 2018, Vol. 39
2018, Vol. 39



好氧反硝化细菌因其具有适应性较强、生长速度快、容易控制等优点, 而受到国内外研究者的青睐[1].目前, 已有大量好氧反硝化细菌的研究报道, 其中以假单胞菌属最多[2], 主要有斯氏假单胞菌(Pseudomonas stutzeri)[3]、铜绿假单胞菌(Pseudomonas aeruginosa)[4]、托拉斯假单胞菌(Pseudomonas tolaasii)[5]和恶臭假单胞菌(Pseudomonas putida)[6]等.而国内外关于好氧反硝化蒙氏假单胞菌(Pseudomonas monteilii)的研究报道很少, 仅有张文艺等[7]和冯雅丽等[8]的研究报道.而张文艺等未报道该蒙氏假单胞菌N1的反硝化特性; 冯雅丽等发现的蒙氏假单胞菌YL-1在pH大于9.0, 温度低于25℃或高于35℃时, 基本丧失对硝酸盐氮的反硝化能力.
事实上, 脱氮生物菌易受外在环境影响, 环境中的温度和酸碱性就是影响生物脱氮的重要因素.目前报道的脱氮微生物大多数是在20~35℃、中性或微碱性条件下进行, 对低温、过酸或过碱的环境条件耐受性较弱.如菌株Pseudomonas stutzeri F1[9]在温度低于20℃或pH值大于8.0的条件下, 其生长与反硝化脱氮作用均受到抑制; 菌株Pseudomonas sp.2-8[10]在温度低于20℃或pH值大于9.0的条件下, 基本丧失反硝化能力.而现实中碱性氮污染废水的来源广泛, 如生产化肥的废水pH值达8.0~10.0[11]; 生产石油树脂的废水pH值可达11.0~13.0[12].因此, 研究耐冷嗜碱高效生物脱氮菌对处理南方地区冬季碱性废水中的氮污染具有重要意义.
本实验从贵州冬水田中分离获得1株具有好氧反硝化作用的菌株H97, 结合形态学、磷脂脂肪酸及16S rRNA基因序列分析, 该菌被鉴定为蒙氏假单胞菌(Pseudomonas monteilii), 通过环境因子对菌株生物脱氮的影响发现, 该菌在15~40℃和pH 7.0~12.0范围内均能进行高效的脱氮作用, 研究结果为南方地区冬季碱性废水中氮污染生物处理提供了一种选择, 以期为此后该菌的具体应用奠定基础.
1 材料与方法 1.1 材料 1.1.1 菌株来源新分离菌株H97为本实验室从贵州省冬水田中分离筛选获得.
1.1.2 培养基(1) BTB培养基(bromothymol blue, BTB)[13]: NaNO2 1.0 g ·L-1, KH2PO4 1.0 g ·L-1, FeCl2 ·6H2O 0.5 g ·L-1, CaCl2 ·7H2O 0.2 g ·L-1, MgSO4 ·7H2O 1.0 g ·L-1, 琥珀酸钠8.5 g ·L-1(固体加2.0%的琼脂), BTB(1.5%溴百里酚蓝溶于无水乙醇)1mL, 调节pH至7.2.
(2) 反硝化培养基(DM)[14]:乙酸钠2.56 g ·L-1, NaNO3 0.31 g ·L-1, KH2PO4 1.5 g ·L-1, Na2HPO4 0.42 g ·L-1, MgSO4 ·7H2O 1.0 g ·L-1, FeSO4 ·7H2O 0.05 g ·L-1, 调节pH至7.2.
(3) LB培养基:胰蛋白胨10.0 g ·L-1, 酵母粉5.0 g ·L-1, NaCl 10.0 g ·L-1, 固体加2.0%的琼脂, 调节pH至7.2.
将上述所有培养基在0.11 MPa、121℃条件下灭菌30min, 冷却后备用.
1.2 细菌鉴定 1.2.1 形态学特征观察将分离纯化的菌株H97于BTB固体培养基上划线, 在15℃条件下培养, 待长出菌落后观察其菌落形态, 采用AFM原子力显微镜(DIMENSION ICON WITH SCANASYST, 美国BRUKER公司)扫描观察菌体形态.
