 2019, Vol. 40
2019, Vol. 40

2. 哈尔滨工业大学城市水资源与水环境国家重点实验室, 哈尔滨 150090;
3. 黑龙江扎龙国家级自然保护区管理局, 齐齐哈尔 161000
 , GAO Wei-feng1 , LIU Feng-qin2 , WANG Wen-feng3 , MA Jian-hua3 , XU Zhuo3 , LIANG Hong2
, GAO Wei-feng1 , LIU Feng-qin2 , WANG Wen-feng3 , MA Jian-hua3 , XU Zhuo3 , LIANG Hong2 , GAO Da-wen1,2
, GAO Da-wen1,2
2. State Key Laboratory of Urban Water Resource and Water Environment, Harbin Institute of Technology, Harbin 150090, China;
3. Heilongjiang Zhalong National Nature Reserve Administration, Qiqihar 161000, China
全球大气中60%~70%的N2O来自于土壤的硝化与反硝化过程[1].湿地作为陆地生态系统的重要组成部分, 其土壤温度、湿度、pH、总有机碳(TOC)和总氮(TN)等土壤理化性质的变化都会影响微生物群落结构及多样性和以微生物为媒介的硝化和反硝化作用, 从而影响湿地N2O的排放[2].当前国内外学者已经在湿地N2O的排放特征、影响因素以及生成机制等方面开展了大量研究[3~5].不同类型湿地由于其影响因子的不同N2O排放也有所差异, 依照地域特点, 典型湿地类型包括:淡水沼泽湿地、典型河口滨海湿地、典型高原湿地以及盐碱湿地.其中淡水沼泽湿地N2O的排放通量一般在非冰冻期表现为以排放为主, 多呈现波动性双峰型变化趋势, 排放峰值常出现在5月和7~8月, 且在有冰雪覆盖时N2O通常会出现微弱的吸收现象[6~9]; 而典型的河口滨海湿地即使在冬季也仍有N2O释放现象, 甚至会在冬季达到N2O排放通量的最大值[10~13]; 与之相比, 典型高原湿地的N2O通量则呈现较大的波动性变化, 一般无明显的时间变化规律[14, 15]; 而作为我国内陆独有的湿地类型——盐碱湿地, 其N2O通量的研究还普遍较少.在研究的各影响因素中土壤水分和土壤温度通常被认为是影响N2O排放的主要控制因素[16, 17].而对于盐碱湿地来说, 其土壤有机质含量相对较少, 且氨氧化菌的丰富度和多样性比较高, 会很大程度地影响微生物硝化作用的强度, 而目前针对于盐碱湿地N2O生成排放中硝化作用对其的影响研究还尚不多见.
扎龙湿地是我国北方典型的盐碱湿地, 位于黑龙江省西部的松嫩平原, 乌裕尔河下游, 占地面积约21万公顷, 是我国北方同纬度地区保留最完善、景观最原始、物种最丰富的沼泽湿地生态系统.芦苇(Phragmites communis)是该地最为典型的植被类型, 且扎龙湿地偏碱性土壤更适宜氨氧化菌的生存, 是潜在的温室气体排放源或汇.目前, 科学家对扎龙湿地芦苇种群的生长特征及影响因子[18, 19]、土壤中营养盐的时空变化规律[20, 21]以及土壤微生物的分布特征[22]都已进行了研究.然而在其温室气体排放方面, 国内学者以往的研究多集中在CH4和CO2上[23, 24], 而对于扎龙湿地氨氧化菌在氨氧化过程中会释放的重要温室气体——N2O的研究目前还鲜见报道.
本文采用静态暗箱-气相色谱法对扎龙芦苇湿地生长季的N2O排放通量特征和影响因素进行了观测和研究, 分析其N2O产生的硝化作用机制, 以便填补扎龙芦苇湿地N2O释放规律的数据空白, 同时也可为探讨硝化作用对盐碱湿地N2O排放的影响, 以及正确评估盐碱湿地在全球温室效应中的作用提供科学的参考依据.
1 材料与方法 1.1 研究区概况研究区位于黑龙江省齐齐哈尔市东南30 km处的扎龙国家级自然保护区内, 地理坐标为47°10'30"N, 124°14' 28"E(图 1), 平均海拔144 m.扎龙湿地属于典型的中温带大陆性季风气候, 年平均气温2~4.2℃, 年均降水量428 mm左右, 且降水多集中在6~8月, 约占全年降水量的67%.试验区地势低洼平坦, 芦苇是其中的优势代表植物, 土壤盐碱性偏高, 地表明显发白, pH值在7.7~8.9之间.

