 2018, Vol. 39
2018, Vol. 39



大气中二氧化碳浓度增加对全球温度升高的影响越来越强烈, 关注全球尺度碳预算的文献和研究也日益增加[1].陆地生态系统由于下表面的复杂多样性和人类活动的强烈干扰, 造成了该生态系统碳循环具有明显地域特征和不确定性, 目前对陆地生态系统碳循环规律、源汇分布和影响因素的研究已经成为全球碳循环研究的主要内容[2~4].近些年在纬度70°N~30°S地区, 建立了以涡度相关为主要研究方法的通量网络, 实时对CO2、水和热通量进行了观测, 研究对象主要集中于温带针叶林、落叶阔叶林、常绿森林、一年生作物、草地、湿地和冻原[5].农田生态系统作为陆地生态系统的重要组成部分[6], 占全球陆地面积10.5%, 受到强烈的人为干扰, 是碳库中最活跃的部分[7].果园是农田生态系统重要土地利用类型, 开展果园生态系统碳收支和影响因素的研究, 是完整探讨整个陆地生态系统功能必不可少的内容.
近些年随着果树作物经济效益的提高, 我国水果优势产区大量麦田和玉米田已经转变为果园, 2015年中国果园种植面积达到1 282万hm2, 约占耕地面积的9.6%.我国是世界上苹果生产第一大国, 2015年的苹果种植面积232.8万hm2和产量4 261万t, 分别占世界苹果的[8]40.6%和42.8%.黄土高原地区因其降雨量少, 昼夜温差大, 光照时间长, 土层深厚等特殊环境条件, 已经成为我国苹果两大优势主产区之一, 栽培面积130万hm2和鲜果产量1 500万t·a-1, 分别占全国苹果总栽培面积和总产量的60%和55%[9].目前果园生态系统中应用涡度相关进行碳通量的研究相对较少, Rossi等[10]在意大利的猕猴桃园和Testi等[11]在西班牙的油橄榄园中有相关报道, 国内仅在葡萄园[12]和梨园[13]进行过这方面的研究.一方面果树作为多年生作物, 由于自身的特征, 有助于将碳长期储存在土壤中或者短期储存在木质部; 另一方面考虑到果树固定C的存储容量和种植面积的大小, 中国果园生态系统在陆地生态系统的C周期中发挥了重要作用, 并有助于碳汇[14].然而目前国内关于苹果园生态系统碳通量的研究较少, 且主要集中在单叶水平[15, 16]、果园土壤呼吸[17~20]等方面.关于定量中国果园生态系统尺度下的碳汇强度信息较少, 尤其是位于黄土高原地区的苹果园生态系统.
目前苹果园已经进行的碳通量研究, 时间上存在连续性差、尺度小的缺点, 针对以上现状, 本研究以陕西黄土高原地区集中连片的成龄苹果园生态系统为研究对象, 利用涡度相关通量塔和配套气象观测系统对果园内碳通量和气象因素进行连续观测(2016-01~2016-12), 开展苹果园下垫面与大气的碳交换研究, 来定量分析苹果园碳通量和环境影响因素在不同时间尺度上变化规律, 并对光合有效辐射PAR和不同层空气、土壤温度对NEE的影响进行探讨.与此同时, 与其它纬度相近的生态系统碳通量进行比较, 通过找出黄土高原苹果园碳存储的有力证据, 以期为准确估算我国黄土高原地区苹果园的碳汇强度提供数据支持, 也对区域碳收支总量的估计以及陆地生态系统碳通量估测的完善都具有重要意义.
1 材料与方法 1.1 研究区概况本试验果园位于苹果主产区陕西省咸阳市旬邑县太村镇(N35°08′52.845″ E108°18′00.561″), 如图 1所示, 试验开始于2015年12月.该地地处黄土高原渭北旱塬, 区域降水稀少且年际和年内分布不均, 主要降水分布在5~9月, 约占全年降水量的80%, 盛行风向为东南和西北方向, 海拔1 254 m.属暖温带大陆性气候, 温度适中, 雨热同期.当地光热资源丰富, 年日照时数2 390 h, 无霜期180 d, 年平均气温9.0℃, >10℃的积温2 955℃, 昼夜温差大.地下水埋深达70 m, 多年平均降水量589 mm, 平均水面蒸发量1 400 mm左右.容易出现当年冬季-次年春季的干旱.土壤类型以黑垆土为主, 耕层0~20 cm土壤颗粒组成为黏粒含量23%, 粉粒含量47%, 砂粒30%, 土壤有机质含量为1.06%, 总氮、磷和钾含量分别为2.18、0.83和12.59 g·kg-1.土壤pH为8.3, 干容重1.24 g·cm-3, 孔隙率51%, 田间持水量28.73%, 凋萎点12.3%.苹果树的主要物候期为萌芽(4月中旬)、开花(4月下旬)、果实形成(5~9月)、果实成熟(10月)和叶片脱落(11月).

