 2018, Vol. 39
2018, Vol. 39

2. 夸祖鲁-纳塔尔大学, 农业、地球与环境科学学院, 南非斯科茨维尔 3209, 南非
 , WANG Yue-man1 , HOU Peng-fu1 , YANG Lin-zhang1 , Alfred Oduor Odindo2 , XUE Li-hong1
, WANG Yue-man1 , HOU Peng-fu1 , YANG Lin-zhang1 , Alfred Oduor Odindo2 , XUE Li-hong1
2. School of Agricultural, Earth & Environmental Sciences, University of KwaZulu-Natal, Scottsville 3209, South Africa
水体富营养化是中国湖泊河流面临的主要污染问题之一[1, 2].根据2010年发布的“全国污染源普查公报”数据显示, 农业生产(含种植业、禽畜养殖业及水产养殖业)过程对水体氮的贡献率已高于50%, 超过工业生产与生活排放成为威胁水环境健康的主要来源[3].在我国人多地少的严峻土地利用情势下, 种植业的稳定输出是保障国家稳定及安全的重要物质基础, 因此种植业在农业生产中处于最为重要且不可替代的地位[4].大量调研数据显示, 农田养分的过量投入是造成水体富营养化的主要原因之一[2, 5~7].因此, 解决水体富营养化问题离不开种植业的养分调控.如何建立连接富营养化水体养分与农田养分的枢纽, 实现养分资源再利用, 是减少农田养分投入并有效缓解水体富营养化的难点.
在诸多农业面源排放的污染物中, 氮是最主要且最难以防治的污染物, 也是目前水体富营养化问题控制的重点[8].水体环境中的氮素多以离子形态存在.以稻季为例, 受降水或灌溉的影响, 化肥氮多以铵态氮形态经由农田地表径流、农田排水和淋失等过程进入水体[9~11], 而铵态氮本身又是水稻生长所必须的氮素形态.将进入水体环境的铵态氮迁移并实现稻田回用需要适宜载体的介入.这个载体材料不仅要能够高效吸附水体铵离子, 更要保证被吸附铵离子的生物有效性, 且应具备环境友好性及易得性.生物炭来源于农业废弃物的裂解, 其多孔的结构及表面的基团[12, 13]为离子的吸附提供了条件.已有研究证明, 每g生物炭对氮的吸附量可以达到0.85~44.24 mg[14~16], 且Taghizadeh-Toosi等[17]发现植物可以再次利用生物炭吸附氨气, 证明了生物炭固持氮的生物有效性.但是, 作为一个具有较好吸附性且能稳定固持氮的材料, 进入土壤中的生物炭是否会与作物产生氮素争夺, 从而影响作物对氮的吸收利用; 到底有多少生物炭固持的氮素是可以被作物利用的; 如果将生物炭固持氮代替化肥施用农田, 这种外源氮添加方式与传统化肥施用对作物生长是否有显著影响.关于这些涉及实际应用的疑问, 仍然十分缺乏试验数据支撑, 而这些疑问本身是对回答“生物炭能否成为合适载体实现氮素还田”以及“如何使用生物炭进行氮素再利用”的关键.
作物对养分的吸收与利用很大程度取决于作物根系在土壤中的生长和形态, 且作物根系形态与地上部分生物量及养分的累积有着紧密的关系[18, 19].另一方面, 根系形态等特征又是其对土壤环境中养分状况的响应结果[20].有研究表明, 相比于其他营养元素, 氮素对作物根系生长形态的影响最为显著, 也较为复杂[21~23].较高氮素供应条件下, 作物根系及地上部分的生物量均会有所增加, 但根冠比会呈现降低趋势, 且根系更为纤细[24]; 较低氮素供应条件下, 当仅达到轻微缺失时, 根系长度显著增加, 根冠比增加[25]; 但是严重缺氮会使整个植株生长受到抑制.由此可见, 在考察作物地上部分生长及养分累积的同时结合根系形态及根系部分的养分累积情况开展研究, 能够更好体现作物对土壤环境中氮素的响应, 以此为前提评价添加外源氮后土壤供氮能力也会更加准确.
目前, 基于生物炭添加对作物根系影响的研究屡见不鲜, 多数研究给出了正面的研究结果, 如增加根尖数、根系体积、面积及总长等[26~28].但是, 对作物根系生长的促进作用或出现根冠比的增加, 是否有可能源于生物炭添加而带来的对养分供应的限制, 并未开展深入探讨[29]; 有关生物炭添加对作物地上、地下部分养分供应关系的影响, 也十分有限[30]; 而将生物炭附载氮作为氮源供给作物生长, 研究作物地上、地下部分对不同氮源添加方式的响应, 还尚未见报道.