1.2.2 特异性磷脂脂肪酸鉴定取纯化菌株H97进行皂化、甲基化、萃取以及碱洗涤后获得上机样品, 用Agilent 6850气相色谱仪(FID检测器)分析特异性磷脂脂肪酸(Phospholipid fatty acid, PLFA)的成分.各脂肪酸成分通过MIDI Sherlock微生物鉴定系统(Version6.1, MIDI, Inc., Newark, DE)进行检测.
1.2.3 菌株H97的16S rRNA基因序列检测以纯化菌株H97的基因组DNA为模板, 采用通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′)进行16S rRNA聚合酶链式反应(PCR)扩增, 将克隆产物送测序公司测序, 将16S rRNA基因序列在GenBank核酸序列数据库中进行同源性比较, 以Neighbor-Joining法[15]构建系统发育树.
1.3 影响菌株H97好氧反硝化特性的因素分析向装有100 mL反硝化培养基的250 mL三角瓶中接入菌株H97, 研究不同温度(5、10、15、20、25、30、35和40℃)、接菌量[0.5×106、1.0×106、1.5×106、2.0×106和2.5×106 CFU ·(100 mL)-1]、C/N值(0、5、10、15、20和25)、初始pH值(6.0、7.0、8.0、9.0、10.0、11.0、12.0和13.0)和碳源种类(柠檬酸钠、丁二酸钠、葡萄糖、蔗糖和乙酸钠)对菌株H97反硝化特性的影响.以上处理均在150 r ·min-1摇床条件下培养48 h后, 测定菌株H97的生长、硝酸盐氮和总氮的浓度, 并计算硝酸盐氮和总氮的去除率.
1.4 分析方法本研究中, 硝酸盐氮的测定采用紫外分光光度法[16], 总氮的测定采用碱性过硫酸钾消解紫外分光光度法[17], 菌株的生长采用光度比浊法(D600).脱氮率计算方法如下:
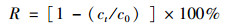
|
(1) |
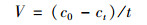
|
(2) |
式中, R为氮的去除率, ct为终态氮浓度(mg ·L-1), c0为初始氮浓度(mg ·L-1), V为氮的去除速率[mg ·(L ·h)-1], t为时间.
采用Excel、SPSS Statistics 19.0、MEGA 6.0和Origin 8.6软件对实验数据进行统计分析与作图.
2 结果与分析 2.1 细菌鉴定 2.1.1 形态学特征在BTB固体培养基上观察到菌株H97的菌落形态特征为:圆形, 呈黑色, 表面湿润光滑, 边缘整齐, 中央略凸起, 革兰氏染色为阴性.经原子力显微镜扫描观察, 结果如图 1所示, 菌体呈短杆状或椭球状, 不产芽孢, 无鞭毛.

|
图 1 原子力显微镜扫描下的菌株H97 Fig. 1 Strain H97 under the scanning atomic force microscope |
磷脂脂肪酸(PLFAs)是微生物细胞膜的主要成分, 不同类群微生物细胞膜的PLFAs存在差异, 同种类群微生物PLFAs含量比较恒定[18], 它的种类和含量是细胞分类法的重要依据之一.气相色谱能快速、灵敏、精确地检测出细菌细胞内的脂肪酸[19].
本研究中, 菌株H97的磷脂脂肪酸鉴定结果如表 1所示, 该菌株与Pseudomonas syringae tomato的相似性为0.569.
|
|
表 1 特异性磷脂脂肪酸的鉴定结果 Table 1 Result of the specific phospholipid fatty acid identification |
2.1.3 16S rRNA基因序列分析
菌株H97的16S rRNA基因全长1 392 bp, 在GenBank中的登录号为KY927410, 系统发育树如图 2所示, 该菌与Pseudomonas monteilii的同源性达99%.基于磷脂脂肪酸鉴定的结果, 在传统分类学上, Pseudomonas monteilii属于丁香假单胞菌(Pseudomonas syringae)[20], 这与该菌16S rRNA基因序列分析的结果一致.因此, 结合形态学、磷脂脂肪酸和16S rRNA基因序列分析的结果, 可以将菌株H97确定为蒙氏假单胞菌(Pseudomonas monteilii).