|
图 1 试验地点在黄土高原的位置 Fig. 1 Location of experimental area in the Loess Plateau |
涡度相关系统位于果园中部偏东, 该地区苹果树平均高度3.6 m, EC系统观测高度位于冠层上方1.1 m, 风浪区长度从200~600 m不等, 满足试验要求.试验区域的地形为典型的黄土高原塬面区, 地势平坦.安装仪器下方为集中连片的苹果园, 面积350亩左右.当地在政府农业部门2003年统一规划下建园, 品种为长富二号.苹果行向为东西走向, 行距5 m, 株距3 m.树形采用疏散分层性, 中心领导干比较明显, 主枝5~7个, 分2~3层.试验地区果园的果树群体基本特征见表 1.该地区由于水资源限制在苹果生育期内不进行灌溉, 为典型的雨养农业区.
|
|
表 1 苹果园生态系统的果树基本特征表 Table 1 1 Basic features of apple trees in the apple orchard ecosystem |
1.2 研究方法
在通量塔4.7 m处安装涡度相关观测系统, 型号为Campbell的IRGASON一体式通量探头, 核心部件为三维超声风速仪(CSAT3A, Campbell Scientific Ltd., USA)和CO2/H2O红外气体分析仪(EC150, Campbell Scientific Ltd., USA), 分别测量风速和超声温度与CO2和H2O浓度.通量数据原始采样频率为10 Hz, 利用数据采集器(CR3000, Campbell Scientific Ltd., USA)记录、在线计算和存储30 min的通量值和其它相关参数.
通量塔上同时配套安装了气象观测系统, 分别安装有二维风速仪(Gill26, Gill Instruments Ltd., Lymington, England), 安装高度8 m, SI-111红外温度传感器(SI-111, Campbell Sci., USA)安装高度4 m, 2层空气温湿度传感器(HMP155A, Vaisala, Helsinki., Finlanda), 安装高度分别为4和8 m, 光合有效辐射传感器(PQS1, Campbell Scientific Ltd., USA)、四分量净辐射表(CNR4, Campbell Scientific Ltd., USA)、总辐射表(CMP3, Campbell Scientific Ltd., USA), 翻斗式雨量筒(TE525, Campbell Scientific Ltd., USA), 安装高度为5 m; 土壤温度传感器(109-L, Campbell Scientific Ltd., USA)和土壤含水量传感器(CS616, Campbell Scientific Ltd., USA)分别有4层, 埋设的深度分别为5、10、20和50 cm.二氧化碳探头(GMP343, Campbell Scientific Ltd., USA)安装高度70 cm, 土壤热通量板[HFP01(SC), Campbell Scientific Ltd., USA]4块, 分布在通量塔周围, 两块位于株间, 两块位于行间, 埋设深度为5 cm, 所有传感器都连接在数据采集器(CR3000, Campbell Scientific Ltd., USA)和扩展板AM25上, 30 min输出一组平均值. 30 min的气象数据和通量数据可以通过无线传输, 实现远程下载, 在通量塔周围安装远程监控, 用于观察果树物候期.
1.3 数据处理 1.3.1 CO2通量使用Eddypro 6.2软件(http://www.licor.com/env/products/eddy covariance)对10 Hz数据处理, 处理过程包括:①对10 Hz原始数据进行野点剔除[21]、二次坐标轴旋转[22]和水汽密度校正[23]等; ②对计算出的碳通量数据依次进行储存通量修正[24]、异常峰值剔除[25]、摩擦风速校正和缺失数据插补等.碳通量通过计算垂直方向风速与CO2密度脉动值的协方差获得, 转化为30 min通量数据.
1.3.2 数据插补在长期通量观测中, 由于仪器故障、系统校正及天气、风速等因素影响, 往往会造成观测数据丢失和异常, 为此, 还必须进行一系列的质量控制, 以保证观测数据的有效性.这主要包括:①湍流不充分(摩擦风速U*<0.15 m·s-1)利用周艳莲等[26]的方法确定U*的临界值为0.15 m·s-1; ②剔除降雨同期数据; ③阈值控制; ④异常突出数据(某一个数值与连续5点平均值之差的绝对值>5个点方差的2.5倍).使用非线性回归法进行数据拟合插补.具体方法为:利用Lloyd等[27]的方程拟合夜间(总辐射<1 W·m-1)碳通量, 即Reco.n与5 cm深度土壤温度(Ts)的关系, 插补生长季(4~11月)夜间和非生长季(12月~次年3月)全天缺失的碳通量, 进而外推至白天.
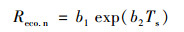
|
(1) |
式中, b1和b2是拟合系数, Reco.n是夜间生态系统的呼吸, 数值上Reco.n等于夜间生态系统NEE值NEEn[μmol·(m2·s)-1], Ts为5 cm的土壤温度.生态系统呼吸的温度敏感性系数(Q10)使用以下公式计算:
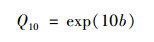
|
(2) |
Michaelis-Menten方程拟合日间(总辐射≥1 W·m-1)碳通量即净生态系统碳交换量(net ecosystem exchange, NEE)与光合有效辐射(photosynthetic active radiation, PAR)的关系[28].
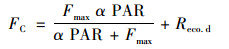
|
(3) |
式中, FC为白天净生态系统碳交换量NEEd[μmol·(m2·s)-1]; α为生态系统的表观量子效率(μmol·μmol-1); Fmax为最大净生态系统CO2交换[μmol·(m2·s)-1]; Reco.d为生态系统的暗呼吸速率[μmol·(m2·s)-1].
