 2017, Vol. 38
2017, Vol. 38



2. 暨南大学化学系, 广州 510632
2. Department of Chemistry, Jinan University, Guangzhou 510632, China
近年来, 随着城市污水量的增多, 城市污水处理厂规模和数量在不断扩大, 产生的剩余污泥越来越多.污泥中含有大量的有机质和营养物质(如N、P、K等), 同时还含有病原微生物、重金属及其它有害物质, 如果得不到合理的处理和处置, 必然会导致二次污染[1].堆肥因对环境危害小, 又符合可持续发展战略, 在实现污泥资源化方面受到广泛的关注.传统的污泥堆肥主要是利用堆肥原料中的土著微生物降解有机物, 但堆肥初期微生物量少, 繁殖较慢, 因此存在发酵时间长、产生臭味且肥效低等问题, 在产业化利用中受到限制[2].近年来, 添加外源微生物因对堆肥腐熟进程和堆肥产品质量有着积极的作用而备受关注. Sarkar等[3]在剩菜剩叶堆肥高温期添加培养好的嗜热菌, 细菌数量的增加提高了高温期微生物的活性, 加快了腐熟进程. Hachicha等[4]发现在橄榄油厂废水污泥的堆肥中接种变色栓菌可以增强腐殖酸的芳香化, 从而加速堆体腐熟. Zhang等[5]在园林废弃物的堆肥中添加白腐真菌, 发现可以提高堆体温度, 加快有机质的分解, 并且高温后再接种更利于堆体有机质的降解[6].
由于堆肥物料成分复杂, 外源微生物菌剂与堆肥土著微生物之间存在竞争, 致使二者均不能充分地发挥其作用[7], 因此, 了解堆肥过程中的微生物动态变化对充分发挥菌剂在堆肥中的作用显得尤为重要.目前多是采用PCR-DGGE技术来分析堆肥过程微生物群落变化, 通过电泳图谱中的几十个条带信息对微生物群落多样性进行定性或半定量的分析, 但这种技术对于群落中相对丰度低于1.00%的非优势微生物检测效果甚微[8].高通量测序技术可以对相对丰度0.01%~0.10%的微生物进行定性和定量的检测, 具有分析结果准确、高速、高灵敏度和高自动化等特点, 近年来已逐渐被应用到包括堆肥领域的微生物群落分析中[9].目前, 对污泥接菌堆肥过程微生物多样性及群落结构和其理化参数之间关系的研究还十分缺乏.本研究通过外接菌种, 并利用高通量测序技术, 从分子生态学的角度研究接菌对堆肥过程中的微生物群落多样性及群落结构的影响, 同时对理化参数进行监测, 并分析微生物群落与理化参数之间的关系, 以期为优化工艺参数及研究堆肥过程中微生物的作用机制提供理论依据.
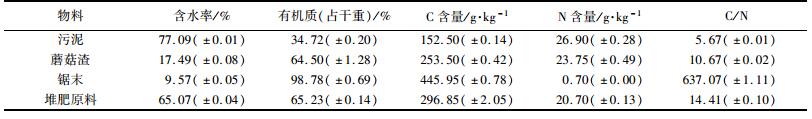
1 材料与方法 1.1 堆肥原料本实验以城市污水处理厂剩余污泥为堆肥基质, 锯末和蘑菇渣为辅料, 污泥取自广州市某污水处理厂, 锯末取自广州瑞兴木质加工厂, 蘑菇渣购自安徽六安心农园艺.实验所采用物料的性质见表 1.
|
|
表 1 堆肥原料的主要成分 Table 1 Primary composition of the composting materials |
1.2 微生物菌剂的制备
高温菌的制备:污泥堆肥过程中, 在高温期取适量堆肥新鲜样, 加入0.85%生理盐水, 堆肥新鲜样与生理盐水的重量体积比(g·mL-1)为1:10, 放入60℃恒温培养箱, 转速为160 r·min-1, 培养时间为24 h.取部分液体置于营养培养基中, 培养基组成为:牛肉膏3 g·L-1, 蛋白胨10 g·L-1, NaCl 15 g·L-1, 蒸馏水1 000 mL, pH为7.4~7.6.培养条件与之前相同, 培养时间为48 h, 得到高温菌, 稳定传至4代, 用于接种堆肥.