|
图 2 菌株H97的系统发育进化树 Fig. 2 Phylogenetic tree of strain H97 |
如图 3所示, 当温度为5℃和10℃时, 菌株H97几乎不能生长; 但当温度上升到15℃时, 48 h内, 硝酸盐氮浓度由52.73 mg ·L-1下降至11.02 mg ·L-1, 平均反硝化速率为0.87 mg ·(L ·h)-1, 总氮的平均去除速率为0.81 mg ·(L ·h)-1, 菌株H97对硝酸盐氮和总氮的去除率分别达80.55%和73.18%;当温度为15~30℃范围内时, 菌株H97的生长和脱氮效率基本保持不变; 当温度继续上升至40℃时, 菌株H97的生长和脱氮率逐渐降低, 对硝酸盐氮和总氮的去除率分别为63.60%和48.65%.

|
图 3 温度对菌株H97反硝化特性的影响 Fig. 3 Effects of temperature on the denitrificationability of strain H97 |
如图 4所示, 当接菌量为0.5×106 CFU时(每100 mL培养基), 硝酸盐氮和总氮的去除率仅为18.01%和17.23%;但当接菌量上升至1.5×106 CFU ·(100 mL)-1时, 48 h内, 硝酸盐氮浓度由48.46mg ·(L ·h)-1下降至5.76mg ·(L ·h)-1, 平均反硝化速率为0.89 mg ·(L ·h)-1, 总氮的平均去除速率为0.68 mg ·(L ·h)-1, 菌株H97对硝酸盐氮和总氮去除率分别达88.12%和59.07%;将接菌量继续增大到2.5×106 CFU ·(100 mL)-1时, 总氮的去除率下降到57.82%.因此, 在实际应用中可以将菌株H97的接菌量控制在1.5~2.0×106 CFU ·(100 mL)-1.

|
图 4 接菌量对菌株H97反硝化特性的的影响 Fig. 4 Effects of inoculation quantity on thedenitrification ability of strain H97 |
如图 5所示, 当C/N小于10时, 菌株H97的生长和反硝化活性受到一定程度的抑制, 对硝酸盐氮的去除不完全, 几乎不能去除总氮; 当C/N上升至15时, 硝酸盐氮浓度由50.93 mg ·L-1下降至0.44 mg ·L-1, 平均反硝化速率为1.05 mg ·(L ·h)-1, 总氮的平均去除速率为0.7 mg ·(L ·h)-1, 该菌对硝酸盐氮和总氮的去除率分别达99.13%和61.42%;当C/N大于15时, 脱氮率呈下降趋势.综合考虑, 菌株H97的最佳C/N为15.

|
图 5 C/N对菌株H97反硝化能力的影响 Fig. 5 Effects of C/N on the denitrification ability of strain H97 |
如图 6所示, 当pH为6.0时, 不利于菌株H97的生长; 当pH 7.0~11.0时, 菌株H97能很好生长, 对硝酸盐氮和总氮的去除率较高, 尤其pH=9.0时, 硝酸盐氮浓度由50.92 mg ·L-1下降到0.47 mg ·L-1, 平均反硝化速率为1.05 mg ·(L ·h)-1, 总氮由53.44 mg ·L-1下降至4.39 mg ·L-1, 平均去除速率1.02 mg ·(L ·h)-1, 该菌对硝态氮和总氮的去除率分别达99.07%和91.78%; pH=11.0时, 菌株H97生长较好, 脱氮效果较强; pH=12.0时, 其生长变弱, 仍具有一定脱氮能力, 对硝酸盐氮和总氮的去除率分别为64.75%和36.78%.

