 2018, Vol. 39
2018, Vol. 39



2. 湿润亚热带生态-地理过程教育部重点实验室, 福州 350007;
3. 福建师范大学亚热带湿地研究中心, 福州 350007
2. Key Laboratory of Humid Sub-tropical Eco-geographical Process of Ministry of Education, Fuzhou 350007, China;
3. Research Centre of Wetlands in Subtropical Region, Fujian Normal University, Fuzhou 350007, China
近年来, 大气库中温室气体增温效应加强引起的全球气候变暖已成为国际社会公认的全球性问题之一[1].甲烷(CH4)和氧化亚氮(N2O)是大气中2种重要的温室气体, 自1750年以来, 其浓度分别增加了150%和20%, 达到了目前的1 803 nL·L-1和324 nL·L-1[1].由于两种温室气在大气库中快速地增加和较高的辐射增温潜势, 不同地表生态系统CH4和N2O排放通量的研究一直是全球变化研究的焦点和核心内容之一[2~4].
陆地水生生态系统作为全球重要生态系统之一, 已被证实是大气库中CH4的重要来源, 在全球温室气体排放中居于重要地位[5, 6].小型池塘作为一类重要的水生生态系统, 其总面积仅占全球水体分布总分布面积的8.6%, 但其CH4排放比例却占到内陆水体总CH4排放的40%[7].基于以上原因, 小型池塘逐渐成为温室气体排放的研究热点[8, 9].但目前已有相关研究主要聚集在水位相对较高时期的水-气界面CH4和N2O通量方面[8~12], 对于一些处于低水位时期或短暂排干的池塘, 特别是受人类活动显著影响的陆基水产养殖塘的关注甚少.
据Verdegem等的估算[13], 全球咸、淡水养殖塘面积约为1.11×105 km2, 其中中国水产养殖塘总面积约为0.59×105 km2, 位居世界第1位.由于养殖过程中, 养殖生物的排泄物、残存饵料和生物尸体等有机物会沉积池底, 加上池中的有机碎屑、死亡的藻类等, 会造成池底老化、水质恶化[14].因此, 为确保一个良好的水产养殖环境, 诸多水产养殖塘中的水体往往在一个养殖周期结束后被排出, 进行短暂的自然晒塘、清淤等管理活动[14].排水活动引起的池塘水位急剧下降势必会对其内部的营养物质循环、沉积物理化性质及整个系统功能产生显著影响.显然, 这种影响对养殖池塘沉积物中温室气体产生与排放过程的改变亦具有不可忽视的作用.对农田和湿地的研究发现[15~19], 排水活动引起的水位降低可显著减少CH4排放但能够显著增加N2O释放.然而, 对于排水后的陆基水产养殖CH4和N2O释放情况并不知晓.积极开展该方面的研究, 以期为今后深入认识水产养殖塘碳氮循环、完善温室气体清单编制、相应减排措施制定和气候预测模型的改良提供数据支撑和科学依据.因此, 本研究于2012年12月中旬在闽江河口区选取一个未排干养殖塘和排干养殖塘为研究对象, 对其CH4和N2O交换通量进行了连续24 h原位观测, 主要探讨以下两个问题:①排干初期, 日变化观测尺度上的养殖塘排水活动是否会显著减少CH4排放并可显著增加N2O释放; ②排干初期, 排水后的养殖塘CH4和N2O通量日变化特征与未排干养殖是否存在差异.
1 材料与方法 1.1 采样站位和采样时间研究区域位于闽江河口鳝鱼滩湿地, 地理坐标为26°00′36″N~26°03′42″N, 119°34′12″E~119°40′40″E(图 1).气候为中亚热带和南亚热带海洋性季风气候的过渡区, 温暖湿润, 多年平均气温和降水量分别为19.7℃和1 346 mm, 年降水日数为153 d, 主要集中于3~9月[20].该区是典型的开放式感潮河口, 潮汐属于典型半日潮, 土壤以潮汐盐土为主.土著物种芦苇(Phragmites australis)、短叶茳芏(Cyperus malaccensis Lam.var.brevifolius Bocklr.)和外来入侵物种互花米草(Spartina alterniflora)为该区域主要的优势植物群落.陆基池塘养殖是区域内的主要农业生产活动, 其水源包括天然降水、河水和潮水.

