 2017, Vol. 38
2017, Vol. 38



2. 中国科学院山地表生与生态调控重点实验室, 成都 610041;
3. 中国科学院大学, 北京 100049
2. Key Laboratory of Mountain Surface Processes and Ecological Regulation, Chinese Academy of Sciences, Chengdu 610041, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
三峡水库采用“蓄清排浊”的水库运行调度机制, 水库从9月底开始蓄水至次年1月(水位高程175 m)逐渐下降, 6月水位高程回落至最低145 m.从而在库区两岸形成了垂直落差30 m, 面积为349 km2的消落带[1].消落带平均淹水期大于6个月, 最长约240 d(145 m)[1].消落带夏季出露, 植物生长旺盛, 9月植物生物量最大时蓄水淹没, 导致植物腐解的同时释放出大量养分, 可能影响库区水环境[2~4].三峡库区消落带在经历数次淹水和落干后, 植物群落多样性发生变化, 物种的结构和组成趋于简单化[5, 6].狗牙根[Cynodon dactylon (Linn.) Pers.]不仅在库区消落带分布范围广, 且耐淹性强, 是自然和人工恢复的主要多年生草本植物[6, 7].但笔者前期的研究发现, 狗牙根随消落带蓄水淹没后, 仍然有矿化分解, 并且养分释放亦较为可观[2, 4].
目前已有研究主要是针对植物本身初始基质含量(如C、N、P、木质素、纤维素含量等)对植物的分解速率和养分释放的影响[3, 4, 8], 对消落带植物在淹水分解中受环境因素影响的研究较少.植物在浸泡腐烂过程的物理、化学以及微生物过程可能会受到温度、pH、光照等因素的影响[8~12].温度是控制水生植物腐解的重要环境因子, 有研究指出夏季水温升高, 使植物的分解速率要明显大于冬季[8].三峡库区水温年变化范围为11~28℃[13], 温度可能通过改变微生物的活性等来影响植物的淹水腐解过程[8].同时, 水体的pH值也会对植物的分解速率产生影响[9, 10], 三峡库区水体的来源范围广泛, 有工业废水、城镇污水、农业排污等, 导致库区水体pH值的时空分布差异性大, 江水pH范围为6~9, 主要偏碱性[14].此外, 最底层消落带淹水时垂直深度最高可达30m, 此时的植物和土壤几乎处于避光状态[15].
综上所述, 消落带植物在淹水时上覆水体的环境条件(水温、pH、光照条件)差异较大, 但水环境条件的变化会对消落带植物淹水分解时养分的释放产生怎样的影响目前还未见报道.因此, 本文针对三峡水库水环境变化的实际情况, 模拟狗牙根浸泡时上覆水温度、pH和光照等条件变化, 探讨三峡水环境变化对消落带优势草本植物——狗牙根氮磷养分释放的影响, 以期为三峡库区植被对干湿交替的生态响应、环境影响和消落带植被恢复重建提供科学依据.
1 材料与方法 1.1 研究区概况三峡库区主要涉及湖北省宜昌、秭归、兴山、巴东和重庆市巫山、巫溪、奉节、云阳、万州、开县、忠县、丰都、石柱、涪陵、武隆、长寿、渝北、巴南、重庆市区和江津市共20个县(市、区).库区范围为东经105°44′~111°39′, 北纬28°32′~31°44′, 总面积5.67万km2[1].本研究所采用的狗牙根均采集自重庆市忠县石宝镇共和村消落带160 m高程处(30°24′~30°52′N, 107°23′~108°09′E), 属于亚热带季风气候, 年降雨量1 000~1 800 mm, 日照充足, 年平均气温18.2℃, 相对湿度80%.
1.2 实验设计多年生草本禾本科植物狗牙根[Cynodon dactylon (Linn.) Pers.]为消落带人工恢复植被的重要物种, 已在三峡库区消落带广泛分布[4, 6].选取狗牙根进行淹水浸泡模拟实验.根据三峡库区的水位调节规律, 在9月水库开始准备蓄水前的160~170 m水位高程处采集本实验所用的狗牙根样品.将采集的狗牙根鲜样(地上部分)混合均匀后取50 g入尼龙网袋内(20 cm×20 cm, 网孔大小为1.0 mm×1.0 mm), 放入盛有2 L去离子水(水温已在培养箱中培养至25℃恒温)的玻璃烧杯中, 尼龙网袋完全淹没在水中, 透光浸泡、水体pH为7.0, 系统为敞口开放环境, 以模拟三峡水库的实际.每间隔1 d, 用pH计测定浸泡上覆水酸碱度, 并用0.1 mol·L-1的氢氧化钠和0.05 mol·L-1的硫酸将上覆水的pH维持为7.0.