|
图 6 pH对菌株H97反硝化能力的影响 Fig. 6 Effects of pH on the denitrification ability of strain H97 |
如图 7所示, 碳源种类对菌株H97的生长及脱氮率的影响存在很大差异.当以柠檬酸钠、丁二酸钠和乙酸钠为碳源时, 菌株H97均能很好地生长, 尤其以丁二酸钠为碳源时, 48 h内, 硝酸盐氮浓度由50.40 mg ·L-1下降至1.17 mg ·L-1, 平均反硝化速率达1.03 mg ·(L ·h)-1, 总氮由55.53 mg ·L-1下降至2.05 mg ·L-1, 平均去除速率为1.11 mg ·(L ·h)-1, 其对硝酸盐氮和总氮的去除率分别达97.69%和96.32%;当以葡萄糖为碳源时, 其对总氮的去除率仅为46.42%;而以蔗糖为碳源时, 该菌几乎不能生长.因此, 综合考虑菌株H97的生长和脱氮效果, 以及丁二酸钠价格昂贵, 在实际污水处理中可以将柠檬酸钠或乙酸钠作为菌株H97的最佳碳源.

|
图 7 碳源种类对菌株H97反硝化能力的影响 Fig. 7 Effects of carbon source on the denitrificationability of strain H97 |
温度是影响微生物反硝化特性的重要因素之一, 主要是通过影响微生物细胞结构来影响其生长和新陈代谢[21].一般地, 好氧反硝化细菌的最适温度为20~35℃, 如Ye等[22]报道了菌株Providencia rettgeri strain YL的最适脱氮温度为25℃; 李卫芬等的研究表明[23], 菌株 Pseudomonas stutzeri F1在温度高于35℃时, 其生长和脱氮能力均受到抑制.上述结论与本研究结果不同, 菌株H97能够适应15~40℃的环境温度, 优于目前分离出的大多数好氧反硝化细菌; 该菌在15℃时生长最好, 脱氮率最高.这可能是因为该菌分离自贵州长期淹水的冬水田泥土中, 在15℃实验条件下分离筛选获得, 对低温环境的适应能力较强, 在低温环境下能较好地进行脱氮作用.因此, 该菌为处理南方地区冬季氮污染水体提供了选择.
接菌量是影响细菌反硝化特性的重要因素之一.在反硝化过程中, 若接菌量不足则不能充分发挥微生物生长繁殖的潜力, 导致脱氮效果差; 而接菌量过多会导致细菌对营养物质的竞争, 部分细菌死亡, 脱氮率降低. He等的研究发现[24], 当接菌量为1.5×106 CFU时, 好氧反硝化细菌Arthrobacter arilaitensis strain Y-10对总氮的去除率最高(52.5%), 当接菌量上升至2.5×106 CFU时, 总氮的去除率下降至32.27%, 与本研究结果相似.
C/N是菌株能否进行完全反硝化的关键因素, 碳氮源的比例直接影响细菌的生长和对有机物质的去除效果.若初始碳源充足, 则能为细菌脱氮提供足够的能量; 若碳源不足, 则不能提供足够的能量供细菌生长, 导致反硝化效率降低[25].但当碳源高于细菌的需求时, 其生长和脱氮率会保持在一个稳定水平, 若继续增加碳源, 脱氮效率不会有太大变化, 甚至有下降趋势[26]. Duan等的研究发现[27], 菌株Vibrio diabolicus SF16对氮的去除率首先随C/N值的增大而升高, 当C/N达10时去除率最高(93.00%±2.31%), 之后随C/N值的增大而降低.菌株H97的最佳C/N为15, 但当C/N为10时, 该菌依然可以表现出良好的脱氮效果, 因此, 在实际应用过程中, 可通过延长生物脱氮时间, 而达到良好的脱氮效果.
一般来说, 过酸或过碱都会影响细菌的酶活性, 导致反硝化能力降低.目前分离出的大多数好氧反硝化细菌最适pH为中性或偏碱性, 在pH=11.0时几乎不能生长, 不表现反硝化活性或反硝化活性很低. Wang等[28]对好氧反硝化假单胞菌HS-N62的研究表明, 当pH达10.0时, 其对硝酸盐氮的去除率不足50%, 与本研究结果不同, 菌株H97能在pH=11.0的碱性条件下高效脱氮, pH=12.0时, 仍能生存和脱氮, 具嗜碱特性, 表明菌株H97对处理碱性环境中的氮污染水体具有应用价值.