|
图 1 研究区域和采样点位置示意 Fig. 1 Location of the sampled ponds in the Minjiang River estuary |
研究样地选取在鳝鱼滩湿地中偏西部五门闸附近堤坝内侧的养殖池塘进行.该区域养殖塘类型主要有鱼虾混合养殖塘和养虾塘两种.通常, 鱼虾混养塘在5月初投放虾苗和鱼苗, 11月中旬左右将虾全部捕获, 2月底鱼被全部捕获, 随后池塘水全部释放至附近的海域; 养虾塘于5月初投放虾苗, 11月中旬左右虾被全部捕获, 12月初开始将池塘水释放至附近海域, 进行短暂晒塘处理.本研究在获得养殖户准许情况下, 分别选择了一个未排干的鱼虾混养塘和已排干的养虾塘为研究对象(两种养殖塘底部均未出现水生植物生长现象), 以此开展养殖塘排干活动对温室气体通量日变化的影响.鱼虾混养塘平均水深为1.35 m, 面积为1 100 m2, 养殖对象为草鱼(Ctenopharyngodon idellus)和白对虾(Litopenaeus vannamei), 鱼养殖密度为5尾·m-2和虾35尾·m-2, 每天07:00、11:00和17:00进行饵料投喂, 投喂的饵料为人工配置饵料(粤海牌, 广州, 中国), 投放强度(冬季养殖期间)范围介于为20~60 kg·d-1; 养虾塘面积为1 200 m2, 研究期间的水深约0.15 m, 池塘的日常管理活动(如饵料投喂、水体增氧等)因虾全部被捕获和池塘水被释放而停止.在上述每种类型池塘, 布设4个样品采样点, 采样时间于2012年12月18日07:00至19日07:00完成.鉴于人员不足(野外采样仅有两个人)和安全问题, 样品采集在白天设定每间隔3 h进行1次, 在夜间设定每间隔4 h进行1次.
气样采集运用目前国际上通用的箱法来完成.未排干的养殖塘是利用悬浮箱进行气样采集, 规格为直径25 cm×高40 cm的底部开口圆筒, 主体部分为透明有机玻璃材质; 排干的养殖塘是利用静态箱进行气样采集, 箱体由顶箱(35 cm长×35 cm宽×60 cm高)和带有边槽的底座(35 cm长×35 cm宽×35 cm高)两部分组成, 材质分别为透明有机玻璃和PVC材料.底座需要在样品采集前一周埋进沉积物中, 埋入的深度为30 cm, 出露沉积物表面约5 cm.每个采集箱内部的顶部安装1个小风扇用于混匀箱内气体, 侧面安装一个气体采样口.气样用100 mL带有三通阀的注射器从箱内抽取50 mL, 1个通量测定共抽气4次, 分别在采样箱覆盖后第0、15、30和45 min采集, 并将抽取的气体注入50 mL气体采集袋中(德林, 大连), 遮光保存运回实验室待测定分析.
1.3 气样分析与通量估算气袋中的CH4与N2O分别采用GC-2010和GC-2014气相色谱仪(岛津, Japan)测定分析, 并根据以下公式计算养殖塘与大气库之间的CH4和N2O交换通量[F, mg·(m2·h)-1][10]:
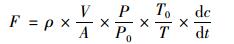
|
式中, ρ为标准状态下温室气体的密度(mg·m-3); V和A分别为箱内空气体积(m3)和采样箱覆盖面积(m2); P和T分别为采样点处的实测气压值(hPa)和采样时刻的箱内绝对温度(K); T0和P0为标准状态下的空气绝对温度(273.15 K)和气压值(1 013.25 hPa); dc/dt为采样期间箱内某种温室气体的浓度变化率.气体通量是负值时, 表示气体由大气库进入养殖塘中; 正值时, 表示气体从养殖塘释放到大气库中.
1.4 水样采集与测定分析与气样采集同步, 在未排干养殖塘的每个悬浮箱附近利用100 mL注射器采集表层水体(10 cm深)3份:1份水样用于水体N、P营养盐浓度的测定分析, 1份水样用于水体叶绿素a(Chlorophyll a, Chl-a)浓度的测定分析, 1份水样用于水体中SO42-浓度的测定分析.用于营养盐和SO42(浓度分析的水样均分别装入100 mL白色聚乙烯瓶, 用于水体叶绿素a浓度分析的水样需装入100 mL棕色聚乙烯塑瓶中.以上水样装入相应样品瓶后, 需立刻加入饱和HgCl2溶液1 mL, 用于抑制水样中的微生物活性[21, 22].水样中营养盐浓度的测定需用孔径为0.45 μm滤纸过滤, 过滤的水样装置于清洁的100 mL白色聚乙烯瓶中, 运用流动注射分析仪(SKALAR San++, Netherlands)分析测定水样中NO3--N、NH4+-N、PO43-指标.水体叶绿素a浓度分析的样品先经过丙酮(纯度90%)浸提, 然后将提取液通过紫外-可见分光光度计(岛津UV-2450, Japan)测定分析上清液吸光值.叶绿素a浓度计算公式参见文献[23~25], 最终结果以μg·L-1表示.采用铬酸钡分光光度法测定水样中的SO42-浓度.
1.5 环境因子原位测定在每个采样时刻, 采用便携式仪器对主要气象参数(气温、气压和风速)和表层沉积物10 cm深的温度、pH值、氧化还原电位Eh和电导率EC进行原位测定.气温、气压和风速采用便携式气象仪(Kestrel-3500, USA)测定; 沉积物温度、pH值和氧化还原电位Eh采用IQ150便携式pH/氧化还原电位/温度计(IQ Scientific Instruments, USA); 沉积物电导率采用2265FS便携式电导/温度计(Spectrum Technologies Inc, USA)测定.水中溶解氧采用多参数水质监测仪(HORIBA, Japan)测量, 测定深度为水面下10 cm.