设置25℃和15℃两种不同的水温环境, 先将水温在培养箱中调整至目标值并保持恒定后加入狗牙根样品进行浸泡.在pH为7.0的透光开放系统中进行不同水温条件下狗牙根浸泡的模拟.
在25℃透光条件下, 另设置3组不同水体pH环境下狗牙根的模拟浸泡.在去离子水基础上, 使用0.1 mol·L-1的氢氧化钠和0.05 mol·L-1的硫酸将上覆水调节至pH值为5.0、7.0、9.0, 每间隔1 d, 用pH计测定浸泡上覆水酸碱度, 并用0.1 mol·L-1的氢氧化钠和0.05 mol·L-1的硫酸维持上覆水的pH至目标值.
为了了解光照与否对狗牙根浸泡的影响, 在上覆水水温为25℃、pH为7.0的基础上, 分别设置透光和避光两种光照条件.每组实验设置3个平行和一个空白对照.
在实验进行的第1、2、3、5、7、10、15、20、30、45、60、90、120、150、200 d(以消落带160 m高程处最长淹水时间为例[4]), 取不同淹水条件下的上覆水10 mL, 经0.45 μm微孔滤膜过滤后用AA3流动注射分析仪(AA3, SEAL Analytical GmbH, 德国)测定养分形态及含量[总氮(TN)、可溶性总氮(DN)、硝态氮(NO3--N)、总磷(TP)、可溶性总磷(DP)及正磷酸盐磷(PO43--P)], 颗粒态氮(PN)含量为总氮减去可溶态氮, 颗粒态磷(PP)含量为总磷减去可溶态磷.取样后补充相应水分, 以保证实验过程中浸泡水体体积的一致.同时每30 d全面换水一次以模拟水库换水周期且避免上覆水养分饱和.
在浸泡前后分别测定狗牙根的干重.植物的全氮、全磷含量是在植物体烘干磨碎后进行测定, 方法如下[16]:用浓硫酸消煮后, 凯氏定氮仪蒸馏法(加氢氧化钠碱化)测定植物全氮; 用硝酸-高氯酸混合液消煮后用钼锑抗比色法测定植物全磷.
1.3 数据计算及处理狗牙根植物体在浸泡后总的氮(磷)损失量为浸泡分解前后氮磷元素的含量和干重乘积的差值, 植物残体养分损失量的计算公式如下:
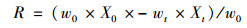
|
(1) |
式中, R为狗牙根干重氮(磷)的损失量(g·kg-1); wt为浸泡200 d后狗牙根的干重(g), Xt为200 d后狗牙根中氮(磷)元素的含量(g·kg-1), w0为浸泡前狗牙根的初始干重(g), X0为浸泡前狗牙根中氮(磷)元素的初始含量(g·kg-1).
狗牙根在浸泡过程中氮(磷)的累积释放量则为上覆水中累积养分浓度与上覆水体积的乘积再比上植物浸泡的初始干重值, 计算如下:
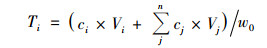
|
(2) |
式中, Ti为养分的累积释放量(g·kg-1); ci、Vi分别为第i次取样时溶液的养分浓度(mg·L-1)和体积(mL); n为换水次数; cj、Vj分别为第j次换水时溶液的养分浓度(mg·L-1)和换水体积(mL).
实验数据使用Excel 2007进行处理后, 采用One way-ANOVA方法对数据进行显著性分析.养分释放量与残体性质的相关性使用Pearson相关分析进行.数据的统计分析使用SPSS 16.0软件, 图形的处理使用Origin 8.1软件.