在反硝化过程中, 碳源作为电子供体和供微生物生长繁殖所需的营养物质, 最终将硝酸盐氮还原成含氮气体, 达到除氮效果[29].不同碳源在微生物代谢过程中的地位不同, 菌株对其的利用率也不同[30].在本研究中, 菌株H97对柠檬酸钠、丁二酸钠和乙酸钠的利用率较高, 对葡萄糖的利用率较低, 几乎不能利用蔗糖.可能是由于结构越简单, 分子量越小的碳源越容易被微生物降解, 而且柠檬酸钠、丁二酸钠和乙酸钠是三羧酸循环的中间产物, 更容易被菌株H97利用[31].
4 结论(1) 本实验从贵州冬水田中分离获得1株具有好氧反硝化作用的菌株H97, 结合形态学、磷脂脂肪酸及16S rRNA基因序列分析, 该菌被鉴定为蒙氏假单胞菌(Pseudomonas monteilii).
(2) 菌株H97具有较强好氧反硝化能力, 在初始硝酸盐氮为50 mg ·L-1时, 其最佳脱氮条件为:温度15℃, 接菌量为1.5×106 CFU ·(100 mL)-1, pH 9.0, C/N=15, 碳源丁二酸钠.
(3) 菌株H97能在15~40℃范围内生存和脱氮, 具有一定的耐冷特性, 对环境温度适应范围较广; 在pH=12.0时, 仍能生长和脱氮, 具嗜碱特性, 表明菌株H97是1株耐冷嗜碱好氧反硝化细菌.
| [1] | Jin R F, Liu T Q, Liu G F, et al. Simultaneous heterotrophic nitrification and aerobic denitrification by the marine origin bacterium Pseudomonas sp. ADN-42[J]. Applied Biochemistry and Biotechnology, 2015, 175(4): 2000-2011. DOI:10.1007/s12010-014-1406-0 |
| [2] | Ji B, Yang K, Wang H Y, et al. Aerobic denitrification by Pseudomonas stutzeri C3 incapable of heterotrophic nitrification[J]. Bioprocess and Biosystems Engineering, 2015, 38(2): 407-409. DOI:10.1007/s00449-014-1271-9 |
| [3] | Sun Y L, Feng L, Li A, et al. Ammonium assimilation:an important accessory during aerobic denitrification of Pseudomonas stutzeri T13[J]. Bioresource Technology, 2017, 234: 264-272. DOI:10.1016/j.biortech.2017.03.053 |
| [4] |
梁贤, 任勇翔, 杨垒, 等. 异养硝化-好氧反硝化菌YL的脱氮特性[J]. 环境科学, 2015, 36(5): 1749-1756. Liang X, Ren Y X, Yang L, et al. Characteristics of nitrogen removal by a heterotrophic nitrification-aerobic denitrification bacterium YL[J]. Environmental Science, 2015, 36(5): 1749-1756. |
| [5] |
何腾霞, 徐义, 李振轮. 耐冷亚硝酸盐型反硝化菌Pseudomonas tolaasii Y-11的鉴定及其脱氮特性[J]. 微生物学报, 2015, 55(8): 991-1000. He T X, Xu Y, Li Z L. Identification and characterization of a hypothermia nitrite bacterium Pseudomonas tolaasii Y-11[J]. Acta Microbiologica Sinica, 2015, 55(8): 991-1000. |
| [6] | Xu Y, He T X, Li Z L, et al. Nitrogen removal characteristics of Pseudomonas putida Y-9 capable of heterotrophic nitrification and aerobic denitrification at low temperature[J]. BioMed Research International, 2017, 2017: 1429018. |
| [7] |
张文艺, 陈雪珍, 陆丽巧, 等. BAF反应器中好氧反硝化菌的分子鉴定及分析[J]. 土木建筑与环境工程, 2012, 34(1): 118-123. Zhang W Y, Chen X Z, Lu L Q, et al. Molecular classification, identification and phylogenetic analysis of aerobic denitrifier N1 in the BAF reator[J]. Journal of Civil, Architectural & Environmental Engineering, 2012, 34(1): 118-123. |
| [8] |
冯雅丽, 李洪珊, 李浩然. 一株深海反硝化菌的分离鉴定及反硝化特性研究[J]. 环境污染与防治, 2016, 38(12): 27-30. Feng Y L, Li H S, Li H R. Identification and denitrification characteristics of a deep sea denitrifier[J]. Environmental Pollution & Control, 2016, 38(12): 27-30. |