|
图 1 采样点位置示意 Fig. 1 Map of sampling site |
采用静态箱-气相色谱法测定N2O通量.静态箱为50 cm×50 cm×50 cm四面和顶部均封闭的不锈钢材质装置.箱内顶部装有两个小风扇和一个可连接便携式数字温度计的热电偶探头(JM-624), 并设有气样采集孔和风扇接线孔于箱顶; 箱体外部包裹泡沫, 以减少采样时箱内温度的波动.箱底基座采用不锈钢槽架(50 cm×50 cm×20 cm), 采样前数天插入地下20 cm固定.为了减少采样时对基座和周围环境的扰动, 在取样点架设了木制栈桥.采样时间为2017年7月4日至10月29日, 采样频率为每月3次, 约10 d取一次, 每次采样时间为当日的09:00~11:00.试验设计3个平行重复, 同时采集气体样品, 采样时需将底座凹槽注水密封, 用50 mL医用注射器在45 min时间段内每隔15 min采集1次气体样品(共抽取4次气体样品).气体样品储存于铝箔采样袋中, 带回实验室用Shimadzu GC2010气相色谱仪电子捕获检测器(ECD)进行分析, 色谱仪设置如下:柱箱温度70℃, 电子捕获检测器工作温度250℃, 色谱柱采用长度为1 m内径2 mm的80~100目Porapak Q填充物来分离N2O, 载气为高纯氮气, 流速20 mL·min-1.采集气体的同时, 同步测定箱内温度、土壤温度和积水深度.
N2O的排放通量表示单位时间、单位面积观测箱内N2O质量的变化, 可采用下述公式进行计算:
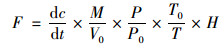
|
式中, F为N2O的排放通量[μg·(m2·h)-1], dc/dt为采样过程中箱内气体浓度随时间的变化率, M为N2O的摩尔质量(g·mol-1), P为采样点气压(Pa), T为采样时绝对温度(K), V0、P0、T0分别为标准状态下的气体摩尔体积(mL·mol-1)、空气气压(Pa)和绝对温度(K), H为水面以上采样箱高度(m).
1.3 土壤样品的采集与测定土壤采集时间为2017年7~10月, 采样频率为每月两次, 采集时间与每月3次气体采集中的两次时间相对应, 在采集气体的同时采集土壤样品, 现场测定5个层次(0、5、20、40、60 cm)土壤温度, 每次采集3个层次(0~20 cm、20~40 cm、40~60 cm)土壤样品.将采集到的土样过2 mm筛以去除其中的石块、植物残根等杂物, 用于相应土壤理化指标和微生物活性的测定.
土壤pH值采用电位法(水土比2.5: 1)测定; 土壤含水率采用105℃烘干比重法测定; 水位深度使用米尺进行现场测量; TOC、TN含量需将样品自然风干, 磨碎后采用元素分析仪检测; NH4+、NO3-含量使用1 mol·L-1 KCl浸提(水土比5: 1), 振荡离心过滤后, 采用连续流动分析仪(Seal Analytical AA3, Norderstedt, Germany)检测; 土壤中氨氧化菌的潜在氨氧化速率(potential ammonia oxidation rate, PAO)需取25 g鲜土样品, 在(25±2)℃, 175 r·min-1下进行避光培养, 采用氯酸盐抑制的方法快速检测(ISO15685, 2012).
1.4 数据统计分析采用线性回归分析N2O通量与土温、水深以及PAO等影响因素的相关关系, 所有数据均采用SPSS 20.0软件进行相关统计分析, 并运用Origin 2016软件进行绘图.
2 结果与分析 2.1 N2O排放特征扎龙芦苇沼泽湿地生长季N2O排放通量具有明显的动态变化特征, 呈现波动性下降变化趋势(图 2).观测期间, N2O通量变化范围为-24.48~91.52 μg·(m2·h)-1, 最大值出现在7月中旬, 最小值出现在生长季末, 此时N2O出现较弱的吸收现象, 但平均通量为(37.49±15.75) μg·(m2·h)-1, 总体上仍然表现为N2O的释放.这表明, 扎龙芦苇湿地生长季为N2O的排放源.在观测初期(7月), N2O排放通量较高, 8月以后逐渐下降, 尤其9~10月通量下降显著且波动性较大, 甚至在10月中下旬出现了负值.对比前人研究的同为盐碱地的白洋淀芦苇湿地[25], 两地区7~8月的N2O通量都明显较高, 之后白洋淀湿地N2O通量虽也下降, 但直至10月末仍有少量N2O释放, 而扎龙湿地在生长季末则出现吸收现象.这可能与不同地区的环境因子差异有关, 使得扎龙芦苇湿地N2O排放变化明显.

|
图 2 扎龙芦苇湿地N2O排放通量(平均值±标准误) Fig. 2 Dynamic change of N2O flux in the Zhalong reed wetland (Mean±SE) |
扎龙芦苇湿地随着土层深度的增加, 土壤温度逐渐降低(图 3).观测初期土壤温度较高, 7月中旬表层土最高土温可达到25.0℃, 且不同土层深度温度变化幅度较大.扎龙湿地0~60 cm整体土壤温度的最高值出现在7月上旬, 平均土温20.2℃, 8月以后土温呈现逐渐下降的趋势, 且不同土层温度差异不大.直至10月末, 土壤温度已大幅度降至6.2℃.

|
图 3 扎龙芦苇湿地不同土层深度土壤温度(平均值±标准误) Fig. 3 Soil temperature at different soil depths of the Zhalong reed wetland (Mean±SE) |
扎龙芦苇湿地在8月中旬出现持续积水情况, 水位深度越来越高, 从最初的3.3 cm逐渐升至9月末的27.0 cm, 其中8月下旬至9月上旬水位涨势最大, 升高了8.5 cm. 10月湿地水位出现小幅度波动回落, 但仍不低于20.0 cm的水位值, 扎龙湿地积水程度较深(图 4).