2 结果与分析 2.1 狗牙根浸泡前后的干重与养分变化测定狗牙根在浸泡前后的干重、全氮和全磷含量, 结果见表 1.狗牙根初始基质全氮含量为(6.92±0.10) g·kg-1, 全磷含量为(1.16±0.03) g·kg-1.经过200 d的模拟浸泡狗牙根干重减少至(15.01±0.91) g, 干重损失率为64%.狗牙根残体的全氮、全磷含量分别为(6.91±0.19) g·kg-1、(0.30±0.02) g·kg-1.
|
|
表 1 狗牙根在淹水浸泡前后的残体干重和氮磷养分含量 Table 1 Dry weight of plant detritus and the nitrogen and phosphorus content of Cynodondactylon (Linn.) Pers. before and after inundation |
2.2 水温对狗牙根养分浸泡释放的影响
狗牙根在25℃和15℃水温下浸泡时上覆水TN、NO3--N、PN、TP、PO43--P和PP的浓度变化如图 1所示.可以看出, 不同温度浸泡下的上覆水各养分浓度在30 d内没有明显的差异, 30 d后温度的变化开始影响养分的浓度.浸泡30 d后, 25℃时上覆水NO3--N和PO43--P浓度均大于15℃, 而TP和PP浓度一直小于15℃时.上覆水中TN和PN浓度在浸泡30~60 d内, 25℃时大于15℃, 60 d后15℃大于25℃.整个浸泡过程中, 上覆水PN的浓度变化与TN变化一致, 至上覆水各养分浓度稳定后(120 d), 25℃浸泡时PN浓度占TN浓度的80%(质量分数)以上, 15℃时PN占TN的90%以上. 120 d后, 25℃和15℃时PP占TP分别为26.5%和57.2%.

|
图 1 狗牙根在不同水温浸泡下的上覆水TN、NO3--N、PN、TP、PO43--P和PP变化 Fig. 1 Variations of TN, NO3--N, PN, TP, PO43--P and PP concentrations in the overlying water when Cynodon dactylon (Linn.) Pers. inundated at different temperatures |
狗牙根浸泡200 d后, 25℃和15℃水温条件下狗牙根TN和TP的释放量对比见图 2. 25℃和15℃条件下上覆水中TN的释放量分别为(2.66±0.29) g·kg-1和(3.76±0.04) g·kg-1; 上覆水中TP的释放量分别为(0.79±0.03) g·kg-1和(1.40±0.02) g·kg-1.

|
图 2 狗牙根在不同温度浸泡200 d后的氮、磷释放量 Fig. 2 Amounts of nitrogen and phosphorus released into overlying water from Cynodon dactylon (Linn.) Pers. after 200 d of inundation at different temperatures |
图 3显示了不同上覆水pH条件下狗牙根浸泡时上覆水的TN、NO3--N、PN、TP、PO43--P和PP浓度变化.上覆水TN和PN浓度变化一致, 且在30 d后pH 5.0浸泡条件下浓度明显高于pH 7.0和pH 9.0时, 碱性和中性条件差异不大.至PN浓度稳定后, 不同pH条件下PN占TN的85%以上.浸泡60 d后上覆水TP和PP浓度快速上升, 浓度随上覆水pH值变化顺序为pH 9.0 > pH 5.0 > pH 7.0.在碱性条件下上覆水中磷以PP为主, 浓度稳定后PP占比达75.5%;酸性和中性条件下, PP占比在40%以下.上覆水NO3--N和PO43--P浓度变化一致, 均在上覆水中性时浓度最大, 碱性条件时最小.

|
图 3 狗牙根在不同pH水体浸泡时上覆水的TN、NO3--N、PN、TP、PO43--P和PP变化 Fig. 3 Variations of TN, NO3--N, PN, TP, PO43--P and PP concentrations in overlying water when Cynodon dactylon (Linn.)Pers. inundated at different pH levels |
不同pH浸泡200 d后TN和TP的释放量如图 4所示. pH 5.0、pH 7.0和pH 9.0溶液下浸泡, 狗牙根向上覆水中释放的TN分别为(3.76±0.08)、(2.66±0.29) 和(2.55±0.12) g·kg-1, TN释放量具有显著差异(P < 0.05); TP的释放量分别为(1.53±0.04)、(0.79±0.03) 和(1.70±0.07) g·kg-1, 具有显著差异(P < 0.05).