1.6 数据处理与统计分析采用Excel 2003对原始数据进行均值及标准误的计算处理.利用SPSS 17.0统计软件包中重复测量方差分析法(Repeated measures ANOVA)检验未排干与排干养殖塘之间的温室气体通量、环境因子差异性. CH4或N2O交换通量与各个测定环境因子间的相关关系利用SPSS 17.0中的Pearson相关分析法进行统计分析.文中所有数据作图均通过OriginPro 7.5完成.
2 结果与分析 2.1 养殖塘排干对CH4通量日变化的影响观测期间, 未排干和排干养殖塘均是大气库中CH4释放源, 日变异系数(coefficient of variation, CV)分别为61%和46%, 呈现出明显的日变化特征(图 2).在白天, 未排干与排干养殖塘向大气库释放CH4,通量强度随时间推移表现出基本一致的变化趋势, 即从观测初始时刻07:00开始增加, 至下午13:00达到最大值, 随后开始降低; 但在夜间, 未排干与排干池塘向大气库释放CH4通量强度表现出不一致的时间变化趋势, 其中未排干池塘释放CH4通量强度呈现先增加后降低的特征[图 2(a)], 而排干池塘呈现随时间推移逐渐降低的趋势[图 2(b)].在日变化尺度上, 未排干池塘与排干池塘释放CH4通量的变化范围分别介于(0.04±0.01)~(0.10±0.01) mg·(m2·h)-1[图 2(a)]和(14.04±5.24)~(33.72±13.07) mg·(m2·h)-1[图 2(b)], 均值分别为(0.07±0.01) mg·(m2·h)-1和(24.74±2.33) mg·(m2·h)-1.重复测量方差分析结果显示, 排干养殖塘在各个观测时段向大气库释放CH4通量均极显著高于未排干养殖塘(P<0.01), 表明冬季养殖塘排干活动可以显著增加其CH4的释放强度.

|
图中阴影部分表示夜间, 下同 图 2 初冬时期未排干与排干养殖塘CH4通量日变化特征 Fig. 2 Diurnal variations of CH4 fluxes from the undrained and drained aquaculture pond in early winter |
图 3显示, 未排干与排干养殖塘N2O通量表现出不一样的昼夜变化规律.观测期间, 未排干养殖塘与大气库N2O通量强度从观测初始时刻07:00开始增加, 至13:00达到最大值, 随后开始降低, 至16:00以后又开始增加, 但增幅较小, 整体上呈现出夜间通量高于白天的特征[(0.006±0.003) mg·(m2·h)-1和(-0.001±0.007) mg·(m2·h)-1, 图 3(a)]; 排干池塘与大气库N2O通量强度从观测初始时刻07:00开始增加, 至13:00达到高峰值, 之后随时间推移而逐渐降低, 整体上表现出白天释放通量显著高于夜间的特征[(1.20±0.21) mg·(m2·h)-1和(0.84±0.10) mg·(m2·h)-1, P<0.05, 图 3(b)].在日变化尺度上, 未排干池塘与排干池塘N2O通量的变化范围分别介于(-0.027±0.011)~(0.011±0.003) mg·(m2·h)-1[图 3(a)]和(0.59±0.13)~(1.76±0.30) mg·(m2·h)-1, 均值分别为(0.002±0.004) mg·(m2·h)-1和(1.07±0.15) mg·(m2·h)-1[图 3(b)], 前者呈现出吸收-排放交替的特征, 后者全天表现大气库N2O释放源.重复测量方差分析结果显示, 排干养殖塘在各个观测时段向大气库释放N2O通量均极显著高于未排干养殖塘(P<0.01), 表明冬季养殖塘排干活动亦可以显著增加其N2O的释放.

|
图 3 初冬时期未排干与排干养殖塘N2O通量日变化特征 Fig. 3 Diurnal variations of N2O fluxes from the undrained and drained aquaculture pond in early winter |
观测期间, 研究样地气象参数、未排干池塘水体营养盐浓度及SO42-浓度的日变化特征分别见图 4和图 5.养殖塘所在区域的气温变化随观测时间推移呈现先增加, 至13:00达到最大值, 随后开始降低的特征, 而风速和气压随时间推移具有较明显的波动变化特征(图 4).未排干池塘水体NO3--N浓度从观测初始时刻07:00至第二天04:00呈现随时间推移而降低的趋势, 随后开始增加[图 5(a)]; 水体NH4+-N浓度从观测初始时刻至16:00随时间推移呈现下降趋势, 之后时刻NH4+-N出现增加, 但波动性较小[图 5(a)]; 水体PO43--P浓度在观测期间随时间变化波动性较大, 呈现增加-降低-增加-降低的特征[图 5(b)]; 水体SO42-浓度从观测初始时刻07:00~00:00随时间变化较小, 之后出现较大幅度的降低[图 5(b)]. 图 6为未排干与排干养殖塘沉积物温度等环境因子日变化特征.观测期间, 两类养殖塘沉积物温度、pH值、氧化还原电位Eh和电导率EC均表现出较为明显的日变化动态, 其中未排干池塘上述环境因子指标波动幅度较排干养殖塘大.差异性分析结果显示, 未排干与排干养殖塘沉积物温度在观测期间差异不显著(P>0.05), 但沉积物pH值和电导率EC均呈现未排干养殖塘显著高于排干养殖塘(P<0.05), 沉积物氧化还原电位呈现前者显著低于后的特征(P<0.05)(图 6).