因此, 本研究以水稻为目标作物, 构建两种生境条件:即不同生境中两种氮源添加方式、同一生境中两种氮源添加方式, 分别比较水稻地上、地下部分生长及氮素分配情况对两种氮源添加方式的响应, 分析水稻对两种氮源添加方式的利用差异, 通过对使用生物炭负载氮替代部分化肥投入进行初探, 以期为基于生物炭负载的养分还田提供可行性数据支持.
1 材料与方法 1.1 供试土壤、作物及生物炭材料 1.1.1 供试土壤本研究供试土壤取自江苏宜兴市丁蜀镇漳渎村(N 31°18′24″, E119°54′41″).该地区位于长江三角洲太湖流域, 属亚热带湿润季风气候, 年均温15.7℃, 平均降水1 100 mm.该区域水稻土多由湖底沉积物发育而成, 耕作层深度18 cm, 经历过百年水稻耕作, 耕作层土壤基本理化性质如下:有机质24.80 g·kg-1, 总氮1.79 g·kg-1, 总磷1.62 g·kg-1, 硝态氮17.10 mg·kg-1, 铵态氮23.3 mg·kg-1, Olsen磷48.90 mg·kg-1, 有效钾108.8 mg·kg-1, 阳离子交换量13.40 cmol·kg-1, pH 6.65.
1.1.2 供试作物本研究供试水稻品种为南粳46号.该品种由江苏省农业科学院粮食作物研究所杂交选育而成的晚粳, 具有株型紧凑, 分蘖力强, 抗逆性强(耐旱、耐湿)等特点, 是供试土壤采集地乃至太湖流域的主要水稻栽种品种之一.
1.1.3 供试生物炭材料本研究供试生物炭由小麦秸秆450℃厌氧煅烧而成.煅烧后, 去除表面灰分, 使用0.5 mol·L-1 HCl, 0.1 mol·L-1 NaOH轮流浸泡清洗, 再用去离子水清洗多次, 烘干(80℃)至恒重, 并过40目筛(0.25mm), 备用.生物炭材料理化属性如下:总碳503.10 mg·g-1、总氮12.52 mg·g-1、铵态氮1.82 mg·kg-1、硝态氮3.00 mg·kg-1、pH 8.9、比表面积(BET)7.37 m2·g-1.
1.2 生物炭氮素的负载本研究在前期工作基础上, 选定200 mg·L-1的氯化铵溶液作为负载溶液.为更好确定生物炭负载氮在土壤中的去向, 负载溶液15N标记丰度为10%氯化铵药品配置而成.干燥生物炭按照1 g生物炭:100 mL溶液的配比投放, 搅拌24 h后过滤, 风干48 h.生物炭上氮素负载量(以N计)通过使用2 mol·L-1氯化钾溶液浸提而直接测的, 为(3.40±0.41)mg·g-1.
1.3 试验设计本试验于2017年6~8月在江苏省农业科学院大棚开展.
依据水稻插秧行间距尺寸, 试验使用长25 cm, 宽20 cm, 高20 cm的PVC材质根箱(用于一穴水稻种植), 且设置分区及不分区两种形式(底端设有排水孔).设置4个处理:氮源以氯化铵溶液(非15N标记氯化铵配制)形式直接添加(SN); 氮源以生物炭负载氮形式添加(BN); 根箱两侧分别使用氯化铵溶液及生物炭负载氮作为氮源添加(SN vs. BN, 其中SN vs. BN SN为使用氯化铵溶液侧, SN vs. BN BN为使用生物炭负载氮侧); 无氮添加(CK).为避免根箱分区不同对试验结果产生差异, 本研究在SN及BN处理下分别设置分区及不分区两种根箱(如图 1).每种根箱及处理条件3个重复, 共计18个根箱.

|
图 1 试验处理及根箱设置 Fig. 1 Design of study treatments and pots |
于2017年6月20日向根箱中填放干土12 kg·pot-1(即2400 kg·hm-2), 分区根箱则两侧各填放6 kg干土; 对于以生物炭负载氮形式添加氮源的处理(或部分), 按照土壤重量1%拌入负载氮素的生物炭(相当于添加氮素408 mg·pot-1, 即81.60 kg·hm-2); 对于以氯化铵溶液形式添加氮源的处理(或部分)直接添加等量氮素.此后, 多次加入去离子水待土壤完全浸润(约3 h), 淹水至距土层5 cm高度.填土淹水次日(2017年6月21日), 每个根箱中移栽1穴(含2~3株, 偶数根尖数)水稻幼苗, 用海绵侧向包裹固定.对于分区根箱, 移栽时用少量海绵将每穴根尖平均分之两侧, 侧向包裹固定同上.移栽后1~30 d及38~60 d, 每天加水保持水层高度, 31~37 d打开根箱底部排水孔自然落干.移栽60 d后(2017年8月19日)收获.