|
图 4 扎龙芦苇湿地积水深度(平均值±标准误) Fig. 4 Water table depths of the Zhalong reed wetland (Mean±SE) |
扎龙芦苇湿地土壤的TOC和TN含量分别在27.45~51.85 g·kg-1和0.80~2.05 g·kg-1, 其TOC含量要明显高于TN含量, 且二者含量随着土壤深度的增加表现出由表层到底层逐渐降低的趋势(图 5).同时, 土壤TOC和TN含量总体上的变化趋势大致相似, 7、8月二者含量都各自保持相对稳定, 在8月末下降到较低值, 9月以后总碳氮含量有所回升, 至10月末再次下降至低值.有所差异的是, 土壤TOC含量在9月回升后有再升高趋势, 10月上旬扎龙芦苇湿地TOC含量达到了最高值, 0~60 cm土层平均含量48.70 g·kg-1; 而土壤TN含量在9月稍有回升后就再次下降, 至10月末达到了其TN含量的最低值, 0~60 cm土层平均含量0.87 g·kg-1.

|
图 5 扎龙芦苇湿地不同土层深度土壤TOC、TN含量 Fig. 5 Content of TOC and TN in different soil depths of the Zhalong reed wetland |
扎龙芦苇湿地土壤的NH4+-N和NO3--N含量分别在0.48~2.23 mg·kg-1和0.42~0.71 mg·kg-1, NH4+-N含量稍高于NO3--N含量(图 6).其中NH4+-N含量随着土壤深度的增加而逐渐降低, 在7月中旬其含量达到最高值后(0~60 cm土层平均含量1.49 mg·kg-1), NH4+-N含量开始逐渐下降, 至10月中旬含量有所回升后又再次降至最低值; 而NO3--N含量则变化不大, 没有明显的变化趋势, 其含量峰值出现在9月中旬, 0~60 cm土层平均含量0.67 mg·kg-1.

|
图 6 扎龙芦苇湿地不同土层深度土壤NH4+-N、NO3--N含量 Fig. 6 Content of NH4+-N and NO3--N in different soil depths of the Zhalong reed wetland |
氨氧化菌属好氧自养型硝化细菌, 因此会广泛分布于扎龙湿地的上层土壤中(0~20 cm).本试验在观测期内对扎龙芦苇湿地0~20 cm土层的PAO进行了连续测定(图 7), 其波动范围(以NO2-/干土计)为4.40~13.45 μg·(g·h)-1, 氨氧化菌活性最高出现在7月中旬, 最低值在10月末, 整体变化趋势与N2O排放基本保持一致.

|
图 7 扎龙芦苇湿地0~20 cm土层氨氧化菌活性(平均值±标准误) Fig. 7 PAO of 0-20 cm soil layer in the Zhalong reed wetland (Mean±SE) |
土壤温度对产生N2O的生物学过程具有十分重要的影响, 可通过影响硝化和反硝化细菌的活性间接影响N2O的产生速率[26].三江平原小叶章湿地的N2O通量与土壤温度(5 cm)呈显著的线性正相关关系[27, 28].本研究也发现N2O排放与土壤温度关系密切, 对二者的相关分析表明(表 1):扎龙芦苇湿地生长季的N2O通量与0、5、20、40、60 cm的土层温度皆呈显著正相关关系(P<0.05), 这就说明该湿地土壤N2O通量的变化在很大程度上受制于0~60 cm土温的影响.扎龙芦苇湿地观测初期由于夏季温度高, 土壤温度也升高, 微生物代谢活性增强, 从而促进了N2O的产生, 因此在7月出现了N2O的排放峰值.通常土壤温度高于5℃时就适于硝化微生物发挥活性产生N2O[29], 且在15~40℃范围内, 硝化作用随温度的升高而升高[30].本研究8月以后N2O通量随土温降低而下降, 由此可见硝化反应对该地N2O的产生具有很大的影响.另外, 上层土温对N2O通量的影响程度要明显高于深层土, 一方面是因为大气中的N沉降能被浅层土壤更好地吸收[31, 32]; 另一主要原因是由于氨氧化菌为好氧菌, 其在浅层土壤中含量较高, 且植物根系大多分布于土壤上层, 根系分泌物多, 根系泌氧也能在根际形成适合好氧微生物生存的微氧环境, 氨氧化菌更活跃[33, 34].氨氧化菌的活性直接影响硝化作用强弱, 因此, 这也表明在本研究区中硝化作用对N2O排放变化的影响可能更大.