| [9] | Zhang H N, Wang H Y, Yang K, et al. Nitrate removal by a novel autotrophic denitrifier (Microbacterium sp.) using Fe(Ⅱ) as electron donor[J]. Annals of Microbiology, 2015, 65(2): 1069-1078. DOI:10.1007/s13213-014-0952-6 |
| [10] |
高喜燕, 刘鹰, 郑海燕, 等. 一株海洋好氧反硝化细菌的鉴定及其好氧反硝化特性[J]. 微生物学报, 2010, 50(9): 1164-1171. Gao X Y, Liu Y, Zheng H Y, et al. Identification and characteristics of a marine aerobic denitrifying bacterium[J]. Acta Microbiologica Sinica, 2010, 50(9): 1164-1171. |
| [11] |
孟范平, 姚瑞华, 刘强, 等. 利用纳米材料复合膜处理高浓度氯化铵废水[J]. 水处理技术, 2006, 32(9): 52-55. Meng F P, Yao R H, Liu Q, et al. Treatment of wastewater containing high strong ammonium chloride by nano membrane device[J]. Technology of Water Treatment, 2006, 32(9): 52-55. |
| [12] | 李积云. 浅析碱性废水处理方法[J]. 中国化工贸易, 2012(11): 147. |
| [13] | Guo Y, Zhou X M, Li Y G, et al. Heterotrophic nitrification and aerobic denitrification by a novel Halomonas campisalis[J]. Biotechnology Letters, 2013, 35(12): 2045-2049. DOI:10.1007/s10529-013-1294-3 |
| [14] | Song Z F, An J, Fu G H, et al. Isolation and characterization of an aerobic denitrifying Bacillus sp. YX-6 from shrimp culture ponds[J]. Aquaculture, 2011, 319(1-2): 188-193. DOI:10.1016/j.aquaculture.2011.06.018 |
| [15] |
王兆阳, 陈国耀, 姜珂, 等. 1株耐冷兼性嗜碱好氧反硝化菌的分离鉴定及反硝化特性[J]. 环境科学, 2014, 35(6): 2341-2348. Wang Z Y, Chen G Y, Jiang K, et al. Identification and denitrification characteristics of a psychrotolerant facultative basophilic aerobic denitrifier[J]. Environmental Science, 2014, 35(6): 2341-2348. |
| [16] | 国家环境保护总局. 水和废水监测分析方法[M]. 第四版. 北京: 中国环境科学出版社, 2002. |
| [17] | HJ 636-2012, 水质总氮的测定碱性过硫酸钾消解紫外分光光度法[S]. |
| [18] |
李南洁, 曾清苹, 何丙辉, 等. 缙云山柑橘林土壤微生物磷脂脂肪酸(PLFAs)及酶活性的季节变化特征[J]. 环境科学, 2017, 38(1): 309-317. Li N J, Zeng Q P, He B H, et al. Seasonal variations of soil microbial PLFAs and soil enzyme activity under the citrus plantation in Mt. Jinyun, Chongqing[J]. Environmental Science, 2017, 38(1): 309-317. |
| [19] |
李正义, 贾俊涛, 曹际娟, 等. 一株分离自美国龙虾的麦氏弧菌的鉴定及其低温状态下细胞脂肪酸的变化[J]. 微生物学报, 2013, 53(6): 628-634. Li Z Y, Jia J T, Cao J J, et al. Identification of Vibrio metschnikovii from Homarus americanus and its changes in membrane fatty acid composition in response to low temperature[J]. Acta Microbiologica Sinica, 2013, 53(6): 628-634. |
| [20] | Sneath P H A, Mair N S, Sharpe E. Bergey's manual of systematic bacteriology[M]. Volume 2. Baltimore, USA: Williams & Wilkins, 1986. |
| [21] | Qin W, Li W G, Zhang D Y, et al. Ammonium removal of drinking water at low temperature by activated carbon filter biologically enhanced with heterotrophic nitrifying bacteria[J]. Environmental Science and Pollution Research, 2016, 23(5): 4650-4659. DOI:10.1007/s11356-015-5561-9 |