1.4 样品采集, 测定及相关计算 1.4.1 作物, 土壤, 田面水样品的采集移栽后第60 d, 测量田面水高度并搅拌水层, 分别从各个根箱或分区根箱的两侧获取田面水样品.倒扣根箱以取出完整作物植株, 并将作物地上、地下部分分离.地上部分样品的获取以根箱为单位, 擦干捆扎后备用; 作物地下部分样品的获取以分区为单位, 即不分区根箱中的作物地下部分收集为1个样品, 分区根箱中作物地下部分按分区进行收集为2个样品.分离黏附在作物地下部分的大块土壤后, 将每个地下部分样品分别套在100目的筛网袋中, 浸泡12 h, 冲洗至根系表面无可见土壤颗粒, 使用镊子将根系进行分离并浸泡在去离子水中备用.收集筛网袋中散落的土壤, 作为根系土壤样品; 同时收集根箱各分区的剩余土壤, 作为非根系土壤样品.
1.4.2 根系形态使用扫描仪(Epson1680)及根系分析程序(WinRhizo Pro 2012)对洗净的根系样品进行扫描和分析后, 得到总根长, 根表面积, 根体积及平均直径等相关根系形态指标.在此基础上, 根据根直径大小, 将总根长, 根表面积, 根体积的数据进行分类, 分为不定根(D>0.3 mm)、粗分枝根(0.1 mm<D≤0.3 mm)、细分枝根(D≤0.1 mm).
1.4.3 作物生物量, 氮累积量及氮素利用率作物地上部分及扫描过根系的地下部分样品, 在105℃的烘箱中放置30 min进行杀青, 然后转入80℃环境下烘干至恒重(约24 h).作物地上、地下部分分别称重, 按照1穴·pot-1, 200 000穴·hm-2折算后, 记作地上、地下部分干物质重(DMA和DMU, t·hm-2; CK处理为DMA0和DMU0, t·hm-2).研磨过100目筛后, 用凯氏定氮仪(Foss Scino KT260, China)分别测定其氮素含量(CNA和CNU, kg·t-1; CK处理为CNA0和CNU0, kg·t-1).氮施用量(NAR, kg·hm-2)为81.60 kg·hm-2.
作物地上部分氮累积量(kg·hm-2):
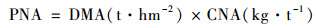
|
(1) |
CK处理地上部分氮累积量为(kg·hm-2):
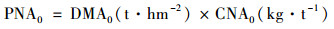
|
(2) |
作物地下部分氮累积量(kg·hm-2):
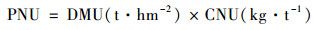
|
(3) |
CK处理地下部分氮累积量为(kg·hm-2):
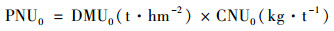
|
(4) |
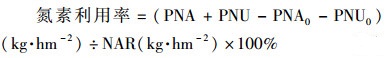
|
(5) |
土壤样品按根系及非根系样品分类后, 按照溶液:鲜土(10 mL:1 g)比例(R, L·kg-1, R=10)使用2 mol·L-1 KCl溶液浸泡, 200 r·min-1往复振荡1 h, 静置后过滤.滤液经流动分析仪(Skalar Corp., Netherlands)测得其中矿质态氮含量(CMN, mg·L-1).鲜土风干, 测得含水率(M, %).根系及非根系鲜土重量分别为m及(2 400-m)(kg·hm-2).
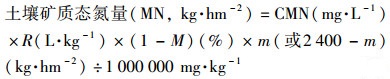
|
(6) |
KCl浸提后的土壤使用去离子水浸泡冲洗多次, 晾干研磨过100目, 使用元素分析仪(Flash EA 1112 series; Thermo Finnigan, USA)测定其中有机态氮含量(CON, g·kg-1).

|
(7) |
田面水高度(h, m), 田面水样品过滤后, 滤液经流动分析仪(Skalar Corp., Netherlands)测得其中田面水氮含量(CWN, mg·L-1).
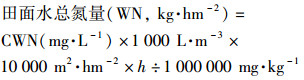
|
(8) |
土壤样品的KCl浸提液以及田面水经凯式定氮仪(Foss Scino KT260, China)消煮后, 过量滴酸至浅粉红色, 放置烘箱70℃蒸干至结晶, 收集晶体并研磨成粉末备用[31].使用同位素质谱计(MAT-251, USA, with analytic error±0.02%)分别测定粉末状液体结晶、浸提后土壤样品(1.4.3节)及植株样品(1.4.3节)的15N丰度, 记作15N在土壤矿质态氮, 土壤有机态氮, 田面水及植株(地上、地下部分)中的氮丰度(Ams、Aos、Aw、Apa、Apu, %). 15N自然丰度取0.368%, 负载用15N氯化铵丰度为10%.