|
图 4 狗牙根在不同pH水体浸泡200 d后的氮、磷释放量 Fig. 4 Amounts of nitrogen and phosphorus released into overlying water from Cynodon dactylon (Linn.) Pers. after 200 d of inundation at different pH levels |
狗牙根在透光和避光条件下浸泡时上覆水TN、NO3--N、PN、TP、PO43--P和PP的浓度变化过程见图 5.当上覆水各养分浓度达到平衡后(120 d), 避光条件下上覆水TN、TP、PO43--P和PP的浓度均大于透光浸泡时, 上覆水NO3--N浓度小于透光时.上覆水PN浓度变化不受光照的影响.避光浸泡过程中, 氮、磷的释放均以颗粒态为主, PN占81.8%, PP占42.8%.

|
图 5 狗牙根在不同光照水体浸泡下上覆水的TN、NO3--N、PN、TP、PO43--P和PP变化 Fig. 5 Variations of TN, NO3--N, PN, TP, PO43--P and PP concentrations in the overlying water when Cynodon dactylon (Linn.) Pers. inundated under light or dark conditions |
狗牙根避光浸泡200 d后TN和TP的释放量明显大于透光时(图 6), 避光时上覆水TN和TP的释放量为(3.87±0.14) g·kg-1和(1.78±0.08) g·kg-1.

|
图 6 狗牙根在光照与否条件浸泡200 d后氮、磷的释放量 Fig. 6 Amounts of nitrogen and phosphorus released into overlying water from Cynodon dactylon (Linn.) Pers. after 200 d inundation under light or dark conditions |
Xiao等[4]研究已经表明了三峡消落带典型植物的浸泡分解具有明显的阶段性, 前期主要是受到初始基质控制下的易溶物质快速溶解过程, 而后则受控于环境条件控制的微生物分解作用[11, 17], 且在120 d内浸泡分解基本完成.植物的分解速率主要受到植物初始基质性质控制, 氮、C/N、木质素等都被认为是主要的影响因素[17].在本研究中, 浸泡时间的推移使得环境因素开始逐步对狗牙根的分解产生影响.浸泡分解结束后, 狗牙根残体中全氮含量的减少幅度明显小于全磷含量的减少, 说明狗牙根在淹水分解时氮磷组分损失的受控过程不同[4].植物分解时, 氮的损失不仅增加了水体的氮负荷, 更会受到微生物作用变为含氮气体进入大气[11].磷主要以磷酸根的形式在植物体内存在, 容易通过淋溶作用流失[17].
3.2 上覆水环境条件改变对狗牙根氮、磷释放的影响在各水环境条件下狗牙根浸泡200 d后, 残体氮、磷的损失量(表 2)与上覆水中TN、TP的释放量存在差异且没有明显的相关性(P > 0.05).这说明了植物浸泡时对水体的营养元素贡献值不能通过植物氮磷的损失来进行估算, 因为在分解过程中, 植物体内的氮、磷不仅能释放到水中, 也可能被微生物吸收或者以气态形式挥发, 而水中的颗粒态氮、磷则可能会重新沉淀后附着在植物体上[17].
|
|
表 2 不同条件浸泡200 d后狗牙根残体的氮、磷损失量1) Table 2 Amount of nitrogen and phosphorus lost from the residual in Cynodon dactylon (Linn.) Pers. |
植物分解时向水中释放养分是复杂的物理、化学、生物过程的集合, 受环境因素、植物形态与性质等的制约[12].本实验选用的浸泡植物为狗牙根, 植物内在机制的影响大致相似, 使得植物养分的浸泡释放主要由外界环境所控制.
水温被认为对湿地水生植物分解的影响要大于植物本身的性质, 温度越高, 植物的物理溶解加速和微生物活性增强[8].温度可以通过调节系统微生物的活性来影响植物溶解态养分的释放[18].一般认为, 25℃时微生物活性最强, 氨化、硝化和反硝化作用增强, 利于大分子化合物的溶解及水中的氮转化为N2O进入大气, 这导致了上覆水中NO3--N和PO43--P的浓度升高[19].然而, 本研究中溶液温度越低, 狗牙根上覆水中TN、TP释放量越大.主要是因为水中植物的分解由微生物和腐生生物共同作用, 而Martínez等[20]研究发现, 微生物分解速率不会随水温发生变化.而植物腐烂过程中产生的腐生生物使植物分解为大量的小分子颗粒物, 而这些腐生生物更利于在温度较低的环境中生存[21].这就导致了低温浸泡时, 狗牙根上覆水中PN、PP的释放量增大, 而由于氮、磷的释放主要以PN、PP为主, 使TN、TP的释放量较大.在三峡水库运行时冬季高水位期平均水温为(14.92±4.67)℃, 夏季低水位期时水温为(21.37±4.70)℃[14].同时, 刘流等[22]的研究表明三峡库区冬季蓄水时水温无分层现象, 而在3~5月蓄水时水温会出现明显的分层, 表层水温在14~22℃, 30 m水深处水温仅10~15℃.因此, 库区冬季高水位蓄水时, 大面积植物淹水且水温较低, 可能在其腐烂分解的过程中对水体氮、磷的产生较大影响.