1.3.3 生态系统总生产力(GPP)和生态系统呼吸(Reco)计算果园生态系统中, 生态系统总初级生产力(GPP)为生态系统呼吸(Reco)(白天呼吸Reday与夜间呼吸Renight之和)与净生态系统交换量(NEE)之差.在碳生产力循环中, NEE提供净生态系统截获和释放能力的信息, NEE数值的正负表示生态系统光合和呼吸能力的强弱(NEE<0表示生态系统光合>呼吸, NEE>0表示生态系统光合<呼吸).涡度相关系统直接获取的是生态系统的NEE, 果园生态系统的GPP和Reco通过以下公式估算[28]:
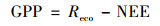
|
(4) |
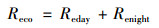
|
(5) |
利用Excel 2010进行数据剔除, 计算平均值, Sigmplot 12.5软件制作相关的图件.利用SPSS 23进行环境参数和NEE数据之间的拟合, 进行数据插补.
2 结果与分析 2.1 果园气象因子的年变化了解果园关键环境变量的季节变化, 对于研究果园与大气之间的碳交换至关重要. 图 2显示了每日4m空气温度最大值(Tmax)和最小值(Tmin)、日总光合有效辐射(PAR)、日总降雨量(PPT)、日均土壤体积含水量(VWC)、日均饱和水汽压差(VPD)、日均4 m、8 m高空气温度(Ta)、日均5 cm、10 cm土壤温度(Ts)和风向频率(FWD)季节变化, 果园各气象要素的变化具有明显的季节特征. Tmax、Tmin、Ta和Ts具有相似的变化趋势, 呈抛物线变化, 具有明显的峰值, Tmax在7月末出现最大值, Tmin、Ta和Ts最大值出现在8月中旬, 最小值都出现在1月末, 年平均值分别为14.8、6.8、10.9和11.5℃.在整个研究期间, Ts-5 cm和Ts-10 cm变化比Ta-4m和Ta-8m更加平稳.研究期间日总PAR最大值[60.22 mol·(m2·d)-1]出现在6月16日, 月总PAR最高值出现在7月, 最低值出现在12月, 降雨和阴天对辐射的影响很大.观测区的2016全年降雨量为379.6 mm, 根据降水量年性划分标准[29], 为枯水年. 5~9月降雨量为273.6 mm, 占全年降雨量的72%, 6月降雨量最高, 为110.5 mm, 12月降雨量最低, 为3.4 mm.降雨量随季节差异较为明显, 主要集中在夏季, 期间超过30 mm的两次降雨分别出现在6月11号(37.8 mm)和8月25号(38.4 mm).土壤含水量受降雨量和温度影响较为明显, 在3~11月保持在0.20 m3·m-3以上.日总量PAR和日平均VPD变化趋势较为一致, 也表现出明显的季节变化, VPD受降雨和辐射的影响非常明显.日平均VPD在6月29日达到了峰值2.34 kPa.风向频率分布图显示, 在全年的风向中东南和西北风向占据38%左右, 其次是东风向18%, 南风向15%左右, 北风向、东北风向、西风向和西南风向小于10%.主风向为西北和东南风向.

|
(a)每日4 m空气最大温度(Tmax)和最小温度(Tmin); (b)日总光合有效辐射(PAR); (c)日总降雨量(PPT)和日5, 10, 20和50 cm平均土壤含水量(VWC); (d)日均饱和水汽压差(VPD); (e)日均4 m和8 m空气温度(Ta-4 m and Ta-8 m)与5 cm和10 cm土壤温度(Ts-5cm and Ts-10 cm); (f)风向频率分布(FWD) 图 2 研究期间苹果园主要气象因子的季节变化 Fig. 2 Seasonal variation of major meteorological factor and apple orchard variables over the course of the study |
利用苹果园连续观测的通量数据, 以半小时为间隔, 分别计算每个月同一时刻NEE的月平均值, 得到苹果园生态系统1 a中不同月份NEE的月平均日变化图 3, 每个月的净生态系统CO2交换(NEE)平均日变化基本呈单峰曲线变化.其中生长季期间(4~11月)呈U形曲线变化趋势, 夜间大部分为正值, 白天为负值, 在非生长季(12月~次年3月), 生态系统的呼吸受到限制, NEE基本表现为正值, CO2通量的变化幅度小. NEE负值越小表示果园生态系统吸收的CO2量越大, 正值越大表示生态系统排放的CO2量越大.生长季期间(4~11月)5、6、7和8月NEE的日变化通量峰值都很接近, 分别为-15.48、-15.73、-16.99和-17.08 μmol·(m2·s)-1, 最大CO2吸收峰值出现在12:00左右, 最大的CO2排放峰值出现在夜间20:00~21:00左右, 分别为2.75、2.79、4.08和3.84 μmol·(m2·s)-1. 8月出现CO2的最大的吸收值, 但最大的CO2排放值出现在7月.非生长季(12月~次年2月), NEE值接近于0, 变化范围为0.30~0.68 μmol·(m2·s)-1, 3月的变化范围为0.48~1.48 μmol·(m2·s)-1, 整体上变化不明显, 其中3月的呼吸强度高于非生长季其它月份.