|
(9) |

|
(10) |
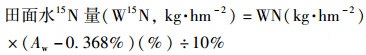
|
(11) |
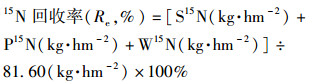
|
(12) |
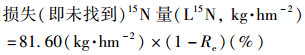
|
(13) |
本研究中土壤, 植株及田面水数据均来源于移栽60 d后的采样, 数据统计以hm2为计算单位.对分区根箱而言, 样品的相关氮量计算进行换算统一到完整根箱体系, 再按照1穴·pot-1, 200 000穴·hm-2折算到hm2.采用SPSS (SPSS ver. 16.0 for Windows, SPSS Inc., USA)中Duncan法对SN, BN及CK处理的数据进行差异显著性分析(ANOVA, P < 0.05), T检验对SN vs. BN处理的两侧样品进行比较分析(P < 0.05), Pearson法分析作物生物量及氮累积量与根系及土壤环境氮含量间的相关性; 采用Microsoft Excel软件对数据进行统计、制图.
2 结果与分析 2.1 两种根箱形式对作物地上、地下部分生物量及氮累积量的影响分区根箱是本研究实现作物对同一生境中两种氮源添加方式利用偏好研究的必要手段, 若根箱分区带来了显著的系统差异则会造成两种生境条件下结果的不可比.分别比较SN及BN在两种根箱(两种生境)条件下水稻生长情况, 结果显示(图 2):根箱分区与否(SN1 vs. SN2/3及BN1 vs. BN2/3)对水稻地上部分生物量[图 2(a)]及氮累积量[图 2(c)]无显著影响; 分区后在两侧同样处理条件下, 两侧(SN2 vs. SN3及BN2 vs. BN3)水稻地下部分生物量[图 2(b)]及氮累积量[图 2(d)]无显著差异, 且是否分区对水稻地下部分生长情况无显著影响.因此, 本研究将使用两种根箱形式的SN及BN处理分别视为同一处理, 则SN及BN处理各6个重复, SN vs. BN及CK处理各3个重复.

|
SN2/3以及BN2/3为SN及BN处理下分区根箱的地上部分 图 2 水稻在两个根箱条件下生物量及氮累积量 Fig. 2 Amount of paddy dry matter and nitrogen assimilation in partitioned and non-partitioned pot conditions |
不同生境中, 生物炭负载氮添加(BN处理)较溶液氮添加方式(SN处理)显著降低了水稻地上部分生物量及氮累积16%及14%, 而地下部分生物量却未因氮源添加方式不同而产生显著差异(图 3).而在同一生境中, 地下部分的差异显著, 生物炭负载氮添加的一侧(SN vs. BN BN), 根系生物量及氮累积分别显著高于溶液氮添加的另一侧(SN vs. BN SN)的27%及20%.

|
对CK、SN及BN处理的水稻地上及地下部分分别进行统计分析, 同一字母表示统计学差异不显著; 其中地上部分相关数据的统计结果由小写字母表示, 地下部分相关数据的统计结果由大写字母表示; 对SN vs. BN处理两侧的水稻地下部分相关数据采用T检验进行分析, 若存在显著差异则标注“*”, 下同 图 3 两种氮源添加方式下水稻生物量及氮累积量 Fig. 3 Amount of paddy dry matter and nitrogen uptake in the two nitrogen application methods |
生物炭负载氮添加在两种生境条件下均增加水稻生物量的根冠比25%~27%, 达显著水平, 且生物炭负载氮添加条件下生物量的根冠比数值与不添加外源氮处理(CK处理)相近(图 4).但两种氮源添加方式并未对水稻氮累积量的根冠比产生显著影响.

|
图 4 水稻生物量及氮累积量的根冠比 Fig. 4 Root-shoot ratio in dry matter and the amount of nitrogen uptake in the paddy |
生物炭负载氮添加方式降低了(BN处理较SN处理)水稻对外源氮36%的利用效率(图 5), 但在两种氮源共存时(SN vs. BN处理较SN处理)水稻氮利用效率并未受到显著影响.

|
图 5 氮肥表观利用效率 Fig. 5 Nitrogen recovery use efficiency |
通过根系扫描及软件分析发现, 不同生境中, 氮源添加方式的改变影响了水稻的根长、根表面积及体积; 而同一生境中, 氮源添加方式进一步影响了水稻根数(表 1).氮源添加方式对水稻根系的平均直径并无显著影响.参照顾东祥[32]等的方法(不定根D>0.3 mm、粗分枝根0.1 mm<D ≤0.3 mm、细分枝根D ≤0.1 mm), 以水稻根直径的粗细对其进行分类及分析.数据显示, 生物炭负载氮添加在不同生境中显著提高了不定根的根系长度, 在同一生境中显著提高了细分枝根、粗分枝根及不定根的根系长度.生物炭负载氮添加方式对根表面积的影响在两种生境条件下均是通过提高不定根的根表面积而实现的.不同生境中, 生物炭负载氮添加方式显著增加了细分枝根及不定根的根体积; 而同一生境中, 仅显著增加了不定根的根体积.同一生境中, 生物炭负载氮添加方式显著增加了粗细分支根的数目, 以此显著增加了整体根数.对根尖样品分析显示, 生物炭负载氮添加在两种生境条件下均显著提高了根系活力.
