.1016/j.envexpbot.2008.10.005 |
| [12] | Wu C, Zou Q, Xue S G, et al. The effect of silicon on iron plaque formation and arsenic accumulation in rice genotypes with different radial oxygen loss (ROL)[J]. Environmental Pollution, 2016, 212: 27-33. DOI:10.1016/j.envpol.2016.01.004 |
| [13] | Williams P N, Santner J, Larsen M, et al. Localized flux maxima of arsenic, lead, and iron around root apices in flooded lowland rice[J]. Environmental Science & Technology, 2014, 48(15): 8498-8506. |
| [14] | Dullo B W, Grootjans A P, Roelofs J G M, et al. Radial oxygen loss by the cushion plant Eriocaulon schimperi prevents methane emissions from an East-African mountain mire[J]. Plant Biology, 2017, 19(5): 736-741. DOI:10.1111/plb.2017.19.issue-5 |
| [15] | Kotula L, Schreiber L, Colmer T D, et al. Anatomical and biochemical characterisation of a barrier to radial O2 loss in adventitious roots of two contrasting Hordeum marinum accessions[J]. Functional Plant Biology, 2017, 44(9): 845-857. DOI:10.1071/FP16327 |
| [16] | Lai W L, Zhang Y, Chen Z H. Radial oxygen loss, photosynthesis, and nutrient removal of 35 wetland plants[J]. Ecological Engineering, 2012, 39: 24-30. DOI:10.1016/j.ecoleng.2011.11.010 |
| [17] |
黄磊, 陈玉成, 赵亚琦, 等. 生物炭添加对湿地植物生长及氧化应激响应的影响[J]. 环境科学, 2018, 39(6): 2904-2910. Huang L, Chen Y C, Zhao Y Q, et al. Influence of biochar application on growth and antioxidative responses of macrophytes in subsurface flow constructed wetlands[J]. Environmental Science, 2018, 39(6): 2904-2910. |
| [18] | Huang L, Chen Y C, Liu G, et al. Non-isothermal pyrolysis characteristics of giant reed (Arundo donax L.) using thermogravimetric analysis[J]. Energy, 2015, 87: 31-40. DOI:10.1016/j.energy.2015.04.089 |
| [19] | Mei X Q, Ye Z H, Wong M H. The relationship of root porosity and radial oxygen loss on arsenic tolerance and uptake in rice grains and straw[J]. Environmental Pollution, 2009, 157(8-9): 2550-2557. DOI:10.1016/j.envpol.2009.02.037 |
| [20] | Jensen S I, Kühl M, Glud R N, et al. Oxic microzones and radial oxygen loss from roots of Zostera marina[J]. Marine Ecology Progress Series, 2005, 293: 49-58. DOI:10.3354/meps293049 |
| [21] | Mei X Q, Yang Y, Tam N F Y, et al. Roles of root porosity, radial oxygen loss, Fe plaque formation on nutrient removal and tolerance of wetland plants to domestic wastewater[J]. Water Research, 2014, 50: 147-159. DOI:10.1016/j.watres.2013.12.004 |
| [22] | Lasfar S, Monette F, Millette L, et al. Intrinsic growth rate:a new approach to evaluate the effects of temperature, photoperiod and phosphorus-nitrogen concentrations on duckweed growth under controlled eutrophication[J]. Water Research, 2007, 41(11): 2333-2340. DOI:10.1016/j.watres.2007.01.059 |
| [23] | Allen L H Jr. Mechanisms and rates of O2 transfer to and through submerged rhizomes and roots via aerenchyma[J]. Annual Proceedings Soil and Crop Science Society of Florida, 1997, 56: 41-54. |
| [24] | Grosse W, Frick H J. Gas transfer in wetland plants controlled by Graham's law of diffusion[J]. Hydrobiologia, 1999, 415: 55-58. DOI:10.1023/A:1003885701458 |
| [25] | Jackson M, Armstrong W. Formation of aerenchyma and the processes of plant ventilation in relation to soil flooding and submergence[J]. Plant Biology, 1999, 1(3): 274-287. DOI:10.1111/plb.1999.1.issue-3 |
| [26] | Cheng X Y, Wang M, Zhang C F, et al. Relationships between plant photosynthesis, radial oxygen loss and nutrient removal in constructed wetland microcosms[J]. Biochemical Systematics and Ecology, 2014, 54: 299-306. DOI:10.1016/j.bse.2014.02.020 |