|
|
表 1 N2O排放通量与不同深度土壤温度的关系1) Table 1 Relationship between N2O fluxes and soil temperature at different soil depths |
此外, 湿地水分含量直接关系到土壤的通气状况和O2含量, 对土壤的硝化和反硝化作用强弱产生影响, 从而引起N2O通量的变化.通过对湿地积水深度与N2O排放的相关分析表明(图 8):二者呈显著的线性负相关关系(P<0.05).这种关系也已被许多研究所证实, 当土壤含水量处于饱和含水量以下时, 土壤水分与N2O通量呈现正相关; 当土壤含水量处于饱和含水量以上时, 积水深度与N2O通量存在着一定的负相关性[35].扎龙芦苇湿地8月中旬后的土壤含水量远在饱和含水量之上, 因此自8月水位逐渐升高后, N2O排放量下降.且过高的积水深度会阻碍N2O的排放扩散, 增加微生物吸收N2O进一步还原为N2的几率[36].这也导致了N2O通量的持续下降甚至在生长季末出现了负值.一般认为, 在土壤透气性良好的条件下, 土壤呈氧化状态, 硝化过程为N2O的主要释放源; 而在透气性较差的条件下, 土壤呈还原态, 反硝化会释放N2O[37].扎龙湿地7月的土壤含水量在45%~75%之间, 正处于研究发现的N2O生成与排放的最适土壤水分值[38], 因此有利于硝化产生更多的N2O.之后随着湿地积水的逐渐加深, 土壤透气性较差.理论上, 淹水造成的厌氧环境, 会促进反硝化过程, 从而增加N2O排放比例[39].但研究区的N2O通量一直在降低, 一方面可能是强还原环境使得N2O还原成了N2, 另一方面也有可能是因为硝化作用的减弱造成的.

|
图 8 N2O排放通量与积水深度的关系 Fig. 8 Relationship between N2O fluxes and water table depth |
扎龙芦苇湿地相比三江平原泥炭沼泽湿地[40, 41]、尕海湿地[14]这类酸性土壤地区, 其土壤TOC和TN含量明显要低很多.有研究表明, 当土壤营养元素处于较高水平下, 异养微生物处于竞争优势, 可能会抑制自养微生物生长, 降低自养微生物多样性[42].并且反硝化作用与土壤有机碳含量相关, 有机质含量高可提高反硝化作用[43, 44].因此, 扎龙芦苇湿地较低的有机质含量可能更适宜自养型氨氧化菌的生长, 同时也降低了土壤的反硝化作用.由于硝化作用的初始底物为NH4+-N, 而反硝化为NO3--N, 通过对湿地NH4+-N含量与N2O通量的相关分析表明(图 9):在0~40 cm土层中二者呈显著的线性正相关关系(P<0.05), 这就说明扎龙湿地N2O的排放在一定程度上会受到中上层土壤NH4+-N含量的影响, NH4+-N的硝化过程可以产生N2O[45], 由此可见硝化反应在该地N2O产生过程中起到了重要作用.而对比于NO3--N含量与N2O通量之间则没有显著的相关关系, 表明反硝化对N2O排放影响较小, 可推断在扎龙芦苇湿地N2O释放过程中, 硝化作用的贡献程度要比反硝化强.

|
图 9 N2O排放通量与不同深度土壤NH4+-N含量的关系 Fig. 9 Relationship between N2O fluxes and soil ammonium nitrogen content at different soil depths |
氨氧化菌是硝化作用生成N2O过程中最关键的微生物, 外部环境因子以及营养状况的变化会十分敏感地影响土壤氨氧化菌活性, 从而影响N2O的产生和排放.通过对0~20 cm土层温度与氨氧化菌活性的相关分析表明(图 10):二者呈极显著的正相关关系(P<0.01).一般认为, 氨氧化菌的最适生长温度为20~30℃, 高于或低于此范围氨氧化菌活性都会有所下降[46].本研究中只有在7月上层土壤(0~20 cm)的平均温度在21℃左右, 达到了氨氧化菌的最适温度值, 此时活性也达到了生长期阶段的最高值.之后氨氧化菌活性明显下降, 因为低温会对微生物细胞膜的流动性和生物大分子的活性产生影响, 不利于其正常代谢.氨氧化菌在较低温度时, 其群落结构多样性和活性均有所降低[47].同时, 土壤碳氮含量和O2含量也是影响氨氧化菌活性的重要因素, 因氨氧化菌为自养微生物, 土壤中的有机质不是其碳源, 盐碱湿地过低的有机质含量可能会增加氨氧化菌的数量, 增强土壤的硝化作用.但硝化作用的正常进行需要消耗大量氧气, 且土壤水分会通过对O2含量的影响来影响氨氧化菌活性[48].研究区8月中旬后越来越高的积水深度导致土壤处于极度厌氧状态, 致使硝化作用减弱, 氨氧化菌活性降低.综合分析, 7月中旬由于上述土壤温度、含水率以及碳氮含量等各方面综合作用达到最适状态, 氨氧化菌活性达到最高, 从而促进了扎龙湿地N2O排放达到了最大值, 之后氨氧化菌活性降低, N2O通量也随之下降.

|
图 10 0~20 cm土层氨氧化菌活性与土壤温度关系 Fig. 10 10 Relationship between PAO and soil temperature in the 0-20 cm layer |
PAO是指土壤或沉积物中参与氨氧化作用的微生物在饱和基质浓度下将氨氮氧化的最大能力[49].由于弱碱性湿地氨氧化菌种群的丰富度和多样性要显著高于酸性湿地[22], 扎龙湿地此类的偏碱环境更适宜氨氧化菌的生存.氨氧化菌在氨单加氧酶(ammonia monooxygenase, AMO)和羟胺氧化还原酶(hydroxylamine oxidoreductase, HAO)的催化作用下可以将铵态氮转化为亚硝态氮, 该过程是硝化作用的第一步, 也是限速步骤[50].因此对于PAO的测定能够在一定程度上反映出土壤中微生物的生态功能及其硝化活性的强弱.同时, 在这一氨氧化过程中, 会生成中间产物羟胺(NH2OH), 其化学分解释放N2O[51].通过对PAO和N2O通量进行相关分析表明(图 11):二者呈极显著的线性正相关关系(P<0.001), 相关系数为0.949.这就说明在扎龙芦苇沼泽湿地生态系统中, N2O的产生和排放极大程度地受制于氨氧化菌活性的影响, 进一步表明了硝化反应对此类盐碱湿地N2O的释放产生了巨大作用.