|
|
表 1 水稻根系形态及根系活力1) Table 1 Root morphology and root activity in the paddy |
2.4 两种氮源添加方式对土壤矿质态氮含量的影响
水稻为喜铵作物, 且耕作条件长期淹水厌氧, 土壤矿质态氮以铵态氮为主要形态.根系土中硝态氮含量显著高于非根系土壤, 而铵态氮含量在添加外源氮条件下与之相反(图 6).

|
图 6 土壤硝态氮及铵态氮含量 Fig. 6 Soil nitrate and ammonium nitrogen content |
对根系土而言, 溶液氮添加方式较生物炭负载氮添加显著提高了土壤硝态氮含量55%~58%, 但非根系土壤硝态氮含量差异不显著[图 6(a)].而生物炭负载氮添加方式较溶液氮添加显著提高土壤(根系土以及非根系土)铵态氮含量48%~53%.氮源添加方式对土壤矿质态氮含量的影响在两种生境条件下结果一致.
2.5 生物炭负载氮的回收以生物炭负载添加氮素的57%~62%在移栽60 d后仍然留在土壤中, 作物仅吸收了不足10%的量, 少于1%留存在田面水中, 其余31%~35%未在盆栽系统中寻回.本试验盆栽放置在大棚环境, 不受降雨影响且无径流产生, 因此未找到15N部分被认为以渗漏或者气态形式发生了损失.比较两种生境条件下从土壤、作物及田面水找回的15N量(表 2), 发现单侧生物炭负载氮添加方式(SN vs. BN BN)较单一生物炭负载氮添加方式(BN)有提高土壤留存15N量及降低物吸收量的趋势, 但未达到显著水平.
|
|
表 2 生物炭负载15N的去向1) Table 2 Fate of 15N adsorbed by biochar |
3 讨论
考虑氮添加量仅为81.60 kg·hm-2, 并不足以供给水稻完成其整个生育期的生长.以整个生育期200 kg·hm-2施氮量计算, 氮添加量仅为整个生长期氮投入量的40%左右.此外, 已有研究发现水稻拔节期根系形状与产量及氮利用率显著相关, 且相关性高于收获期[32, 33].为避免因氮素缺乏而带来水稻生长的误差, 更真实反映水稻正常生长时两种氮源添加方式产生的影响, 本试验将水稻的生长终止在移栽后60 d.
3.1 土壤矿质态氮含量及作物地下部分生长对作物地上部分生长的影响水稻根系是作物汲取养分的主要器官, 其形态与环境尤其是土壤环境中养分的含量息息相关[21, 22].相关性分析发现(表 3), 本研究中土壤(根际土及非根际土)矿质态氮含量与水稻地下部分生物量(0.57~0.58, P < 0.05)及氮累积量(0.62~0.63, P < 0.05)呈显著正相关, 而与地上部分生长并不显著.
|
|
表 3 作物地上地下部分生长、根系形态及土壤氮含量的相关性分析1) Table 3 Correlation analysis among the conditions of paddy growth (aboveground and underground parts), root morphology and soil mineral nitrogen content |
作物地下部分在供给地上部分养分的同时, 也因为自身生长与地上部分发生养分分配.若环境养分存在欠缺(程度较轻), 作物为更多汲取环境中的养分会增加地下与地上部分生物量的比例; 而养分充足时, 地上及地下部分会出现同步增长[20, 23].本研究中, 地上部分氮累积量与地下部分氮累积量呈显著正相关关系(0.54, P < 0.05), 说明本研究养分供应是相对充足的.
另一方面, 地下部分生物量主要由根系的体积(0.67, P < 0.01)及根长(0.57, P < 0.05)决定, 与根尖数、根表面积及平均直径无显著相关性; 而地上部分氮累积量与根尖数呈显著负相关(0.51, P < 0.05).有研究发现, 富余养分供应条件下, 地上部分生长促进的同时, 作物根系会出现根尖数量增加且根系偏细的现象[24, 25].而本研究中根尖数量与地上部分氮累积量负相关, 而根系直径与地上部分生长未发现显著相关, 说明试验条件下的养分供应未达到过剩水平.
相比于土壤矿质态氮含量正相关的水稻地下部分, 本研究中水稻地上部分的生长对土壤矿质态氮含量的变化未有显著响应, 这一现象说明, 水稻根系对土壤中氮素敏感性更高, 且本试验条件对作物的氮供应处于适当范围, 各处理均未对水稻地上部分生长产生明显影响.