|
图 3 苹果园NEE的月平均日变化 Fig. 3 Diurnal variation of apple orchard monthly mean NEE |
通量数据处理后计算GPP、Reco和NEE的日总量和月总量.本研究中GPP、Reco和NEE日总量和月总量的季节性变化如图 4.苹果园在DOY101(日序, 下同)前后开始吸收碳, 在DOY325前后停止吸收碳. DOY101~136期间GPP呈线性增加, NEE值呈线性下降, 相比而言Reco的变化弱于GPP和NEE, 使得生态系统迅速变成强烈的碳汇, 且GPP明显高于Reco.日最大GPP、Reco和NEE分别为11.12、5.04和-7.34 g·(m2·d)-1. GPP和NEE在DOY136~244期间保持相对稳定, 之后开始缓慢下降, 到DOY325, 二者的值接近于0, 与月值变化趋势相一致, 而Reco日值的季节变化波动较大.冬季和早春, 在DOY1~101和325~366的期间即叶片脱落后至树木萌芽前, NEE和Reco速率很小, 果园通过土壤呼吸不断损失碳, 日GPP、Reco和NEE的值受降雨的波动较大, 如7月有降雨的天数为11 d, 降雨日GPP、Reco和NEE的平均值分别为6.14、2.75和-3.39 g·(m2·d)-1, 晴天日GPP、Reco和NEE的平均值分别为8.45、3.76和-4.68 g·(m2·d)-1.月GPP和NEE总量的变化和日总量的季节变化趋势相似, 在生长季初期4~5月迅速增加, 5~8月保持相对稳定, 8~11月缓慢下降.月GPP和Reco值最大值都出现7月分别为238.97 g·(m2·月)-1和105.38 g·(m2·月)-1, 而NEE的最大值出现在8月为-144.44 g·(m2·月)-1. 5、6、7和8月GPP和NEE月累积值非常接近.相比之下, 月Reco总量的变化先缓慢增加, 7月出现最大值, 后缓慢下降.在非生长季, NEE为正值且接近于Reco, 变化波动不大, GPP的值接近于0. NEE和Reco月总量的最大值出现在3月为20.39 g·(m2·月)-1.黄土高原成龄苹果园生态系统NEE月总量除非生长季12、1、2和3月为正值外(表现为碳源), 生长季(4~11月)都为负值(表现为碳汇).

|
图 4 苹果园GPP、Reco和NEE的日总量和月总量的季节变化 Fig. 4 Seasonal variation of daily and monthly total GPP, Reco and NEE of the apple orchard |
通过把日NEE、GPP和Reco总量累加得到图 5中苹果园累积GPP、Reco和NEE的季节变化.果园生态系统的GPP、Reco和NEE年累积总量分别为1223.2、525.2和-698 g·(m2·a)-1.累积GPP和NEE在萌芽前和落叶后的增长速率很慢, 同期与二者相比Reco的增长略微明显.在果树萌芽后, GPP和NEE的增长速率很快, 曲线的斜率接近于1, 在9月增长速度开始减小, 而Reco增长速率则要缓慢很多.累积NEE、GPP和Reco的变化趋势与NEE、GPP和Reco日和月总量的变化趋势相一致.

|
图 5 苹果园累积GPP、Reco和NEE的季节变化 Fig. 5 Seasonal variation of cumulative GPP, Reco, and NEE of the apple orchard |
光是光合作用的驱动者, 光合有效辐射是影响光合作用的主要因子之一.选取生长期内不同季节(春、夏和秋季)多个典型阴天和晴天半小时PAR和NEE数据作图, 探讨阴天和晴天对NEE的影响. 图 6(a)~6(c)显示苹果园NEE在不同季节阴天和晴天的日变化, 苹果园NEE值在晴天和阴天的变化曲线都为U形曲线.晴天变化相对剧烈, 最大NEE值出现在11:30~12:00, 分别为-19.89、-21.31和-17.39 μmol·(m2·s)-1.阴天NEE变化相对平稳, 最大NEE出现在11:00~13:00, 分别为-13.82、-16.68和-10.64 μmol·(m2·s)-1. 图 6(d)~6(f)显示苹果园在阴天和晴天PAR日变化. PAR变化趋势阴天为U形, 晴天为V形, 昼间由于乌云遮挡, 阴天PAR值显著低于晴天, 晴天的峰值分别为1 897.3、1 936.9和1 539.9 μmol·(m2·s)-1, 出现的时间为13:00~13:30, 阴天的峰值分别为456.8、1 214.5和655.4 μmol·(m2·s)-1, 出现的时间分别为11:00~13:00.在不同季节晴天和阴天的日变化中, 晴天最大NEE值和PAR的峰值出现时间并不一致, 而阴天则对应一致, 表明苹果树在晴天最大辐射值时出现微弱的光合午休.不同季节晴天的NEE和PAR值显著大于阴天, 其中夏季晴天与阴天的NEE和PAR差异最小.

|
图 6 苹果园PAR和NEE在不同时期晴天和阴天的的日变化 Fig. 6 Diurnal variation in the sunny and cloudy days in the apple orchard PAR and NEE for different periods |
苹果园的结构和功能在一年中发生明显的变化, 因此选择生长季(4~11月)以月为单位研究白天NEE如何响应PAR的变化, 如图 7所示.基于方程(3)的回归分析表明, PAR变化引起NEEd变化的相关性超过80%, 说明在生长季白天净碳吸收量主要受光合作用的影响, 而光合有效辐射直接影响光合作用效率.拟合生态系统表观量子效率(α)的变化范围为0.010 9~0.055 4 μmol·μmol-1, 量子产量完全在阔叶森林的公开数据[30]范围之内.拟合最大净生态系统交换Fmax和生态系统的暗呼吸速率Reco.d的变化范围分别为-10.35~-31.60 μmol·(m2·s)-1和0.54~3.28 μmol·(m2·s)-1, 最大值分别出现在8月和7月, 最小值都出现在11月.拟合参数Fmax和Reco.d数值的变化趋势与月NEE和Reco的变化趋势相似, 最大值出现的时间和月变化一致.