|
图 11 N2O排放通量与0~20 cm土层氨氧化菌活性关系 Fig. 11 Relationship between N2O fluxes and PAO in the 0-20 cm soil layer |
(1) 扎龙芦苇沼泽湿地生长季N2O的排放通量呈现波动性下降趋势, 变化范围为-24.48~91.52 μg·(m2·h)-1, 最大值出现在7月中旬, 整体平均通量为(37.49±15.75) μg·(m2·h)-1, 表现为大气N2O的排放源.
(2) 土壤温度、积水深度和养分含量都会影响N2O排放, 扎龙芦苇湿地N2O排放通量与不同深度的土层温度皆呈显著正相关关系(P<0.05), 且上层土壤温度对N2O排放通量的影响程度要明显高于深层土; 而与积水深度则呈显著负相关关系(P<0.05).且湿地TOC和TN含量相对较低, N2O通量与0~40 cm土层的NH4+-N含量呈显著正相关关系, 而与NO3--N含量没有关系, 硝化作用程度要比反硝化强.
(3) 扎龙芦苇沼泽湿地氨氧化菌活性受外部环境因子敏感影响, 其土壤氨氧化菌活性与土壤温度呈极显著正相关关系(P<0.01), 且活性变化趋势与N2O排放通量相似, 相关分析表明二者存在极显著正相关关系(P<0.001), 进一步表明了硝化反应对此类盐碱湿地N2O的释放产生了巨大作用.
| [1] | Syakila A, Kroeze C. The global nitrous oxide budget revisited[J]. Greenhouse Gas Measurement and Management, 2011, 1(1): 17-26. DOI:10.3763/ghgmm.2010.0007 |
| [2] | Gödde M, Conrad R. Influence of soil properties on the turnover of nitric oxide and nitrous oxide by nitrification and denitrification at constant temperature and moisture[J]. Biology and Fertility of Soils, 2000, 32(2): 120-128. DOI:10.1007/s003740000247 |
| [3] | Kachenchart B, Jones D L, Gajaseni N, et al. Seasonal nitrous oxide emissions from different land uses and their controlling factors in a tropical riparian ecosystem[J]. Agriculture, Ecosystems & Environment, 2012, 158: 15-30. |
| [4] | Audet J, Hoffmann C C, Andersen P M, et al. Nitrous oxide fluxes in undisturbed riparian wetlands located in agricultural catchments:Emission, uptake and controlling factors[J]. Soil Biology and Biochemistry, 2014, 68: 291-299. DOI:10.1016/j.soilbio.2013.10.011 |
| [5] |
王立新, 吴忠伟, 刘华民, 等. 2013年乌梁素海湖滨带土壤-植物系统甲烷和一氧化二氮通量及生态系统总呼吸研究[J]. 湿地科学, 2016, 14(6): 763-772. Wang L X, Wu Z W, Liu H M, et al. Fluxes of methane and nitrous oxide and total ecosystem respiration of soil and plant system in lakeshore of Wuliangsu Lake in 2013[J]. Wetland Science, 2016, 14(6): 763-772. |
| [6] | Augustin J, Merbach W, Rogasik J. Factors influencing nitrous oxide and methane emissions from minerotrophic fens in northeast Germany[J]. Biology and Fertility of Soils, 1998, 28(1): 1-4. DOI:10.1007/s003740050455 |
| [7] | Zhu R B, Liu Y S, Ma J, et al. Nitrous oxide flux to the atmosphere from two coastal tundra wetlands in eastern Antarctica[J]. Atmospheric Environment, 2008, 42(10): 2437-2447. DOI:10.1016/j.atmosenv.2007.12.016 |
| [8] |
宋长春, 张丽华, 王毅勇, 等. 淡水沼泽湿地CO2、CH4和N2O排放通量年际变化及其对氮输入的响应[J]. 环境科学, 2006, 27(12): 2369-2375. Song C C, Zhang L H, Wang Y Y, et al. Annual dynamics of CO2, CH4, N2O emissions from freshwater marshes and affected by nitrogen fertilization[J]. Environmental Science, 2006, 27(12): 2369-2375. DOI:10.3321/j.issn:0250-3301.2006.12.001 |
| [9] |
王毅勇, 郑循华, 宋长春, 等. 三江平原典型沼泽湿地氧化亚氮通量[J]. 应用生态学报, 2006, 17(3): 493-497. Wang Y Y, Zheng X H, Song C C, et al. N2O flux in typical wetlands of Sanjiang Plain[J]. Chinese Journal of Applied Ecology, 2006, 17(3): 493-497. DOI:10.3321/j.issn:1001-9332.2006.03.027 |
| [10] | Inubushi K, Furukawa Y, Hadi A, et al. Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use change in tropical peatlands located in coastal area of South Kalimantan[J]. Chemosphere, 2003, 52(3): 603-608. DOI:10.1016/S0045-6535(03)00242-X |
| [11] |
胡泓, 王东启, 李杨杰, 等. 崇明东滩芦苇湿地温室气体排放通量及其影响因素[J]. 环境科学研究, 2014, 27(1): 43-50. Hu H, Wang D Q, Li Y J, et al. Greenhouse gases fluxes at Chongming Dongtan Phragmites australis wetland and the influencing factors[J]. Research of Environmental Sciences, 2014, 27(1): 43-50. |
| [12] |