3.2 生物炭负载氮添加对作物生长及氮利用的影响生物炭负载氮添加方式(两种生境条件下, BN以及SN vs. BN BN)在氮施用及水稻移栽60 d后仍然保持了土壤(根系土及非根系土)较高的铵态氮含量(图 6), 却减少了作物(图 3, 尤其是地上部分)的生物量及氮累积, 甚至在不同生境中显著降低了作物氮利用率(图 5, BN处理), 不难推测, 生物炭负载氮进入土壤环境后, 确实发生了因生物炭较强吸附作用而一定程度限制作物对氮吸收利用的现象.同一生境中, 图 6生物炭负载氮添加的一侧根长、根表面积、根体积及根尖数的显著增加(表 1), 与此前比较生物炭添加对作物根系影响的研究结果基本一致[27, 29].本研究中根系生长及形态差异可视为水稻根系为获取更多养分, 因外源氮添加方式不同而作出的响应.由此可见, 将生物炭负载氮完全替代化学肥料施用农田中, 会一定程度影响作物对氮的吸收利用.
同一生境中生物炭负载氮添加的一侧水稻地下部分生物量及氮累积量同步增加(图 2), 且两侧不同氮添加方式处理(SN vs. BN处理)的水稻地上部分氮累积量[图 1(b)]及氮利用效率(图 5)未与溶液氮添加方式(SN处理)有显著差异, 说明在生物炭负载氮及溶液氮共存的情况下, 作物氮利用并不会受到显著影响.此外, 本研究对比两种生境条件下作物对负载氮(15N)的吸收量, 也未发现显著差异(表 2).这意味着, 即使在一侧根系处于溶液氮添加条件时(SN vs. BN SN), 作物也并未减弱对另一侧(SN vs. BN SN)生物炭负载氮的吸收利用, 生物炭负载的氮生物有效性并不低; 同时, 超过一半原本生物炭负载的氮发现留存在土壤中可成为下一季作物生长的养分来源, 确实达到了养分迁移存储的目的.
因此, 利用生物炭作为氮素载体、构建富营养水体及农田养分回用枢纽具可行性, 但生物炭负载氮素不可全量替代化学肥料作为外源氮投入, 寻找适宜添加比例可以在实现养分回用同时减少氮素流失风险, 双向削减农田系统的面源污染.
4 结论(1) 通过在两种生境条件下比较两种氮源添加方式对水稻生长的影响, 发现溶液氮添加方式更有利于水稻地上部分生物量及氮累积; 生物炭负载氮的添加方式显著增加了水稻根冠生物量比值.
(2) 将生物炭负载氮完全替代化学肥料施用农田中, 会因生物炭对氮的吸附作用而影响水稻生物量及氮累积, 但是在两种添加方式共存的情况下, 水稻并不会对某种氮呈现吸收利用偏好, 且氮利用率未受到显著影响.
(3) 综上所述, 生物炭可以作为载体实现水体到农田的氮素迁移, 以适宜比例与普通化学肥料共同施用并不会对作物氮素利用产生不良影响并可增加土壤矿质态氮固持量.
| [1] | Le C, Zha Y, Li Y, et al. Eutrophication of lake waters in China:cost, causes, and control[J]. Environmental Management, 2010, 45(4): 662-668. DOI:10.1007/s00267-010-9440-3 |
| [2] |
朱兆良, Norse D, 孙波. 中国农业面源污染控制对策[M]. 北京: 中国环境科学出版社, 2006. Zhu Z L, Norse D, Sun B. Policy for reducing non-point pollution from crop production in China[M]. Beijing: China Environmental Science Press, 2006. |
| [3] | 中华人民共和国环境保护部, 中华人民共和国国家统计局, 中华人民共和国农业部.第一次全国污染源普查公报[EB/OL]. http://www.stats.gov.cn/tjsj/tjgb/qttjgb/qgqttjgb/201002/t20100211_30641.html, 2010-02-11. |
| [4] | Chen J. Rapid urbanization in China:a real challenge to soil protection and food security[J]. Catena, 2007, 69(1): 1-15. DOI:10.1016/j.catena.2006.04.019 |
| [5] |
张红举, 陈方. 太湖流域面源污染现状及控制途径[J]. 水资源保护, 2010, 26(3): 87-90. Zhang H J, Chen F. Non-point pollution statistics and control measures in Taihu Basin[J]. Water Resources Protection, 2010, 26(3): 87-90. DOI:10.3969/j.issn.1004-6933.2010.03.022 |
| [6] |
杨林章, 王德建, 夏立忠. 太湖地区农业面源污染特征及控制途径[J]. 中国水利, 2004(20): 29-30. Yang L Z, Wang D J, Xia L Z. Features and ways of control of non-point agricultural pollution in Taihu Area[J]. China Water Resources, 2004(20): 29-30. DOI:10.3969/j.issn.1000-1123.2004.20.006 |
| [7] |
杨林章, 施卫明, 薛利红, 等. 农村面源污染治理的"4R"理论与工程实践——总体思路与"4R"治理技术[J]. 农业环境科学学报, 2013, 32(1): 1-8. Yang L Z, Shi W M, Xue L H, et al. Reduce-Retain-Reuse-Restore technology for the controlling the agricultural non-point source pollution in countryside in China:general countermeasures and technologies[J]. Journal of Agro-Environment Science, 2013, 32(1): 1-8. |
| [8] |
薛利红, 何世颖, 段婧婧, 等. 基于养分回用-化肥替代的农业面源污染氮负荷削减策略及技术[J]. 农业环境科学学报, 2017, 36(7): 1226-1231. Xue L H, He S Y, Duan J J, et al. Agricultural non-point source pollution nitrogen load reduction strategy and technology of nutrient reusing in agricultural fields to replace fertilizer[J]. Journal of Agro-Environment Science, 2017, 36(7): 1226-1231. |