|
图 4 初冬时期养殖塘气象参数日变化特征 Fig. 4 Diurnal variations of meteorological parameters from the aquaculture pond in early winter |

|
图 5 初冬时期未排干养殖塘水体营养盐、SO42-、溶解氧(DO)和叶绿素a浓度日变化特征 Fig. 5 Diurnal variations of water nutrients, SO42-, dissolved oxygen, and Chl-a concentration from the undrained aquaculture ponds in early winter |

|
图 6 初冬时期养殖塘沉积物温度、pH、电导率(EC)和氧化还原电位(Eh)日变化特征 Fig. 6 Diurnal variations of sediment temperature, pH, conductivity, and redox potential from the aquaculture ponds in early winter |
相关分析结果显示(表 1和表 2), 未排干池塘CH4释放通量与气温、风速和沉积物温度均呈现出极显著正相关关系(P<0.01), 与沉积物电导率EC呈现极显著负相关关系(P<0.01), N2O通量只与沉积物温度呈现显著正相关关系(P<0.05), 但与其它环境因子相关性未达到显著水平(P>0.05);排干池塘CH4释放通量与各测定的环境因子相关性均不显著(P>0.05), N2O通量分别与气温、沉积物温度和氧化还原电位Eh呈现显著(P<0.05)或极显著(P<0.01)正相关关系; 综合两个养殖塘发现, CH4、N2O通量均与pH、电导率呈现出极显著负相关关系(P<0.01), 与氧化还原电位Eh呈现极显著正相关关系(P<0.01).
|
|
表 1 养殖塘CH4通量与环境因子的相关分析1) Table 1 Correlation coefficient between CH4 fluxes and environmental factors |
|
|
表 2 养殖塘N2O通量与环境因子的相关分析 Table 2 Correlation coefficient between N2O fluxes and environmental factors |
3 讨论 3.1 排水活动对河口区养殖塘温室气体通量的影响
CH4是由土壤或沉积物中多种细菌形成的复杂食物网对土壤或沉积物中有机物进行厌氧分解, 在最终形成CH4产生底物的情况下, 由多种厌氧产甲烷古菌协助生成[26].水位(水深)条件作为影响湿地生态系统土壤或水生生态系统沉积物氧化/还原状况的重要因子, 其变化对土壤中微生物参与下的有机物分解和甲烷代谢过程(如甲烷产生与排放)具有显著影响[18, 27].对自然湿地和农田生态系统的诸多研究表明, 排水活动引起的系统水位下降能显著减少土壤向大气库中释放CH4[15~19].这是因为排水活动或其它因素引起的系统水位下降可有效增加大气环境中氧气扩散渗透至土壤, 引起土壤从一个厌氧向好氧环境转变, 进而导致甲烷产生菌活性降低, 最终造成较低的CH4生成[28, 29].然而, 本研究发现, 排水后的养殖塘在日观测尺度上向大气库释放CH4强度均显著高于持续淹水的养殖塘, 表明排水活动引起水位降低可显著提高养殖塘的CH4释放能力.分析其原因, 主要跟以下两个方面的综合作用有关:一是, 养殖塘水体虽被排出, 水位发生了急剧降低, 但此时为排水初期, 池塘底部还滞留约0.15 m深的水体, 渗透扩散到沉积物中的氧气有限[图 6(d)], 并且仅有的氧气对表层沉积物影响较大, 而池塘深处的沉积物还处在一个低氧环境, 对整个沉积物的CH4产生和排放贡献较大; 二是, 已有研究表明, CH4在从沉积物经过水体向大气库传输过程中, 因水体溶解氧的存在而时常被氧化, 并且CH4被氧化程度随水体深度增加而提高[12, 30, 31].就本研究而言, 实验期间未排干池塘由于保持着增氧活动, 其水体溶氧水平呈现出显著高于排干池塘的特征[(11.34±0.75) mg·L-1与(6.16±0.44) mg·L-1, 均值是基于白天的观测值计算获得, 夜间数据未采用是因为溶氧仪在进行排干池塘测定时未能稳定读数], 且未排干池塘的平均水深为1.35 m显著高于排干池塘水深的0.15 m.因此, 引起排水后的养殖塘释放CH4强度显著高于持续淹水养殖塘, 也跟排水池塘水体中较低的溶解氧水平以及较低水位(水深0.15 m)使得CH4由沉积物向大气的传输过程中经过水体的路径缩短, 被水体氧化的机会和程度降低有关[30, 32, 33].