许鑫王豪, 邹欣庆, 刘晶茹. 苏北潮滩温室气体排放的时空变化及影响因素[J]. 环境科学, 2016, 37(6): 2383-2392. Xu X W H, Zou X Q, Liu J R. Temporal and spatial dynamics of greenhouse gas emissions and its controlling factors in a coastal saline wetland in North Jiangsu[J]. Environmental Science, 2016, 37(6): 2383-2392. |
| [13] |
谢文霞, 赵全升, 张芳, 等. 胶州湾河口湿地秋冬季N2O气体排放通量特征[J]. 地理科学, 2011, 31(4): 464-469. Xie W X, Zhao Q S, Zhang F, et al. Characteristics of N2O flux in estuary wetland of Jiaozhou Bay in autumn and winter[J]. Scientia Geographica Sinica, 2011, 31(4): 464-469. |
| [14] |
马维伟, 王辉, 李广, 等. 尕海湿地CH4、CO2和N2O通量特征初步研究[J]. 草业学报, 2015, 24(8): 1-10. Ma W W, Wang H, Li G, et al. A preliminary study of carbon dioxide, methane and nitrous oxide fluxes from the Gahai wetland[J]. Acta Prataculturae Sinica, 2015, 24(8): 1-10. |
| [15] |
王德宣, 宋长春, 王跃思, 等. 若尔盖高原沼泽湿地N2O排放通量研究[J]. 生态科学, 2005, 24(3): 193-196. Wang D X, Song C C, Wang Y S, et al. N2O flux from the mire in Ruoergai Plateau[J]. Ecologic Science, 2005, 24(3): 193-196. DOI:10.3969/j.issn.1008-8873.2005.03.001 |
| [16] | Lohila A, Aurela M, Hatakka J, et al. Responses of N2O fluxes to temperature, water table and N deposition in a northern boreal fen[J]. European Journal of Soil Science, 2010, 61(5): 651-661. DOI:10.1111/j.1365-2389.2010.01265.x |
| [17] | Regina K, Nykänen H, Silvola J, et al. Fluxes of nitrous oxide from boreal peatlands as affected by peatland type, water table level and nitrification capacity[J]. Biogeochemistry, 1996, 35(3): 401-418. DOI:10.1007/BF02183033 |
| [18] |
焦德志, 荣子, 曹瑞, 等. 扎龙湿地保护区异质生境芦苇种群分株构件的数量特征[J]. 生态学报, 2017, 37(23): 7843-7853. Jiao D Z, Rong Z, Cao R, et al. Quantitative characteristics of the ramet module of Phragmites australis populations in heterogeneous habitats in the Zhalong Wetland National Reserve[J]. Acta Ecologica Sinica, 2017, 37(23): 7843-7853. |
| [19] |
杨晓杰, 佟守正, 李旭业, 等. 扎龙湿地芦苇群落生长特征对水深梯度变化的响应[J]. 东北林业大学学报, 2012, 40(12): 67-70. Yang X J, Tong S Z, Li X Y, et al. Response of growth feature of reed communities in Zhalong wetland to the change of water depth gradient[J]. Journal of Northeast Forestry University, 2012, 40(12): 67-70. DOI:10.3969/j.issn.1000-5382.2012.12.016 |
| [20] |
张静, 马玲, 丁新华, 等. 扎龙湿地不同生境土壤微生物生物量碳氮的季节变化[J]. 生态学报, 2014, 34(13): 3712-3719. Zhang J, Ma L, Ding X H, et al. Seasonal dynamics of soil microbial biomass C and N in different habitats in Zhalong wetland[J]. Acta Ecologica Sinica, 2014, 34(13): 3712-3719. |
| [21] |
叶华香, 臧淑英, 肖海丰, 等. 扎龙湿地表层沉积物营养盐空间分布特征及评价[J]. 自然资源学报, 2013, 28(11): 1966-1976. Ye H X, Zang S Y, Xiao H F, et al. Spatial distribution and evaluation of nutrients in surface sediments of Zhalong wetland[J]. Journal of Natural Resource, 2013, 28(11): 1966-1976. DOI:10.11849/zrzyxb.2013.11.013 |
| [22] |
谢月.两种典型东北寒温带沼泽湿地中氨氧化微生物种群的分布特征研究[D].哈尔滨: 哈尔滨工业大学, 2017. Xie Y. Study on the distribution characteristics of ammonia-oxidizing microbes population of two typical marsh wetland soil in northeast cool temperate zone[D]. Harbin: Harbin Institute of Technology, 2017. |
| [23] |
黄璞祎, 于洪贤, 柴龙会, 等. 扎龙芦苇湿地生长季的甲烷排放通量[J]. 应用生态学报, 2011, 22(5): 1219-1224. Huang P Y, Yu H X, Chai L H, et al. Methane emission flux of Zhalong Phragmites australis wetlands in growth season[J]. Chinese Journal of Applied Ecology, 2011, 22(5): 1219-1224. |
| [24] |
杨荐, 关爱洁, 陈碧琪, 等. 人为补水对扎龙芦苇湿地甲烷排放的影响[J]. 安徽农业科学, 2014, 42(22): 7456-7458. Yang J, Guan A J, Chen B Q, et al. Effects of artificial recharge on methane emission from Zhalong reed wetland[J]. Journal of Anhui Agricultural Sciences, 2014, 42(22): 7456-7458. DOI:10.3969/j.issn.0517-6611.2014.22.060 |