| [9] |
赵冬, 颜廷梅, 乔俊, 等. 太湖地区稻田氮素损失特征及环境效应分析[J]. 生态环境学报, 2012, 21(6): 1149-1154. Zhao D, Yan T M, Qiao J, et al. Characteristics of N loss and environmental effect of paddy field in Taihu area[J]. Ecology and Environmental Sciences, 2012, 21(6): 1149-1154. |
| [10] | Wang X Z, Zhu J G, Gao R, et al. Nitrogen cycling and losses under rice-wheat rotations with coated urea and urea in the Taihu Lake region[J]. Pedosphere, 2007, 17(1): 62-69. DOI:10.1016/S1002-0160(07)60008-3 |
| [11] |
薛利红, 俞映倞, 杨林章. 太湖流域稻田不同氮肥管理模式下的氮素平衡特征及环境效应评价[J]. 环境科学, 2011, 32(4): 1133-1138. Xue L H, Yu Y L, Yang L Z. Nitrogen balance and environmental impact of paddy field under different N management methods in Taihu Lake region[J]. Environmental Science, 2011, 32(4): 1133-1138. |
| [12] | Atkinson C J, Fitzgerald J D, Hipps N A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils:a review[J]. Plant and Soil, 2010, 337(1-2): 1-18. DOI:10.1007/s11104-010-0464-5 |
| [13] | Lehmann J, Joseph S. Biochar for environmental management:science and technology[M]. London: Science and Technology, 2009. |
| [14] | Yao Y, Gao B, Zhang M, et al. Effect of biochar amendment on sorption and leaching of nitrate, ammonium, and phosphate in a sandy soil[J]. Chemosphere, 2012, 89(11): 1467-1471. DOI:10.1016/j.chemosphere.2012.06.002 |
| [15] | Ding Y, Liu Y X, Wu W X, et al. Evaluation of biochar effects on nitrogen retention and leaching in multi-layered soil columns[J]. Water, Air, & Soil Pollution, 2010, 213(1-4): 47-55. |
| [16] | Hale S E, Alling V, Martinsen V, et al. The sorption and desorption of phosphate-P, ammonium-N and nitrate-N in cacao shell and corn cob biochars[J]. Chemosphere, 2013, 91(11): 1612-1619. DOI:10.1016/j.chemosphere.2012.12.057 |
| [17] | Taghizadeh-Toosi A, Clough T J, Sherlock R R, et al. Biochar adsorbed ammonia is bioavailable[J]. Plant and Soil, 2012, 350(1-2): 57-69. DOI:10.1007/s11104-011-0870-3 |
| [18] |
蔡昆争, 骆世明, 段舜山. 水稻群体根系特征与地上部生长发育和产量的关系[J]. 华南农业大学学报, 2005, 26(2): 1-4. Cai K Z, Luo S M, Duan S S. The relationship between root system of rice and aboveground characteristics and yield[J]. Journal of South China Agricultural University, 2005, 26(2): 1-4. |
| [19] |
魏海燕, 张洪程, 张胜飞, 等. 不同氮利用效率水稻基因型的根系形态与生理指标的研究[J]. 作物学报, 2008, 34(3): 429-436. Wei H Y, Zhang H C, Zhang S F, et al. Root morphological and physiological characteristics in rice genotypes with different n use efficiencies[J]. Acta Agronomica Sinica, 2008, 34(3): 429-436. DOI:10.3321/j.issn:0496-3490.2008.03.013 |
| [20] |
凌启鸿, 张国平, 朱庆森, 等. 水稻根系对水分和养分的反应[J]. 江苏农学院学报, 1990, 11(1): 23-28. Ling Q H, Zhang G P, Zhu Q X, et al. Responses of root system of rice to water and nutrients[J]. Journal of Jiangsu Agricultural college, 1990, 11(1): 23-28. |
| [21] | Marschner H, Kirkby E A, Cakmak I. Effect of mineral nutritional status on shoot-root partitioning of photoassimilates and cycling of mineral nutrients[J]. Journal of Experimental Botany, 1996, 47(l): 1255-1263. |