黄孢原毛平革菌孢子液的制备:黄孢原毛平革菌购自广东省微生物菌种保藏中心, 4℃斜面保存的菌种转接于PDA平板, 37℃培养5~7 d后, 平板表面形成丰富的白色粉状孢子, 用无菌水将孢子洗到灭菌的三角瓶中, 形成孢子悬液. PDA培养基组成为:20%马铃薯汁, 20 g·L-1葡萄糖, 3 g·L-1 KH2PO4, 1.5 g·L-1 MgSO4, 8 mg·L-1盐酸硫胺, 20 g·L-1琼脂, 剩余为蒸馏水, pH为6.0.
1.3 堆肥实验设计堆肥反应器为D 40 cm, H 40 cm的不锈钢桶, 在反应器外壁包一层厚度约为5 cm的保温棉.污泥与锯末、蘑菇渣按质量比10:2:1混合均匀, 采用间歇通风, 通风量为0.36 m3·h-1.堆肥设计2个处理:T1为接菌堆肥, 在第0 d接种高温菌, 接种量为2 g·kg-1, 以提高堆体温度, 在第12 d时接种黄孢原毛平革菌孢子液, 接种量为20 mL·kg-1, 使纤维素、木质素等难降解物质得到分解. T2为对照, 不加菌, 只添加等体积的无菌水.堆肥总时间为40 d, 取样时进行人工翻堆.
1.4 测定项目及方法 1.4.1 理化指标分析堆体温度采用多点监测取平均值的方法每天监测.新鲜堆样用去离子水按1:10(g·mL-1)浸提后, 滤液中的水溶性有机碳用TOC仪进行测定, pH用pH计进行测定.堆体含水率的测定采用恒重法.总氮通过元素分析仪测定, 新鲜堆样经KCl浸提后采用连续流动分析仪测定滤液中的NH4+-N和NO3--N[10].堆体有机质含量通过马弗炉550℃灼烧6 h进行测定.种子发芽指数(GI)的测定:新鲜堆样用去离子水按照固液比1:10浸提1 h, 吸取5 mL滤液注入到底部铺有滤纸的9 cm培养皿中, 并在培养皿中均匀放置20颗白菜种子, 用去离子水作对照, 每个处理设3个重复, 在25℃黑暗条件下培养48 h, 用公式(1) 测定种子发芽率[11].
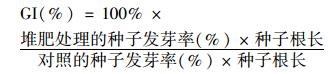
|
(1) |
分别取第0、4、19、33 d的堆肥样品, 进行高通量测序分析.采用Powersoil土壤DNA提取试剂盒对堆肥样品的细菌基因组DNA进行提取, 对16S V4高变区进行PCR扩增, 细菌的通用引物序列为515F(5′-CAATTCMTTTRAGTTT-3′)和806R(5′-GTGCCAGCMGCCGCGGTAA-3′).
测序结果分析:利用Trimmomatic软件对原始数据进行质量过滤, 得到质控后的有效数据.利用Mothur及Flash等软件对每对PE reads进行拼接, 获得原始的拼接序列.再利用Mothur对拼接后的序列进行质量控制及过滤, 得到有效的拼接片段.根据barcode及引物信息等, 利用Qiime将拼接后序列分配至相应的样品中.
多样性及群落结构分析:微生物群落的α多样性分析, 通常计算的参数包括Chao1、observed_species、Shannon、Simpson等.用unifrac的方法来计算加权矩阵以进行β多样性的分析.利用Qiime对每个OTU的代表序列进行物种分类, 生成不同分类水平上的物种丰度表.利用典型对应分析(CCA)研究堆体微生物群落和理化参数之间的关系.