硝化和反硝化是土壤(或沉积物)中N2O生成的两个主要的微生物过程, 其N2O产生能力与排放强度也因不同水文条件下具体微生物过程不同而呈现显著差异[34~36].诸多研究发现, 在持续淹水或水位较高环境中, 沉积物中因较低的含氧量和丰富的碳、氮营养物质使得反硝化作用成为N2O产生的主要来源, 保持着相对较低的N2O排放量[18, 27, 34, 36, 37]; 然而, 在水位较低或含水量不饱和的土壤环境中, 土壤因时常呈好气状态, 硝化过程是其N2O产生的主要来源, 拥有相对较高的N2O排放量[18, 35, 38, 39].本研究中, 未排干与排干养殖塘在日变化观测尺度上的N2O通量均值分别为(0.002±0.004) mg·(m2·h)-1和(1.07±0.15) mg·(m2·h)-1, 后者高出前者约500倍, 表明排水活动引起养殖塘水位降低亦可显著提高N2O的释放, 这与上述研究结果基本一致.就本研究而言, 排水活动之所以会显著提高养殖塘N2O释放, 其很大程度上可能跟N2O生成的两个主要微生物过程的共同作用有关.一方面, 研究期间的排干池塘水深介于10~15 cm, 相比未排干池塘, 其较低水深环境有利于大气中部分氧气渗透至池塘表层沉积物中, 加上表层沉积物中残留着大量生物残体、虾排泄物和饵料残体, 可促使沉积物表层微生物参与下的硝化作用产N2O进行.另一方面, 正如前文所述, 排干池塘残留的部分水体可阻止大气中氧气向沉积物深层渗透, 进而为深层沉积物反硝化作用产N2O进行提供了适宜低氧环境.
从日变化特征来看, 排干与未排干养殖塘CH4通量随时间变化整体趋势基本一致(图 2), 这一结果表明, 排水活动引起水位降低对养殖塘CH4通量日变化动态的改变影响较小, 但对养殖塘释放通量大小的改变影响较大.对N2O而言, 本研究发现未排干与排干养殖塘N2O通量随时间变化的趋势在白天趋同, 但在夜间呈现相反的变化趋势(图 3), 表明排水活动引起水位降低不仅可以改变养殖塘N2O通量释放强度, 而且也会对养殖塘N2O通量夜间变化趋势产生影响.
3.2 河口区养殖塘初冬时期温室气体通量日变化特征及主要影响因素研究期间, 未排干和排干养殖塘CH4释放通量随时间推移虽均呈现较大波动, 但整体上均表现为昼低夜高的特征(图 2), 这与相关研究得到的结果相似[9, 40].本研究中, 未排干养殖塘CH4的释放值夜高昼低, 一方面是由于夜间水体浮游藻类的光合作用消失, 池塘鱼虾及微生物进行呼吸作用时会消耗水体中大量氧气[40, 41], 有利于降低CH4在水体扩散传输过程中被氧化消耗的量; 另一方面, 由于养殖塘平均水深仅有1.2 m, 水深较浅, 白天相对强烈的太阳辐射会引起养殖塘表层水温略高于底层水温的稳定热力层结状态, 不利于池塘CH4向大气环境扩散, 而在夜间水体辐射冷却引起水体呈现表层水温略低于底层水温的不稳定热力层结情况, 上下水层较容易混合, 有利于池塘CH4向大气环境扩散[40, 42].此外, 排干养殖塘水-气界面CH4通量与风速的显著正相关关系表明, 夜间相对较高的风速条件也是引起其CH4通量高于白天的一个重要原因.这是由于夜间较高的风速环境有利于上下层水体得到充分交换[9], 有利于降低水-气界面的气体扩散阻力, 进而有利于提高水体中CH4向外扩散[43, 44].对于排干养殖塘而言, 由于本研究中只测定了土温、pH值、电导率EC和Eh等4个理化指标, 且这些指标与CH4通量的相关关系均较弱, 所以引起其CH4释放值夜高昼低的具体原因尚不清楚.但通过对氧化还原电位Eh分析发现[图 6(d)], 养殖塘夜间沉积物的厌氧条件较白天高, 该结果表明夜间沉积物环境维持相对较高的CH4产生能力是引起其CH4释放通量较白天高的一个潜在影响因素.
未排干和排干养殖塘N2O释放通量随时间推移亦均呈现较大波动, 整体上分别表现为昼低夜高和昼高夜低的特征(图 3).未排干养殖塘N2O释放通量呈现上述变化特征, 分析其原因, 很大程度上亦是由于夜间养殖塘水体在较大风力扰动影响下和水体辐射冷却作用下, 上下层水体得到充分交换, 水-气界面的气体扩散阻力降低而引起.对于排干养殖塘而言, 本研究发现, 其N2O释放通量变化轨迹与全天气温、沉积物温度的变化轨迹基本一致[图 3、图 4(a)和图 6(a)], 且N2O释放通量与温度亦呈现出显著正相关系, 表明温度是引起排干养殖塘N2O通量呈现上述日动态的重要因素.类似研究结果也呈现在对湿地[31, 45]和农田[46]的研究中, 并且认为其作用机制跟沉积物中参与硝化-反硝化作用的微生物活性对不同环境温度的响应存在差异性有关.此外, 本研究还发现, 养殖塘表层10 cm深处沉积物氧化还原电位Eh在观测期间呈现白天高于夜间[图 6(a)], 且与N2O释放通量亦具有显著正相关关系(表 2), 表明白天养殖塘表层沉积物中相对较弱的厌氧环境所引起的相对较高的硝化作用是造成其通量高于夜间的另一个重要原因.