水体pH值不仅能改变某些物质的溶解状态, 也会影响微生物的分解作用, 这两者都关系到植物浸泡时对养分的释放.狗牙根在中性条件淹水分解初期会消耗水体中的O2产生CO2导致pH值有一个快速的降低, 但长期浸泡后对上覆水体pH的影响不大[2].酸性或碱性浸泡条件下, 植物中的木质素、纤维素等结构发生改变, 加速植物组织中养分的溶解, 增加了狗牙根向水体中释放养分的量, 导致酸碱条件下狗牙根养分浸泡释放量大[9].而碱性条件下, 不利于溶液中铝浓度的增加, 从而使磷不易结合成稳定的化合物, 更利于磷的释放[10].三峡库区水体以弱碱性为主[14], 当存在排污口时, 会使水体酸性或碱性增大, 均能加速消落带植物淹水后对氮、磷的释放, 可能会使得三峡库区水体富营养水平加剧.
光照与否主要通过影响系统中微生物的活性及光催化降解过程而改变植物分解和养分释放[23].本研究中狗牙根在避光条件下更易于向上覆水中释放氮磷, 特别是对TP和PP释放的促进作用明显.一般而言, 光照能够为微生物分解者提供充足的能量及营养物质[24], 从而加速微生物对植物的分解和木质素、纤维素的降解[23].本研究的结果与此相反, 因为避光条件下微生物裂解酶的增加会导致植物体中有机物质的光化学聚合作用减弱而催化降解作用增强[25], 从而更利于狗牙根浸泡向水体中释放养分.同时, Bosco等[12]认为光照虽然能促进植物分解时干重的下降, 但不能直接影响到植物体内木质素等物质的化学分解.在浸泡初始时狗牙根中大量的有机物质会通过物理作用洗脱至上覆水中, 光照条件下这些可溶性有机物更易于被光催化降解而不是被微生物所利用, 使水中可以提供给微生物生长的营养元素减少[26], 从而抑制了养分的释放.由于三峡库区水位高程较低处的植物受水淹胁迫, 淹水时间长且水深高水压大, 库区水位到达最高175 m时, 最底层的植物和土壤光照弱几乎处于避光状态[15].因此, 综合消落带最底层植物淹水时的水温和光照等多方面条件, 认为底层植物的淹水浸泡对库区水体的影响更大.
4 结论(1) 不同淹水条件浸泡时, 消落带优势植物种狗牙根的干重下降, 植物体内氮、磷养分释放.浸泡结束后植物残体的氮、磷损失量与上覆水TN、TP释放量存在差异.
(2) 狗牙根浸泡上覆水氮磷养分的浓度在前30 d几乎不受环境条件的影响. 120 d后植物分解完全, 上覆水中氮磷养分浓度保持稳定, 且颗粒态氮和颗粒态磷在释放过程中占主要地位.不同温度, pH和光照条件下, 狗牙根淹水浸泡200 d后上覆水TN释放量为(2.55±0.12)~(3.87±0.14) g·kg-1, TP释放量为(0.79±0.03)~(1.78±0.08) g·kg-1.