2 结果与讨论 2.1 堆肥过程中理化参数的变化 2.1.1 温度、pH、有机质、WSOC、含水率的变化温度是反映堆体内微生物活性变化的一个重要指标, 如图 1(a)所示, T1和T2的温度在第2 d达到最高, 分别是70.75℃和68.10℃.两堆体在第12 d开始二次升温, T1和T2的温度在55℃以上分别持续了7 d和5 d, 因此两堆体符合无害化标准[12]. T1和T2的pH在第12 d达到最大, 分别为8.43和8.32[图 1(b)].到堆肥结束, 两堆体的pH分别为8.08、8.13.两堆体的有机质有一个明显的降低趋势[图 1(c)], T1和T2的有机质分别减少了9.23%、6.67%.堆肥中水溶性有机碳(WSOC)是能被微生物优先利用来进行新陈代谢的重要物质, 如图 1(d), 堆肥初期由于温度升高, 有机碳快速分解, WSOC的含量急剧增加[13].之后, 随着微生物的生长繁殖, 微生物大量消耗WSOC, 堆体中的WSOC含量开始降低, 接菌可以促进堆肥基质中水溶性有机碳的生物降解.堆肥结束, T1和T2的含水率分别减少了7.93%和5.19%[图 1(e)], 因反应器是密封的, 只在盖子上留有出气口, 产生的水蒸气不能完全排出, 使得堆体的含水率变化不明显.

|
图 1 堆肥过程理化参数的变化 Fig. 1 Changes of physicochemical parameters during the sludge composting |
总氮的变化如图 1(f)所示, 随着堆肥的进行, 堆体中的氮含量有所降低, 氮损失主要发生在高温期[9].到堆肥结束, T1和T2的氮含量分别损失了3.82%和7.54%.如图 1(g)所示, 在堆肥高温阶段, 高的氨化速率使得NH4+-N的浓度快速增加[14], 随着温度的降低, 积累的高浓度NH4+-N开始逐渐下降, 减少的NH4+-N部分以氨气的形式挥发损失, 部分经硝化细菌的作用转化为NO3--N, 部分被微生物固定转化为有机氮[15].在图 1(h)中, 两堆体中的NO3--N含量在整个堆肥阶段呈上升趋势.在最初阶段, 由于高温和高浓度的NH4+-N, 抑制了硝化细菌的活性, 所以堆肥初始阶段NO3--N的浓度增加缓慢.随着温度的降低, 硝化反应增强, 生成的NO3--N浓度开始增加[16].在两次接种的共同作用下, T1中硝态氮的含量增加得比T2中的多.
2.1.3 种子发芽率种子发芽率(GI)是评价最终堆肥产品毒性的重要指标[17].当GI大于80%时, 可以认为堆肥完全腐熟[18].由图 1(i)可知, 堆肥初期有机质的分解, 产生了大量的NH4+-N和小分子有机酸, 且堆体本身含有一些有害于作物生长的物质, 抑制了种子的发芽.随着堆肥进行, 小分子有机酸被分解, 或者转化成高分子腐殖酸, 同时氨的挥发和重金属固定使得GI升高[19].堆肥结束后, T1和T2的GI分别为81.86%和64.87%.
2.2 细菌群落相似性和多样性分析细菌群落相似性可以通过PCoA和UPGMA聚类进行分析.细菌群落的分布主要包括三块[图 2(a)], 堆肥初期的样品(第0 d)、高温期的样品(第4 d)、降温期和腐熟期的样品(第19、33 d).第0、4、19 d的样品相距较远, 说明堆肥过程尤其是在前19 d两堆体中细菌群落发生了较大的演变.而堆肥19 d后样品聚在一起, 说明这些样品中细菌群落有高度的相似性, 降温腐熟阶段的细菌群落结构有着高度稳定性[20]. T1和T2在相同时期的样品相距很近, 说明添加的菌剂不会对细菌群落结构造成明显的改变.从UPGMA聚类分析[图 2(b)]也可以看出, 同一堆肥的不同阶段的细菌群落结构相似性较低, 如T1的高温期和降温期的相似性系数为78.00%.同一时期不同堆肥的细菌群落结构相似性较高[21], 比如T1、T2的高温期的相似性系数为96.50%.