| [25] |
牛翠云, 王树涛, 郭艳杰, 等. 白洋淀湿地芦苇型水陆交错带土壤氮素形态变化和N2O排放特征及氮储量研究[J]. 河北农业大学学报, 2017, 40(3): 72-79. Niu C Y, Wang S T, Guo Y J, et al. Studies on variation characteristics of soil nitrogen forms, nitrous oxide emission and nitrogen storage of the Phragmites australis-dominated land/inland water ecotones in Baiyangdian wetland[J]. Journal of Agricultural University of Hebei, 2017, 40(3): 72-79. |
| [26] | 张丽华.氮输入对沼泽湿地碳收支的影响[D].长春: 中国科学院东北地理与农业生态研究所, 2007. |
| [27] | Yu J B, Liu J S, Wang J D, et al. Nitrous oxide emission from Deyeuxia angustifolia freshwater marsh in northeast China[J]. Environmental Management, 2007, 40(4): 613-622. DOI:10.1007/s00267-006-0349-9 |
| [28] |
孙志高, 刘景双, 杨继松, 等. 生长季与非生长季小叶章湿地N2O通量特征及排放贡献[J]. 草业学报, 2009, 18(6): 242-247. Sun Z G, Liu J S, Yang J S, et al. N2O flux characteristics and emission contributions of Calamagrostis angustifolia wetland during growth and non-growth seasons[J]. Acta Prataculturae Sinica, 2009, 18(6): 242-247. DOI:10.3321/j.issn:1004-5759.2009.06.033 |
| [29] | Agehara S, Warncke D D. Soil moisture and temperature effects on nitrogen release from organic nitrogen sources[J]. Soil Science Society of America Journal, 2005, 69(6): 1844-1855. DOI:10.2136/sssaj2004.0361 |
| [30] |
郑循华, 王明星, 王跃思, 等. 温度对农田N2O产生与排放的影响[J]. 环境科学, 1997, 18(5): 1-5. Zheng X H, Wang M X, Wang Y S, et al. Impacts of temperature on N2O production and emission[J]. Environmental Science, 1997, 18(5): 1-5. |
| [31] | Pihlatie M, Pumpanen J, Rinne J, et al. Gas concentration driven fluxes of nitrous oxide and carbon dioxide in boreal forest soil[J]. Tellus B:Chemical and Physical Meteorology, 2007, 59(3): 458-469. DOI:10.1111/j.1600-0889.2007.00278.x |
| [32] | Pihlatie M K, Kiese R, Brüggemann N, et al. Greenhouse gas fluxes in a drained peatland forest during spring frost-thaw event[J]. Biogeosciences, 2010, 7(5): 1715-1727. DOI:10.5194/bg-7-1715-2010 |
| [33] | Gilbert B, Frenzel P. Rice roots and CH4 oxidation:the activity of bacteria, their distribution and the microenvironment[J]. Soil Biology and Biochemistry, 1998, 30(14): 1903-1916. DOI:10.1016/S0038-0717(98)00061-3 |
| [34] | Trias R, Ruiz-Rueda O, García-Lledó A, et al. Emergent macrophytes act selectively on ammonia-oxidizing bacteria and archaea[J]. Applied and Environmental Microbiology, 2012, 78(17): 6352-6356. DOI:10.1128/AEM.00919-12 |
| [35] | Rudaz A O, Wälti E, Kyburz G, et al. Temporal variation in N2O and N2 fluxes from a permanent pasture in Switzerland in relation to management, soil water content and soil temperature[J]. Agriculture, Ecosystems & Environment, 1999, 73(1): 83-91. |
| [36] |
陈海燕, 李虎, 王立刚, 等. 京郊典型设施蔬菜地N2O排放规律及影响因素研究[J]. 中国土壤与肥料, 2012(5): 5-10. Chen H Y, Li H, Wang L G, et al. Characteristics and influencing factors on nitrous oxide emissions from typical greenhouse vegetable fields in Beijing suburbs[J]. Soils and Fertilizers Sciences in China, 2012(5): 5-10. |
| [37] |
于君宝, 刘景双, 孙志高, 等. 中国东北区淡水沼泽湿地N2O和CH4排放通量及主导因子[J]. 中国科学D辑:地球科学, 2009, 39(2): 177-187. Yu J B, Liu J S, Sun Z G, et al. The fluxes and controlling factors of N2O and CH4 emissions from freshwater marsh in Northeast China[J]. Science China Earth Sciences, 2009, 39(2): 177-187. |
| [38] |
颜晓元, 施书莲, 杜丽娟, 等. 水分状况对水田土壤N2O排放的影响[J]. 土壤学报, 2000, 37(4): 482-489. Yan X Y, Shi S L, Du L J, et al. N2O emission from paddy soil as affected by water regime[J]. Acta Pedologica Sinica, 2000, 37(4): 482-489. DOI:10.3321/j.issn:0564-3929.2000.04.007 |