4 结论(1) 从释放强度角度来看, 排干养殖塘在各个观测时刻释放CH4和N2O通量强度均极显著高于未排干养殖塘(P<0.01), 表明排干初期的排水活动可显著增强养殖塘释放CH4和N2O的能力.
(2) 从时间变化特征来看, 未排干与排干养殖塘CH4和N2O释放通量随时间推移在白天表现出基本一致的变化趋势, 但在夜间两者的时间变化趋势显著不同; 整体上, 未排干和排干养殖塘CH4释放通量均表现为昼低夜高的特征, 而N2O释放通量分别表现为昼低夜高和昼高夜低的特征.
致谢: 本研究在样品分析过程中得到福建师范大学湿润亚热带生态-地理过程省部共建教育部重点实验室的杨柳明、彭园珍老师的帮助, 在此表示感谢.| [1] | PICC. Climate Change 2013:the Physical Science Basis. Contribution of Working Group i to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[R]. Cambridge, United Kingdom and New York, NY, USA:Cambridge University Press, 2013. |
| [2] |
李建鸿, 蒲俊兵, 孙平安, 等. 不同地质背景水库区夏季水-气界面温室气体交换通量研究[J]. 环境科学, 2015, 36(11): 4032-4042. Li J H, Pu J B, Sun P A, et al. Summer greenhouse gases exchange flux across water-air interface in three water reservoirs located in different geologic setting in Guangxi, China[J]. Environmental Science, 2015, 36(11): 4032-4042. |
| [3] | Kayranli B, Scholz M, Mustafa A, et al. Carbon storage and fluxes within freshwater wetlands:a critical review[J]. Wetlands, 2010, 30(1): 111-124. DOI:10.1007/s13157-009-0003-4 |
| [4] | Song H L, Liu X T. Anthropogenic effects on fluxes of ecosystem respiration and methane in the Yellow River Estuary, China[J]. Wetlands, 2016, 36(S1): 113-123. DOI:10.1007/s13157-014-0587-1 |
| [5] | Tranvik L J, Downing J A, Cotner J B, et al. Lakes and reservoirs as regulators of carbon cycling and climate[J]. Limnology and Oceanography, 2009, 54(6): 2298-2314. |
| [6] | Bastviken D, Tranvik L J, Downing J A, et al. Freshwater methane emissions offset the continental carbon sink[J]. Science, 2011, 331(6013): 50. DOI:10.1126/science.1196808 |
| [7] | Holgerson M A, Raymond P A. Large contribution to inland water CO2 and CH4 emissions from very small ponds[J]. Nature Geoscience, 2016, 9(3): 222-226. DOI:10.1038/ngeo2654 |
| [8] | Holgerson M A. Drivers of carbon dioxide and methane supersaturation in small, temporary ponds[J]. Biogeochemistry, 2015, 124(1-3): 305-318. DOI:10.1007/s10533-015-0099-y |
| [9] |
赵佳玉, 张弥, 肖薇, 等. 基于光谱分析仪的通量-梯度法测量小型池塘水-气界面温室气体交换通量[J]. 环境科学, 2017, 38(1): 41-51. Zhao J Y, Zhang M, Xiao W, et al. Greenhouse gas fluxes at water-air interface in small pond using flux-gradient method based on spectrum analyzer[J]. Environmental Science, 2017, 38(1): 41-51. |
| [10] | Yang P, He Q H, Huang J F, et al. Fluxes of greenhouse gases at two different aquaculture ponds in the coastal zone of southeastern China[J]. Atmospheric Environment, 2015, 115: 269-277. DOI:10.1016/j.atmosenv.2015.05.067 |
| [11] | Martinez-Cruz K, Gonzalez-Valencia R, Sepulveda-Jauregui A, et al. Methane emission from aquatic ecosystems of Mexico City[J]. Aquatic Sciences, 2017, 79(1): 159-169. DOI:10.1007/s00027-016-0487-y |
| [12] |
龙丽, 肖尚斌, 张成, 等. 亚热带浅水池塘水-气界面甲烷通量特征[J]. 环境科学, 2016, 37(12): 4552-4559. Long L, Xiao S B, Zhang C, et al. Characteristics of methane flux across the water-air interface in subtropical shallow ponds[J]. Environmental Science, 2016, 37(12): 4552-4559. |
| [13] | Verdegem M C J, Bosma R H. Water withdrawal for brackish and inland aquaculture, and options to produce more fish in ponds with present water use[J]. Water Policy, 2009, 11(S1): 52-68. DOI:10.2166/wp.2009.003 |
| [14] | 李生, 朱旺明, 周萌. 南美白对虾高效养成新技术与实例[M]. 北京: 海洋出版社, 2013. |
| [15] | MacDonald J A, Fowler D, Hargreaves K J, et al. Methane emission rates from a northern wetland; response to temperature, water table and transport[J]. Atmospheric Environment, 1998, 32(19): 3219-3227. DOI:10.1016/S1352-2310(97)00464-0 |
| [16] | Kang G D, Cai Z C, Feng X Z. Importance of water regime during the non-rice growing period in winter in regional variation of CH4 emissions from rice fields during following rice growing period in China[J]. Nutrient Cycling in Agroecosystems, 2002, 64(1-2): 95-100. |