(3) 随三峡水环境的变化, 消落带狗牙根浸泡时养分的释放也有所不同.如库区冬季蓄水水温较低时, 植物浸泡上覆水氮磷的释放量增大; 当存在酸性或碱性污水排放时, 有利于植物的腐解, 会促进养分的释放; 当水位较高时底层植物处于避光条件下, 更有利于植物浸泡向水体中释放养分.因此, 对三峡水环境条件的确定有助于估算消落带植物浸泡过程对库区水体水质的影响.
| [1] | Yuan X Z, Zhang Y W, Liu H, et al. The littoral zone in the Three Gorges Reservoir, China:challenges and opportunities[J]. Environmental Science and Pollution Research, 2013, 20(10): 7092-7102. DOI:10.1007/s11356-012-1404-0 |
| [2] |
王建超, 朱波, 汪涛, 等. 三峡库区典型消落带草本植物氮磷养分浸泡释放实验[J]. 环境科学, 2012, 33(4): 1144-1151. Wang J C, Zhu B, Wang T, et al. Nitrogen and phosphorus release from herbaceous vegetation under simulated inundation experiment of water-level fluctuation zone in the Three Gorges Reservoir area[J]. Environmental Science, 2012, 33(4): 1144-1151. |
| [3] |
袁庆叶, 谢宗强, 杨林森, 等. 水淹条件下三峡水库消落带常见草本植物的分解[J]. 应用生态学报, 2014, 25(8): 2229-2237. Yuan Q Y, Xie Z Q, Yang L S, et al. Decomposition of herbaceous species in reservoir riparian region of Three Gorges Reservoir under flooding condition[J]. Chinese Journal of Applied Ecology, 2014, 25(8): 2229-2237. |
| [4] | Xiao L W, Zhu B, Kumwimba M N, et al. Plant soaking decomposition as well as nitrogen and phosphorous release in the water-level fluctuation zone of the Three Gorges Reservoir[J]. Science of the Total Environment, 2017, 592: 527-534. DOI:10.1016/j.scitotenv.2017.03.104 |
| [5] |
雷波, 王业春, 由永飞, 等. 三峡水库不同间距高程消落带草本植物群落物种多样性与结构特征[J]. 湖泊科学, 2014, 26(4): 600-606. Lei B, Wang Y C, You Y F, et al. Diversity and structure of herbaceous plant community in typical water-level-fluctuation zone with different spacing elevations in Three Gorges Reservoir[J]. Journal of Lake Sciences, 2014, 26(4): 600-606. DOI:10.18307/2014.0415 |
| [6] |
王建超, 朱波, 汪涛. 三峡库区典型消落带淹水后草本植被的自然恢复特征[J]. 长江流域资源与环境, 2011, 20(5): 603-610. Wang J C, Zhu B, Wang T. Characteristics of restoration of natural herbaceous vegetation of typical water-level fluctuation zone after flooding in the Three gorges Reservoir area[J]. Resources and Environment in the Yangtze Basin, 2011, 20(5): 603-610. |
| [7] |
谭淑端, 张守君, 张克荣, 等. 长期深淹对三峡库区三种草本植物的恢复生长及光合特性的影响[J]. 植物科学学报, 2009, 27(4): 391-396. Tan S D, Zhang S J, Zhang K R, et al. Effect of long-time and deep submergence on recovery growth and photosynthesis of three grass species in Three Gorges Reservoir area[J]. Journal of Wuhan Botanical Research, 2009, 27(4): 391-396. |
| [8] | Liu G D, Sun J F, Tian K, et al. Long-term responses of leaf litter decomposition to temperature, litter quality and litter mixing in plateau wetlands[J]. Freshwater Biology, 2016, 62(1): 178-190. |
| [9] | Merrix F L, Lewis B R, Ormerod S J. The effects of low pH and palliative liming on beech litter decomposition in acid-sensitive streams[J]. Hydrobiologia, 2006, 571(1): 373-381. DOI:10.1007/s10750-006-0269-y |
| [10] | Clivot H, Charmasson F, Felten V, et al. Danger, Interactive effects of aluminium and phosphorus on microbial leaf litter processing in acidified streams:a microcosm approach[J]. Environmental Pollution, 2014, 186: 67-74. DOI:10.1016/j.envpol.2013.11.024 |
| [11] | Raposeiro P M, Ferreira V, Guri R, et al. Leaf litter decomposition on insular lentic systems:effects of macroinvertebrate presence, leaf species, and environmental conditions[J]. Hydrobiologia, 2017, 784(1): 65-79. DOI:10.1007/s10750-016-2852-1 |