| [22] | Wang Y, Mi G H, Chen F J, et al. Response of Root Morphology to Nitrate supply and its contribution to nitrogen accumulation in Maize[J]. Journal of Plant Nutrition, 2005, 27(12): 2189-2202. DOI:10.1081/PLN-200034683 |
| [23] |
樊剑波, 张亚丽, 万小羽, 等. 水稻根系与氮素吸收利用之研究进展[J]. 中国农学通报, 2007, 23(2): 236-240. Fan J B, Zhang Y L, Wan X Y, et al. Progress in research of rice root related to nitrogen uptake and utilization[J]. Chinese Agricultural Science Bulletin, 2007, 23(2): 236-240. DOI:10.3969/j.issn.1000-6850.2007.02.056 |
| [24] |
张亚洁, 华晶晶, 黄银琪, 等. 旱、水稻根系生长对水氮响应的研究[J]. 江苏农业科学, 2017, 45(11): 55-59. Zhang Y J, Hua J J, Huang Y Q, et al. Study on response of root growth of upland rice and paddy rice to water and nitrogen[J]. Jiangsu Agricultural Sciences, 2017, 45(11): 55-59. |
| [25] |
孙虎威, 王文亮, 刘尚俊, 等. 低氮胁迫下水稻根系的发生及生长素的响应[J]. 土壤学报, 2014, 51(5): 1096-1102. Sun H W, Wang W L, Liu S J, et al. Formation of rice root regulated by nitrogen deficiency[J]. Acta Pedologica Sinica, 2014, 51(5): 1096-1102. |
| [26] | Prendergast M T, Duvall M, Sohi S P. Biochar-root interactions are mediated by biochar nutrient content and impacts on soil nutrient availability[J]. European Journal of Soil Science, 2014, 65(1): 173-185. DOI:10.1111/ejss.12079 |
| [27] |
张伟明, 孟军, 王嘉宇, 等. 生物炭对水稻根系形态与生理特性及产量的影响[J]. 作物学报, 2013, 39(8): 1445-1451. Zhang W M, Meng J, Wang J Y, et al. Effect of biochar on root morphological and physiological characteristics and yield in rice[J]. Acta Agronomica Sinica, 2013, 39(8): 1445-1451. |
| [28] |
李中阳, 齐学斌, 樊向阳, 等. 生物质炭对冬小麦产量、水分利用效率及根系形态的影响[J]. 农业工程学报, 2015, 31(12): 119-124. Li Z Y, Qi X B, Fan X Y, et al. Influences of biochars on growth, yield, water use efficiency and root morphology of winter wheat[J]. Transactions of the Chinese Society of Agricultural Engineering, 2015, 31(12): 119-124. DOI:10.11975/j.issn.1002-6819.2015.12.016 |
| [29] | Yang L, Liao F, Huang M, et al. Biochar improves sugarcane seedling root and soil properties under a pot experiment[J]. Sugar Tech, 2015, 17(1): 36-40. DOI:10.1007/s12355-014-0335-0 |
| [30] | Wang H F, Zheng H, Jiang Z X, et al. Efficacies of biochar and biochar-based amendment on vegetable yield and nitrogen utilization in four consecutive planting seasons[J]. Science of the Total Environment, 2017, 593-594: 124-133. DOI:10.1016/j.scitotenv.2017.03.096 |
| [31] | Zhang J B, Cai Z C, Cheng Y, et al. Denitrification and total nitrogen gas production from forest soils of Eastern China[J]. Soil Biology and Biochemistry, 2009, 41(12): 2551-2557. DOI:10.1016/j.soilbio.2009.09.016 |
| [32] |
顾东祥, 汤亮, 徐其军, 等. 水氮处理下不同品种水稻根系生长分布特征[J]. 植物生态学报, 2011, 35(5): 558-566. Gu D X, Tang L, Xu Q J, et al. Root growth and distribution in rice cultivars as affected by nitrogen and water supply[J]. Chinese Journal of Plant Ecology, 2011, 35(5): 558-566. |
| [33] |
李娜, 杨志远, 代邹, 等. 水氮管理对不同氮效率水稻根系性状、氮素吸收利用及产量的影响[J]. 中国水稻科学, 2017, 31(5): 500-512. Li N, Yang Z Y, Dai Z, et al. Effects of water-nitrogen management on root traits, nitrogen accumulation and utilization and grain yield in rice with different nitrogen use efficiency[J]. Chinese Journal of Rice Science, 2017, 31(5): 500-512. |