| [17] | Blodau C, Moore T R. Experimental response of peatland carbon dynamics to a water table fluctuation[J]. Aquatic Sciences, 2003, 65(1): 47-62. DOI:10.1007/s000270300004 |
| [18] | Dinsmore K J, Skiba U M, Billett M F, et al. Effect of water table on greenhouse gas emissions from peatland mesocosms[J]. Plant and Soil, 2009, 318(1-2): 229-242. DOI:10.1007/s11104-008-9832-9 |
| [19] |
张怡, 吕世华, 马静, 等. 冬季水分管理和水稻覆膜栽培对川中丘陵地区冬水田CH4排放的影响[J]. 生态学报, 2016, 36(4): 1095-1103. Zhang Y, Lü S H, Ma J, et al. Effects of water management in winter and of plastic film mulching during rice cultivation on CH4 emission from paddy field in a hilly region of Central Sichuan[J]. Acta Ecologica Sinica, 2016, 36(4): 1095-1103. |
| [20] |
郑彩虹, 曾从盛, 陈志强, 等. 闽江河口区湿地景观格局演变研究[J]. 湿地科学, 2006, 4(1): 29-35. Zheng C H, Zeng C S, Chen Z Q, et al. A study on the changes of landscape pattern of estuary wetlands of the Minjiang River[J]. Wetland Science, 2006, 4(1): 29-35. |
| [21] | Zhang L, Wang L, Yin K D, et al. Pore water nutrient characteristics and the fluxes across the sediment in the Pearl River estuary and adjacent waters, China[J]. Estuarine, Coastal and Shelf Science, 2013, 133: 182-192. DOI:10.1016/j.ecss.2013.08.028 |
| [22] |
杨平, 金宝石, 谭立山, 等. 九龙江河口区养虾塘沉积物-水界面营养盐交换通量特征[J]. 生态学报, 2017, 37(1): 192-203. Yang P, Jin B S, Tan L S, et al. Temporal variation of nutrients fluxes across the sediment-water interface of shrimp ponds and influencing factors in the Jiulong River Estuary[J]. Acta Ecologica Sinica, 2017, 37(1): 192-203. |
| [23] | Lorenzen C J. Determination of chlorophyll and pheo-pigments:spectrophotometric equations[J]. Limnology and Oceanography, 1967, 12(2): 343-346. DOI:10.4319/lo.1967.12.2.0343 |
| [24] | Jeffrey S W, Humphrey G F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton[J]. Biochemie und Physiologie der Pflanzen, 1975, 167(2): 191-194. DOI:10.1016/S0015-3796(17)30778-3 |
| [25] |
张莹, 王玉珏, 王跃启, 等. 2013年夏季渤海环境因子与叶绿素a的空间分布特征及相关性分析[J]. 海洋通报, 2016, 35(5): 571-578. Zhang Y, Wang Y J, Wang Y Q, et al. Spatial distribution and correlation of environmental factors and chlorophyll a concentrations in the Bohai Sea during the summer of 2013[J]. Marine Science Bulletin, 2016, 35(5): 571-578. DOI:10.11840/j.issn.1001-6392.2016.05.011 |
| [26] |
张坚超, 徐镱钦, 陆雅海. 陆地生态系统甲烷产生和氧化过程的微生物机理[J]. 生态学报, 2015, 35(20): 6592-6603. Zhang J C, Xu Y Q, Lu Y H. Microbial mechanisms of methane production and oxidation in terrestrial ecosystems[J]. Acta Ecologica Sinica, 2015, 35(20): 6592-6603. |
| [27] | Yang J S, Liu J S, Hu X J, et al. Effect of water table level on CO2, CH4 and N2O emissions in a freshwater marsh of Northeast China[J]. Soil Biology and Biochemistry, 2013, 61: 52-60. DOI:10.1016/j.soilbio.2013.02.009 |
| [28] | Lombardi J E, Epp M A, Chanton J P. Investigation of the methyl fluoride technique for determining rhizospheric methane oxidation[J]. Biogeochemistry, 1997, 36(2): 153-172. DOI:10.1023/A:1005750201264 |
| [29] | Strack M, Waddington J M, Tuittila E S. Effect of water table drawdown on northern peatland methane dynamics:implications for climate change[J]. Global Biogeochemical Cycles, 2004, 18(4): GB4003. |
| [30] | Bastviken D, Cole J J, Pace M L, et al. Fates of methane from different lake habitats:connecting whole-lake budgets and CH4 emissions[J]. Journal of Geophysical Research:Biogeosciences, 2008, 113(G2): G02024. |
| [31] |
盛宣才, 吴明, 邵学新, 等. 模拟水位变化对杭州湾芦苇湿地夏季温室气体日通量的影响[J]. 生态学报, 2016, 36(15): 4792-4800. Sheng X C, Wu M, Shao X X, et al. Effects of simulated water levels on diurnal variation in the emission of three greenhouse gases in reed wetlands in summer[J]. Acta Ecologica Sinica, 2016, 36(15): 4792-4800. |