|
图 7 苹果园生长季(4~11月)白天生态系统净碳交换(NEEd)对光合有效辐射(PAR)的响应 Fig. 7 Light-response curves of daily net system exchange(NEEd) to photosynthetic active radiation(PAR) of the apple orchard growing season(Apr.-Nov.) |
温度对CO2交换的影响, 主要通过影响Reco发挥作用, 分为温度对根系及微生物活性的直接影响和温度对光合作用及碳素分布的间接影响, 继而进一步影响生态系统的净碳交换[31].夜间净生态系统CO2交换量NEE数值上等于Reco.n, 基于全年的数据, 选取U>0.15 m·s-1数据质量好的夜间NEE数据和相对应的气象数据, 利用指数方程(1)对Reco和5 cm、10 cm土壤温度以及在4 m和8 m高空气温度的关系进行拟合比较. 图 8生态系统呼吸和不同温度之间关系的结果表明, 0.5 h尺度的全年夜间果园生态系统呼吸与不同层土壤、空气温度之间主要呈现正相关指数关系, 相关系数表现为Ts-5 cm>Ts-10 cm>Ta-4 m>Ta-8 m, 苹果树各物候期夜间生态系统呼吸与土壤温度的相关性较高, 其中土壤呼吸对5 cm土壤温度的敏感系数Q10=2.365.

|
图 8 苹果园夜间生态系统呼吸(Reco.n)和不同层土壤、空气温度之间的关系 Fig. 8 Relationship between apple orchard nighttime ecosystem respiration(Reco.n) and different layers of soil and air temperature |
果园是农田生态系统的重要类型之一, 对完整陆地碳循环具有重要意义.本文以中国四大苹果主产区之一的黄土高原地区成龄苹果园为研究对象, 利用涡度相关系统, 基于全年(2016-01~2016-12)的通量观测数据和气象数据, 分析了黄土高原地区苹果园的碳通量特征在不同时间尺度上的变化特征以及主要气象因子对生态系统净CO2交换的影响.生长季(4~11月)内NEE白天为负值, 夜间为正值, 随温度升高, 辐射增强, 苹果树萌芽, 树冠叶幕逐渐形成, 苹果园生态系统碳汇能力迅速增加、持续时间逐渐延长, 光合和呼吸强度逐渐增加, 7月达到呼吸峰值, 8月达到净CO2吸收峰值, 之后随着温度下降, 辐射减弱, 叶片老化, 苹果树的光合和呼吸能力逐渐下降, 10月下旬苹果树叶片开始脱落, 到11月上旬净CO2吸收和排放降到最低值, 11月中旬左右叶片完全脱落.虽然日光合有效辐射在7月达到最大, 但由于7月出现连续的降雨和阴天, 影响了辐射, 进而影响了净CO2吸收.在非生长季(12月~次年3月), 由于树上没有叶片不进行光合作用, 生态系统只进行CO2排放, 温度降低进一步限制了土壤呼吸, 由于12月末至2月末土壤表层上冻, 土壤含水量低, 土壤呼吸进一步受到限制, 碳排放量很低, 此时NEE值接近于零. 3月温度升高, 地表温度>0℃, 土壤解冻, 苹果树的根系开始活动, 使得整个生态系统呼吸增强, 呼吸强度明显高于非生长季其它月份.在苹果树生长季内(4~11月), 净碳交换的日总量只有一个峰值[-7.34 g·(m2·d)-1], 与Guo等[12]研究的葡萄园和Zhang等[13]研究的梨园结果相一致, 而与Shen等[32]对华北平原冬小麦-夏玉米生态系统具有两个峰值不同.不同的碳交换特征可能是苹果树作为多年生果树与其他作物之间的物候期差异引起的.在没有高温、缺水和养分不足胁迫的条件下GPP数值的季节变化依赖于叶片截获PAR的数量[33], 在本研究中, 最大的CO2吸收出现在5~8月, 这个时期的光合作用约占全年光合作用的70%左右.在5~8月期间, 苹果树冠层叶幕几乎全部出现, 叶片的光合能力达到最强, PAR达到全年中的较高水平, 空气日平均温度15.6~25.2℃, 土壤的平均含水量0.25 m3·m-3, 降雨量243.6 mm. 7月Reco出现最大值, 在光合作用和呼吸作用的共同作用下, GPP也在此时达到最大值.