|
图 2 PCoA和UPGMA聚类分析 Fig. 2 Analysis of principal coordinates and arithmetic mean clustering |
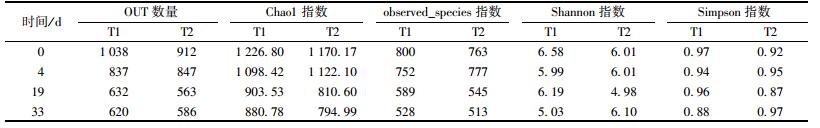
observed_species指数和Chao1指数反映样品中群落的丰富度. Shannon指数和Simpson指数反映的是群落的多样性, 受样品群落中物种丰富度和物种均匀度的影响.如表 2所示, 随着堆肥的进行, 两个堆体的observed_species值和Chao1值降低, 说明堆肥会降低群落的丰富度, OUT数量的变化也印证了这个结果.在第4 d, T2中细菌群落的丰富度和多样性都比T1大, 这是因为T1、T2都处于高温期, 且T1温度比T2高, 能杀死更多的微生物, 使得群落的丰富度和多样性降低得更多.在第0、19 d, 因为外源菌种的添加, T1细菌群落的丰富度和多样性高于T2的.堆肥至第33 d, T1群落的丰富度高于T2的, 但受物种均匀度的影响, T2群落的多样性高于T1.
|
|
表 2 各个样品细菌群落多样性 Table 2 Diversity of bacterial community in each sample |
2.3 堆体细菌群落分布
如图 3(a)所示, 在门分类水平上, 所有样品中主要的8个门为放线菌门(Actinobacteria)、装甲菌门(Armatimonadetes)、拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)、浮霉菌门(Planctomycetes)、变形菌门(Proteobacteria)和互养菌门(Synergistetes).不同时期在门水平各细菌群落的相对丰度不同, 第0 d时, T1和T2中拟杆菌门(12.75%、31.11%)、厚壁菌门(22.28%、17.87%)、浮霉菌门(18.38%、11.02%)、变形菌门(17.48%、17.87%)比较丰富.第4 d(高温期)T1和T2中厚壁菌门相对丰度大量增加, 分别为74.66%、72.49%, 有研究报道显示厚壁菌门在高温期是最优势菌门, 这主要与厚壁菌门可以在高温下(>55℃)存活, 并能参与各种代谢活动有关[9, 22].与堆肥初始期相比, 高温期除了厚壁菌门和放线菌门, 其它菌门的相对丰度都降低了.且随着温度的降低, 微生物的活性开始再次增强, 厚壁菌门所占比例下降, 放线菌门、变形菌门所占比例增加.第19 d(降温期)时, T1和T2中放线菌门(30.17%、51.51%)、厚壁菌门(32.50%、27.91%)、变形菌门(27.36%、12.69%)占有较大比例.堆肥至第33 d(腐熟期)时, T1和T2中放线菌门(57.85%、27.86%)、厚壁菌门(19.21%、29.13%)、变形菌门(11.16%、19.31%)是优势菌.随着堆肥的进行, 放线菌门的相对丰度呈上升趋势, 放线菌门可以通过分泌各种抗生素抑制病原微生物[22], 因此, 放线菌门的增多利于堆肥产品生物毒性的降低.从图 1(i)也可以看出, 第19 d后, 两堆体的种子发芽率快速增加.

|
图 3 样品中门和属水平细菌群落相对丰度 Fig. 3 Relative abundances of phyla and genera in each sample |
在属水平上, 各样品中细菌群落的相对丰度分布如图 3(b)所示, 堆肥初期, T1中优势细菌主要为Mangroviflexus、Exiguobacterium、Zavarzinella、Caldilinea, 其中Mangroviflexus(9.67%)、Zavarzinella(9.87%)的相对丰度较高, 分别属于拟杆菌门、浮霉菌门, T2中优势细菌为Mangroviflexus、Zavarzinella、Caldilinea、Rikenella, 其中Mangroviflexus(25.23%)相对丰度较高. Mangroviflexus是一种可以发酵各种底物产生乙酸、丙酸、琥珀酸的厌氧嗜温菌[23], 主要存在于污泥中.进入高温期后, 堆肥初期的优势细菌的相对丰度降低, 新的优势细菌出现. Ureibacillus、Symbiobacterium是T1、T2中主要优势细菌, 属于厚壁菌门, 耐高温, 两者的相对丰度在T1中更高, 分别是12.94%、11.88%. Ureibacillus既能分解可溶性有机物, 同时可以产生胞外水解酶, 分解纤维素、半纤维素大分子化合物[24], 经常在堆肥高温阶段被检测到. Symbiobacterium常从堆肥、土壤、动物粪便中分离出[25].在降温期, T1、T2中优势细菌主要为Actinomadura、Alkalilimnicola、Paenibacillus、Calditerricola. T1中Alkalilimnicola(8.51%)所占比例较高, 属于变形菌门. T2中Calditerricola(9.44%)所占比例较高, 属于厚壁菌门. Alkalilimnicola是好氧菌, 但在厌氧条件下, 可以将硝酸盐或N2O作为电子受体生长[26].