| [39] | Smith K A, Ball T, Conen F, et al. Exchange of greenhouse gases between soil and atmosphere:interactions of soil physical factors and biological processes[J]. European Journal of Soil Science, 2003, 54(4): 779-791. DOI:10.1046/j.1351-0754.2003.0567.x |
| [40] |
孙志高, 刘景双, 王金达, 等. 三江平原小叶章湿地开垦前后N2O通量特征与影响因素分析[J]. 山东农业大学学报(自然科学版), 2007, 38(3): 443-449, 456. Sun Z G, Liu J S, Wang J D, et al. N2O flux characteristics and its affecting factors before and after reclamation of Calamagrostis angustifolia wetland in Sanjiang Plain[J]. Journal of Shandong Agricultural University (Natural Science), 2007, 38(3): 443-449, 456. DOI:10.3969/j.issn.1000-2324.2007.03.027 |
| [41] |
朱晓艳, 宋长春, 郭跃东, 等. 三江平原泥炭沼泽湿地N2O排放通量及影响因子[J]. 中国环境科学, 2013, 33(12): 2228-2234. Zhu X Y, Song C C, Guo Y D, et al. N2O emissions and its controlling factors from the peatlands in the Sanjiang Plain[J]. China Environmental Science, 2013, 33(12): 2228-2234. |
| [42] | Wei S, Norton J M. Microbial control of nitrate concentrations in an agricultural soil treated with dairy waste compost or ammonium fertilizer[J]. Soil Biology and Biochemistry, 2000, 32(10): 1453-1457. DOI:10.1016/S0038-0717(00)00050-X |
| [43] | Guo J P, Zhou C D. Greenhouse gas emissions and mitigation measures in Chinese agroecosystems[J]. Agricultural and Forest Meteorology, 2007, 142(2-4): 270-277. DOI:10.1016/j.agrformet.2006.03.029 |
| [44] |
朱永官, 王晓辉, 杨小茹, 等. 农田土壤N2O产生的关键微生物过程及减排措施[J]. 环境科学, 2014, 35(2): 792-800. Zhu Y G, Wang X H, Yang X R, et al. Key microbial processes in nitrous oxide emissions of agricultural soil and mitigation strategies[J]. Environmental Science, 2014, 35(2): 792-800. |
| [45] | Wrage N, Van Groenigen J W, Oenema O, et al. A novel dual-isotope labelling method for distinguishing between soil sources of N2O[J]. Rapid Communications in Mass Spectrometry, 2005, 19(22): 3298-3306. DOI:10.1002/(ISSN)1097-0231 |
| [46] |
王新, 姚晓红, 吴逸飞, 等. 氨氧化细菌的筛选鉴定及其最适反应条件研究[J]. 浙江农业学报, 2015, 27(6): 1042-1047. Wang X, Yao X H, Wu Y F, et al. Screening and identification of ammonia oxidizing bacteria and optimization of ammoxidation conditions[J]. Acta Agriculturae Zhejiangensis, 2015, 27(6): 1042-1047. DOI:10.3969/j.issn.1004-1524.2015.06.24 |
| [47] | Urakawa H, Tajima Y, Numata Y, et al. Low temperature decreases the phylogenetic diversity of ammonia-oxidizing archaea and bacteria in aquarium biofiltration systems[J]. Applied and Environmental Microbiology, 2008, 74(3): 894-900. DOI:10.1128/AEM.01529-07 |
| [48] | Sliekers A O, Haaijer S C M, Stafsnes M H, et al. Competition and coexistence of aerobic ammonium-and nitrite-oxidizing bacteria at low oxygen concentrations[J]. Applied Microbiology and Biotechnology, 2005, 68(6): 808-817. DOI:10.1007/s00253-005-1974-6 |
| [49] | O'Sullivan C A, Wakelin S A, Fillery I R P, et al. Factors affecting ammonia-oxidising microorganisms and potential nitrification rates in southern Australian agricultural soils[J]. Soil Research, 2013, 51(3): 240-252. DOI:10.1071/SR13039 |
| [50] | Wei B, Yu X, Zhang S T, et al. Comparison of the community structures of ammonia-oxidizing bacteria and archaea in rhizoplanes of floating aquatic macrophytes[J]. Microbiological Research, 2011, 166(6): 468-474. DOI:10.1016/j.micres.2010.09.001 |
| [51] | Frame C H, Casciotti K L. Biogeochemical controls and isotopic signatures of nitrous oxide production by a marine ammonia-oxidizing bacterium[J]. Biogeosciences, 2010, 7(9): 2695-2709. DOI:10.5194/bg-7-2695-2010 |