3.2 苹果园生态系统碳通量与其它生态系统的比较苹果园生态系统GPP、NEE和Reco年累积量分别为1 223.2、-698.0和525.2 g·(m2·a)-1, 表明黄土高原成龄苹果园具有较高的固碳能力, 为强烈的碳汇, 这与Wu等[14]对我国东、西、中部地区不同龄期苹果园碳汇潜力的调查结果一致, 即成龄苹果园具有较高的碳汇潜力.如表 2所示, 与人工林[34]相比, 苹果园生态系统具有较高固碳潜力的原因是研究对象为果园生态系统, 人为进行的修剪、施肥和除草等各种管理措施, 都使得苹果树向积累更多的碳、产生更大的经济效益方向生长.与纬度相近的其它果园生态系统相比较, 苹果园生态系统每年NEE值(绝对值)低于西北旱区武威酿酒葡萄园[12], 在单叶水平葡萄和苹果的光合能力差异并不明显, 如孙文泰等[15]测定长富二号苹果单叶光合为16.93 μmol·(m2·s)-1, 刘玉兰等[35]测定酿酒葡萄苹果单叶光合为15 μmol·(m2·s)-1左右, 产生差异的原因可能与两个试验地区不同PAR值和日照时数有关, 本试验地区年日照时数2 390 h, 武威地区日照时数3 000 h以上; 而略高于林型相近的梨园[13], 但是低于地中海气候下种植的常绿果树橄榄树[5], 造成的原因可能是梨园每年约6个月显示负的NEE值, 而苹果园在8个月显示负的NEE值, 油橄榄种植园长的碳吸收期, 全年12个月表现为碳汇.与纬度相近的一年生农田生系统[36]相比, 苹果园NEE值(绝对值)显著大于该生态系统, 显示了苹果园更强的碳汇能力.苹果树强的碳汇特性可能和它的光合特性和物候期有关.苹果适宜种植在光照时间长, 辐射强和昼夜温差大的地方, 夜间的低温环境会抑制呼吸, 使得夜间的正的NEE值更小.
|
|
表 2 不同生态系统NEE比较 Table 2 Comparison of net CO2 exchange for different ecosystems |
3.3 光合有效辐射和不同温度对苹果园生态系统碳通量的影响
苹果园NEE在不同季节晴天和阴天的日变化中, 当晴天光合有效辐射达到最大值时观察到不明显的光合午休现象, 没有出现明显双峰型曲线, 这与孙文泰等[15]在陇东旱地果园中苹果光合的研究结果相似, 与Guo等[12]研究武威西北旱区的葡萄园和孙霞等[16]在新疆地区苹果园中观察到明显的光合午休现象的结果即双峰型曲线不一致, 这可能与两个地区生长季日间最高温度和辐射强弱有关.苹果园生态系统白天CO2通量对PAR的响应结果表明, 直角双曲线方程能很好地用来描述果园生态系统PAR和白天净CO2的交换的关系, 二者之间的相关系数达到0.8以上.同时也说明, 在生长季辐射强弱是影响果园生态系统白天净CO2交换最主要的因素, 这一点和净碳交换在晴天和阴天的日变化中也可以看出.与唐祥等[34]研究人工林和朱咏莉等[37]研究稻田的结果相一致.本研究分析了夜间生态系统呼吸与不同层次土壤、空气温度的响应, 相关系数的大小表现为Ts-5 cm>Ts-10 cm>Ta-4 m>Ta-8 m, 其中Reco.n与5 cm土壤温度的相关性最好, 这与牛晓栋等[38]的研究报导结果相一致.苹果园生态系统年Q10=2.365趋向于生态系统[39]Q10平均幅度范围(1.3~5.6)的中间值.
4 结论(1) 黄土高原成龄苹果园生态系统NEE的日变化基本呈现单峰曲线变化, 生长季内12:00左右出现通量峰值, 日变化具有很大的季节差异.月总量在生长季(4~11月)为碳汇, 呈“U”形变化趋势, NEE月总量的大小依次为8月>6月>5月>7月>9月>10月>4月>11月, Reco月总量的大小依次为7月>8月>6月>5月>9月>10月>4月>11月.非生长季(12月~次年3月)表现为碳源, 碳源能力较弱, Reco月总量的大小依次为3月>12月>1月>2月.
(2) 苹果园生态系统的碳通量在生长季变化幅度大, 非生长季变化不明显, 全年表现为“一峰两谷”的变化趋势.直角双曲线方程能对苹果园生态系统生长季白天净碳通量与光合有效辐射的响应进行很好描述, 二者之间的相关性达到80%以上.生态系统呼吸速率随不同层次空气、土壤温度的升高呈正指数增加, 其中5cm土壤温度与苹果园生态系统呼吸之间的相关性最高, Q10值为2.365.
(3) 苹果园的年净生态系统碳交换为-698.0g·(m2·a)-1, 与纬度相近的其它生态系统相比, 苹果园生态系统具有较高的固碳能力和生态价值.