| [22] | Ye J, Zhao B, An Q, et al. Nitrogen removal by Providencia rettgeri strain YL with heterotrophic nitrification and aerobic denitrification[J]. Environmental Technology, 2016, 37(17): 2206-2213. DOI:10.1080/09593330.2016.1146338 |
| [23] |
李卫芬, 傅罗琴, 邓斌, 等. 1株好氧反硝化菌的分离鉴定及反硝化特性研究[J]. 环境科学, 2011, 32(8): 2403-2408. Li W F, Fu L Q, Deng B, et al. Identification and denitrification characteristics of an aerobic denitrifier[J]. Environmental Science, 2011, 32(8): 2403-2408. |
| [24] | He T X, Li Z L. Identification and denitrification characterization of a novel hypothermia and aerobic nitrite-denitrifying bacterium, Arthrobacter arilaitensis strain Y-10[J]. Desalination and Water Treatment, 2016, 57(41): 19181-19189. DOI:10.1080/19443994.2015.1099471 |
| [25] | Kim J K, Park K J, Cho K S, et al. Aerobic nitrification-denitrification by heterotrophic Bacillus strains[J]. Bioresource Technology, 2005, 96(17): 1897-1906. DOI:10.1016/j.biortech.2005.01.040 |
| [26] |
孙庆花, 于德爽, 张培玉, 等. 1株海洋异养硝化-好氧反硝化菌的分离鉴定及其脱氮特性[J]. 环境科学, 2016, 37(2): 647-654. Sun Q H, Yu D S, Zhang P Y, et al. Identification and nitrogen removal characteristics of a heterotrophic nitrification-aerobic denitrification strain isolated from marine environment[J]. Environmental Science, 2016, 37(2): 647-654. |
| [27] | Duan J M, Fang H D, Su B, et al. Characterization of a halophilic heterotrophic nitrification-aerobic denitrification bacterium and its application on treatment of saline wastewater[J]. Bioresource Technology, 2015, 179: 421-428. DOI:10.1016/j.biortech.2014.12.057 |
| [28] | Wang P, Yuan Y Z, Li Q, et al. Isolation and immobilization of new aerobic denitrifying bacteria[J]. International Biodeterioration & Biodegradation, 2013, 76: 12-17. |
| [29] | Zheng H Y, Liu Y, Gao X Y, et al. Characterization of a marine origin aerobic nitrifying-denitrifying bacterium[J]. Journal of Bioscience and Bioengineering, 2012, 114(1): 33-37. DOI:10.1016/j.jbiosc.2012.02.025 |
| [30] | Yang X P, Wang S M, Zhou L X. Effect of carbon source, C/N ratio, nitrate and dissolved oxygen concentration on nitrite and ammonium production from denitrification process by Pseudomonas stutzeri D6[J]. Bioresource Technology, 2012, 104: 65-72. DOI:10.1016/j.biortech.2011.10.026 |
| [31] |
杨新萍, 钟磊, 周立祥. 有机碳源及DO对好氧反硝化细菌AD6脱氮性能的影响[J]. 环境科学, 2010, 31(6): 1633-1639. Yang X P, Zhong L, Zhou L X. Effect of carbon source and dissolved oxygen on denitrification by aerobic denitrifier Pseudomonas mendocina AD6[J]. Environmental Science, 2010, 31(6): 1633-1639. |