| [32] | Schütz H, Seiler W, Conrad R. Processes involved in formation and emission of methane in rice paddies[J]. Biogeochemistry, 1989, 7(1): 33-53. |
| [33] | Hu Z Q, Wu S, Ji C, et al. A comparison of methane emissions following rice paddies conversion to crab-fish farming wetlands in southeast China[J]. Environmental Science and Pollution Research, 2016, 23(2): 1505-1515. DOI:10.1007/s11356-015-5383-9 |
| [34] | Martikainen P J, Nykänen H, Crill P, et al. Effect of a lowered water table on nitrous oxide fluxes from northern peatlands[J]. Nature, 1993, 366: 51-53. DOI:10.1038/366051a0 |
| [35] | Xu H, Xing G X, Cai Z C, et al. Nitrous oxide emissions from three rice paddy fields in China[J]. Nutrient Cycling in Agroecosystems, 1997, 49(1-3): 23-28. |
| [36] |
李英臣, 宋长春, 刘德燕. 湿地土壤N2O排放研究进展[J]. 湿地科学, 2008, 6(2): 124-129. Li Y C, Song C C, Liu D Y. Advances in studies of N2O emission in wetland soils[J]. Wetland Science, 2008, 6(2): 124-129. |
| [37] | Kudo Y, Noborio K, Shimoozono N, et al. The effective water management practice for mitigating greenhouse gas emissions and maintaining rice yield in central Japan[J]. Agriculture, Ecosystems & Environment, 2014, 186: 77-85. |
| [38] |
李香兰, 徐华, 蔡祖聪. 水分管理影响稻田氧化亚氮排放研究进展[J]. 土壤, 2009, 41(1): 1-7. Li X L, Xu H, Cai Z C. Effect of water management on nitrous oxide emission from rice paddy field:a review[J]. Soils, 2009, 41(1): 1-7. |
| [39] | Mander V, Maddison M, Soosaar K, et al. The Impact of pulsing hydrology and fluctuating water table on greenhouse gas emissions from constructed wetlands[J]. Wetlands, 2011, 31(6): 1023-1032. DOI:10.1007/s13157-011-0218-z |
| [40] |
汪国骏, 胡明明, 王雨春, 等. 蓄水初期三峡水库草堂河水-气界面CO2和CH4通量日变化特征及其影响因素[J]. 湖泊科学, 2017, 29(3): 696-704. Wang G J, Hu M M, Wang Y C, et al. Diurnal variation and influencing factors of carbon dioxide and methane emissions at water-air interface of Caotang River, Three Gorges Reservoir in the initial impoundment period[J]. Journal of Lake Sciences, 2017, 29(3): 696-704. DOI:10.18307/2017.0319 |
| [41] |
杨平, 仝川, 何清华, 等. 闽江口养殖塘水-大气界面温室气体通量日进程特征[J]. 环境科学, 2012, 33(12): 4194-4204. Yang P, Tong C, He Q H, et al. Diurnal variations of greenhouse gas fluxes at the water-air interface of aquaculture ponds in the Min River estuary[J]. Environmental Science, 2012, 33(12): 4194-4204. |
| [42] | Bartlett K B, Crill P M, Bonassi J A, et al. Methane flux from the Amazon River floodplain:emissions during rising water[J]. Journal of Geophysical Research:Atmospheres, 1990, 95(D10): 16773-16788. DOI:10.1029/JD095iD10p16773 |
| [43] |
杨平, 仝川. 淡水水生生态系统温室气体排放的主要途径及影响因素研究进展[J]. 生态学报, 2015, 35(20): 6868-6880. Yang P, Tong C. Emission paths and measurement methods for greenhouse gas fluxes from freshwater ecosystems:a review[J]. Acta Ecologica Sinica, 2015, 35(20): 6868-6880. |
| [44] |
黄文敏, 朱孔贤, 赵玮, 等. 香溪河秋季水-气界面温室气体通量日变化观测及影响因素分析[J]. 环境科学, 2013, 34(4): 1270-1276. Huang W M, Zhu K X, Zhao W, et al. Diurnal changes in greenhouse gases at water-air interface of Xiangxi river in autumn and their influencing factors[J]. Environmental Science, 2013, 34(4): 1270-1276. |
| [45] |
卢妍, 宋长春, 徐洪文. 沼泽湿地草甸N2O排放通量日变化规律研究[J]. 上海环境科学, 2009, 28(4): 139-142, 148. Lu Y, Song C C, Xu H W. Studies of diurnal variation of nitrous oxide emission fluxes from marsh wetland meadow[J]. Shanghai Environmental Sciences, 2009, 28(4): 139-142, 148. |
| [46] |
谢军飞, 李玉娥. 土壤温度对北京旱地农田N2O排放的影响[J]. 中国农业气象, 2005, 26(1): 7-10. Xie J F, Li Y E. Effect of soil temperature on N2O emission in upland farm of Beijing[J]. Chinese Journal of Agrometeorology, 2005, 26(1): 7-10. |