致谢: 感谢国家苹果产业技术体系咸阳综合试验站查养良站长和旬邑县园艺站辛选民站长在仪器安装过程中的帮助.| [1] | Alexander L, Allen S, Bindoff N L, et al. Summary for policymakers[A]. In: IPCC (Ed. ). Climate Change 2013: The Physical Science Basis. Working Group Ⅰ Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge, United Kingdom and New York: Cambridge University Press, 2013. 710-719. |
| [2] | 李克让. 土地利用变化和温室气体净排放与陆地生态系统碳循环[M]. 北京: 气象出版社, 2002: 35-48. |
| [3] | Baldocchi D D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems:past, present and future[J]. Global Change Biology, 2003, 9(4): 479-492. DOI:10.1046/j.1365-2486.2003.00629.x |
| [4] | Schimel D S, House J I, Hibbard K A, et al. Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems[J]. Nature, 2001, 414(6860): 169-172. DOI:10.1038/35102500 |
| [5] | Nardino M, Pernice F, Rossi T, et al. Annual and monthly carbon balance in an intensively managed Mediterranean olive orchard[J]. Photosynthetica, 2013, 51(1): 63-74. DOI:10.1007/s11099-012-0079-6 |
| [6] |
宋长春. 湿地生态系统碳循环研究进展[J]. 地理科学, 2003, 23(5): 622-628. Song C C. Advance in research on carbon cycling in wetlands[J]. Scientia Geographica Sinica, 2003, 23(5): 622-628. |
| [7] |
韩广轩, 周广胜, 许振柱. 中国农田生态系统土壤呼吸作用研究与展望[J]. 植物生态学报, 2008, 32(3): 719-733. Han G X, Zhou G S, Xu Z Z. Research and prospects for soil respiration of farmland ecosystems in China[J]. Journal of Plant Ecology, 2008, 32(3): 719-733. |
| [8] | 中华人民共和国国家统计局. 国家数据[EB/OL]. http://data.stats.gov.cn/gwwz.htm,2016-01-19. |
| [9] | Wang R, Guo S L, Jiang J S, et al. Tree-scale spatial variation of soil respiration and its influence factors in apple orchard in Loess Plateau[J]. Nutrient Cycling in Agroecosystems, 2015, 102(2): 285-297. DOI:10.1007/s10705-015-9699-0 |
| [10] | Rossi F, Facini O, Georgiadis T, et al. Seasonal CO2 fluxes and energy balance in a kiwifruit orchard[J]. Italian Journal of Agrometeorology, 2007, 1: 44-56. |
| [11] | Testi L, Orgaz F, Villalobos F. Carbon exchange and water use efficiency of a growing, irrigated olive orchard[J]. Environmental and Experimental Botany, 2008, 63(1-3): 168-177. DOI:10.1016/j.envexpbot.2007.11.006 |
| [12] | Guo W H, Kang S Z, Li F S, et al. Variation of NEE and its affecting factors in a vineyard of arid region of northwest China[J]. Atmospheric Environment, 2014, 84: 349-354. DOI:10.1016/j.atmosenv.2013.11.057 |
| [13] | Zhang Y C, Shen Y J, Xu X L, et al. Characteristics of the water-energy-carbon fluxes of irrigated pear (Pyrus bretschneideri Rehd) orchards in the North China Plain[J]. Agricultural Water Management, 2013, 128: 140-148. DOI:10.1016/j.agwat.2013.07.007 |
| [14] | Wu T, Wang Y, Yu C J, et al. Carbon sequestration by fruit trees-Chinese apple orchards as an example[J]. PLoS One, 2012, 7(6): e38883. DOI:10.1371/journal.pone.0038883 |
| [15] |
孙文泰, 赵明新, 尹晓宁, 等. 陇东旱地果园地表覆盖方式对苹果光合特性的影响[J]. 干旱地区农业研究, 2013, 31(3): 19-25, 60. Sun W T, Zhao M X, Yin X N, et al. Effects of different mulch modes on apple photosynthesis in dry-land orchards of east Gansu Province[J]. Agricultural Research in the Arid Areas, 2013, 31(3): 19-25, 60. |
| [16] |
孙霞, 柴仲平, 蒋平安. '红富士'苹果光合特性日变化特征研究[J]. 北方果树, 2010(5): 4-6. Sun X, Chai Z P, Jiang P A. Study the diurnal variation of photosynthetic characteristics on 'Red Fuji' apples[J]. Northern Fruits, 2010(5): 4-6. |
| [17] |
贾曼莉, 郭宏, 杜毅飞, 等. 生草对渭北黄土高原苹果园土壤呼吸的影响[J]. 草地学报, 2014, 22(3): 518-527. Jia M L, Guo H, Du Y F, et al. Influences of interplanting herbage on the soil respiration of apple orchard in Weibei Loess Plateau[J]. Acta Agrestia Sinica, 2014, 22(3): 518-527. |
| [18] |
李泽. 渭北旱塬苹果园土壤呼吸的时空变异特征及其影响因子[D]. 杨陵: 西北农林科技大学, 2012. 31-34. Li Z. Spatio-temporal character of soil respiration and its influence factors in apple orchard of Weibei dryland[D]. Yangling: Northwest A&F University, 2012. 31-34. http://cdmd.cnki.com.cn/Article/CDMD-10712-1012436762.htm |
| [19] |
王蕊, 郭胜利, 刘庆芳, 等. 苹果园土壤呼吸的变化及生物和非生物因素的影响[J]. 环境科学, 2014, 35(5): 1915-1921. Wang R, Guo S L, Liu Q F, et al. Variation characteristic in soil respiration of apple orchard and its biotic and abiotic influencing factors[J]. Environmental Science, 2014, 35(5): 1915-1921. |