| [12] | Bosco T, Bertiller M B, Carrera A L. Combined effects of litter features, UV radiation, and soil water on litter decomposition in denuded areas of the arid Patagonian Monte[J]. Plant and Soil, 2016, 406(1-2): 71-82. DOI:10.1007/s11104-016-2864-7 |
| [13] |
任华堂, 陈永灿, 刘昭伟. 三峡水库水温预测研究[J]. 水动力学研究与进展, 2008, 23A(2): 141-148. Ren H T, Chen Y C, Liu Z W. Study on water temperature prediction in Three Gorges Reservoir[J]. Chinese Journal of Hydrodynamics, 2008, 23A(2): 141-148. |
| [14] |
郭胜, 李崇明, 郭劲松, 等. 三峡水库蓄水后不同水位期干流氮、磷时空分异特征[J]. 环境科学, 2011, 32(5): 1266-1272. Guo S, Li C M, Guo J S, et al. Spatio-temporal variation of nitrogen, phosphorus in different period in Three Gorges Reservoir after its impoundment[J]. Environmental Science, 2011, 32(5): 1266-1272. |
| [15] |
施美芬, 曾波, 申建红, 等. 植物水淹适应与碳水化合物的相关性[J]. 植物生态学报, 2010, 34(7): 855-866. Shi M F, Zeng B, Shen J H, et al. A review of the correlation of flooding adaptability and carbohydrates in plants[J]. Chinese Journal of Plant Ecology, 2010, 34(7): 855-866. |
| [16] |
鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. Lu R K. Analytical methods of soil and agricultural chemistry[M]. Beijing: China Agricultural Science and Technology Press, 2000. |
| [17] | Moore R T, Trofymow A J, Prescott E C, et al. Patterns of carbon, nitrogen and phosphorus dynamics in decomposing foliar litter in Canadian forests[J]. Ecosystems, 2006, 9(1): 46-62. DOI:10.1007/s10021-004-0026-x |
| [18] | Schaller J, Hines J, Brackhage C, et al. Silica decouples fungal growth and litter decomposition without changing responses to climate warming and N enrichment[J]. Ecology, 2014, 95(11): 3181-3189. DOI:10.1890/13-2104.1 |
| [19] | Prado R, Erdocia X, Labidi J. Effect of the photocatalytic activity of TiO2 on lignin depolymerization[J]. Chemosphere, 2013, 91(9): 1355-1361. DOI:10.1016/j.chemosphere.2013.02.008 |
| [20] | Martínez A, Monroy S, Pérez J, et al. In-stream litter decomposition along an altitudinal gradient:does substrate quality matter?[J]. Hydrobiologia, 2016, 766(1): 17-28. DOI:10.1007/s10750-015-2432-9 |
| [21] | Boyero L, Pearson R G, Gessner M O, et al. A global experiment suggests climate warming will not accelerate litter decomposition in streams but might reduce carbon sequestration[J]. Ecology Letters, 2011, 14(3): 289-294. DOI:10.1111/ele.2011.14.issue-3 |
| [22] |
刘流, 刘德富, 肖尚斌, 等. 水温分层对三峡水库香溪河库湾春季水华的影响[J]. 环境科学, 2012, 33(9): 3046-3050. Liu L, Liu D F, Xiao S B, et al. Effects of thermal stratification on spring blooms in Xiangxi Bay of the Three Gorges Reservoir[J]. Environmental Science, 2012, 33(9): 3046-3050. |
| [23] | Foereid B, Bellarby J, Meier-Augenstein W, et al. Does light exposure make plant litter more degradable?[J]. Plant and Soil, 2010, 333(1): 275-285. |
| [24] | Day T A, Zhang E T, Ruhland C T. Exposure to solar UV-B radiation accelerates mass and lignin loss of Larrea tridentata litter in the Sonoran Desert[J]. Plant Ecology, 2007, 193(2): 185-194. DOI:10.1007/s11258-006-9257-6 |
| [25] | Harvey G R, Boran D A, Chesal L A, et al. The structure of marine fulvic and humic acids[J]. Marine Chemistry, 1983, 12(2-3): 119-132. DOI:10.1016/0304-4203(83)90075-0 |
| [26] | Tranvik L J, Bertilsson S. Contrasting effects of solar UV radiation on dissolved organic sources for bacterial growth[J]. Ecology Letters, 2001, 4(5): 458-463. DOI:10.1046/j.1461-0248.2001.00245.x |