| [20] | 张红星. 黄土高原小麦田和苹果园生态系统土壤呼吸研究[D]. 北京: 中国科学院大学, 2009. 28-55. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2165126 |
| [21] | Vickers D, Mahrt L, Blodgett P. Quality control and flux sampling problems for tower and aircraft data[J]. Journal of Atmospheric and Oceanic Technology, 1997, 14(3): 514-526. |
| [22] | Nakai M, Kashiwazaki N, Takizawa A, et al. Morphologic changes in boar sperm nuclei with reduced disulfide bonds in electrostimulated porcine oocytes[J]. Reproduction, 2006, 131(3): 603-611. DOI:10.1530/rep.1.01001 |
| [23] | Webb E K, Pearman G I, Leuning R. Correction of flux measurements for density effects due to heat and water vapour transfer[J]. Quarterly Journal of the Royal Meteorological Society, 1980, 106(447): 85-100. DOI:10.1002/(ISSN)1477-870X |
| [24] | Zha T S, Kellomäki S, Wang K Y, et al. Carbon sequestration and ecosystem respiration for 4 years in a scots pine forest[J]. Global Change Biology, 2004, 10(9): 1492-1503. DOI:10.1111/gcb.2004.10.issue-9 |
| [25] | Papale D, Reichstein M, Aubinet M, et al. Towards a standardized processing of net ecosystem exchange measured with eddy covariance technique:algorithms and uncertainty estimation[J]. Biogeosciences, 2006, 3(4): 571-583. DOI:10.5194/bg-3-571-2006 |
| [26] | 周艳莲, 孙晓敏, 朱治林, 等. 几种不同下垫面地表粗糙度动态变化及其对通量机理模型模拟的影响[J]. 中国科学D辑地球科学, 2006, 36(增刊1): 244-254. |
| [27] | Lloyd J, Taylor J A. On the temperature dependence of soil respiration[J]. Functional Ecology, 1994, 8(3): 315-323. DOI:10.2307/2389824 |
| [28] | 于贵瑞, 孙晓敏. 陆地生态系统通量观测的原理与方法[M]. 北京: 高等教育出版社, 2006: 255-260. |
| [29] |
张北赢, 徐学选, 刘文兆, 等. 黄土丘陵沟壑区不同降水年型下土壤水分动态[J]. 应用生态学报, 2008, 19(6): 1234-1240. Zhang B Y, Xu X X, Liu W Z, et al. Dynamic changes of soil moisture in loess hilly and gully region under effects of different yearly precipitation patterns[J]. Chinese Journal of Applied Ecology, 2008, 19(6): 1234-1240. |
| [30] | Ruimy A, Jarvis P G, Baldocchi D D, et al. CO2 fluxes over plant canopies and solar radiation:a review[J]. Advances in Ecological Research, 1995, 26: 1-68. DOI:10.1016/S0065-2504(08)60063-X |
| [31] | Davidson E A, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest[J]. Global Change Biology, 1998, 4(2): 217-227. DOI:10.1046/j.1365-2486.1998.00128.x |
| [32] | Shen Y J, Zhang Y C, Bridget R, et al. Energy/water budgets and productivity of the typical croplands irrigated with groundwater and surface water in the North China Plain[J]. Agricultural and Forest Meteorology, 2013, 181: 133-142. DOI:10.1016/j.agrformet.2013.07.013 |
| [33] | Auzmendi I, Marsal J, Girona J, et al. Daily photosynthetic radiation use efficiency for apple and pear leaves:seasonal changes and estimation of canopy net carbon exchange rate[J]. European Journal of Agronomy, 2013, 51: 1-8. DOI:10.1016/j.eja.2013.05.007 |
| [34] |
唐祥, 陈文婧, 李春义, 等. 北京八达岭林场人工林净碳交换及其环境影响因子[J]. 应用生态学报, 2013, 24(11): 3057-3064. Tang X, Chen W J, Li C Y, et al. Net carbon exchange and its environmental affecting factors in a forest plantation in Badaling, Beijing of China[J]. Chinese Journal of Applied Ecology, 2013, 24(11): 3057-3064. |
| [35] |
刘玉兰, 郑有飞, 张晓煜, 等. 光质和光强对酿酒葡萄光合速率及糖分积累的影响[J]. 中国农业气象, 2006, 27(4): 286-288, 292. Liu Y L, Zheng Y F, Zhang X Y, et al. A study on impacts of light conditions on net photosynthesis and sugar accumulation of wine grape[J]. Chinese Journal of Agrometeorology, 2006, 27(4): 286-288, 292. |
| [36] | 于贵瑞, 孙晓敏. 中国陆地生态系统碳通量观测技术及时空变化特征[M]. 北京: 科学出版社, 2008: 544-546. |
| [37] |
朱咏莉, 吴金水, 童成立, 等. 稻田CO2通量对光强和温度变化的响应特征[J]. 环境科学, 2008, 29(4): 1040-1044. Zhu Y L, Wu J S, Tong C L, et al. Responses of CO2 fluxes to light intensity and temperature in rice paddy field[J]. Environmental Science, 2008, 29(4): 1040-1044. |
| [38] |
牛晓栋, 江洪, 张金梦, 等. 浙江天目山老龄森林生态系统CO2通量特征[J]. 应用生态学报, 2016, 27(1): 1-8. Niu X D, Jiang H, Zhang J M, et al. Characteristics of CO2 flux in an old growth mixed forest in Tianmu Mountain, Zhejiang, China[J]. Chinese Journal of Applied Ecology, 2016, 27(1): 1-8. |
| [39] | Tjoelker M G, Oleksyn J, Reich P B. Modelling respiration of vegetation:evidence for a general temperature-dependent Q10[J]. Global Change Biology, 2001, 7(2): 223-230. DOI:10.1046/j.1365-2486.2001.00397.x |