随着堆体温度降至与室温接近, 堆肥进入腐熟期. T1、T2中优势细菌主要为Actinomadura、Alkalilimnicola、Persicitalea、Paenibacillus.在两个堆体中, 属于放线菌门的Actinomadura所占比例最大, 分别为17.81%、9.48%.有学者发现在堆肥后期Actinomadura会出现, 因此提出可以将Actinomadura作为评价堆体腐熟的相关指标之一[27].在本研究中, 第19 d时, Actinomadura是优势菌属, 且T1和T2中第33 d与第19 d样品中的优势菌门一致, 说明在19 d时, 堆体可能已开始腐熟.在不同时期, 厚壁菌门一直是优势菌门, 而接种的高温菌中的厚壁菌门的相对丰度为99.06%[图 4(a)], 高温菌的接入增加了T1厚壁菌门的相对丰度, 使T1中厚壁菌门的相对丰度在前19 d比T2高, 而且高温菌的优势菌属Ureibacillus在堆肥高温期也是优势菌属[图 4(b)].菌剂的添加会影响优势菌所占比例, 但不会改变优势菌的种类.

|
图 4 门和属水平高温菌群落相对丰度 Fig. 4 Relative abundances of phyla and genera in the thermophilic bacteria |
采用CCA分析堆体理化参数(温度、pH、有机质、WSOC、含水率、TN)与主要菌属之间的关系.如图 5, CCA1和CCA2分别解释了43.78%和35.45%的样品差异, 前2个排序轴共解释了79.23%的细菌种群结构变化, 说明这6个理化参数对堆肥过程中细菌群落结构的变化有显著的影响.可把图中主要菌属分为3簇(Ⅰ、Ⅱ、Ⅲ), 从理化参数和主要菌属与原点连线之间的夹角可以看出理化参数与细菌群落的关系, 第Ⅰ簇内的6个菌属与温度呈正相关关系; 第Ⅱ簇内的12个菌属与含水率、有机质、总氮呈正相关, 与WSOC、pH呈负相关; 第Ⅲ簇内的21个菌属与WSOC、pH呈正相关关系, 另外第Ⅲ簇内的Aciditerrimonas、Aeribacillus、Aneurinibacillus还与温度呈正相关. T1中与温度相关的9个菌属的总相对丰度在4个样品中都比T2多, 且在第4 d的样品中最多, 这是出现T1温度高于T2, 并在第4 d(与第0、19、33 d相比)温度达最高的原因. T1和T2堆肥原料一样, 有无菌剂添加是二者唯一的差异, 因此可知外接菌种是引起T1温度高于T2的主要原因.由理化参数箭头的连线长度可知, 不同的理化参数对细菌群落结构的影响不同, pH对细菌群落结构影响最大, 其次是WSOC、有机质和温度, 含水率和TN的影响稍弱.因此可通过改变堆体pH来影响微生物群落结构, 从而改善堆肥质量, 有学者就通过在污泥堆肥中添加不同pH的锯末来降低氮损失[16].

|
图 5 细菌群落与理化参数的典型对应分析 Fig. 5 CCA of physicochemical parameters and bacterial community structure |
(1) 外接菌种可以提高堆体温度, 延长高温持续时间, 降低氮损失, 加快堆肥腐熟, 提高堆体中细菌群落的丰富度.在整个堆肥过程中, 细菌群落结构发生了较大的演变, 同一堆肥的不同阶段的细菌群落结构相似性较低, 同一时期不同堆肥的细菌群落结构相似性较高.
(2) 在不同时期, 厚壁菌门一直是优势菌门, 高温菌的接入增加了堆体中厚壁菌门的相对丰度, 使得T1中厚壁菌门的相对丰度在前19 d比T2高, 且添加的高温菌中优势菌属Ureibacillus在堆肥高温期也是优势菌属.菌剂的添加会影响优势菌所占比例, 但不会改变优势菌的种类.
(3) 不同的理化参数对细菌群落结构影响不同, pH对细菌群落结构影响最大, 其次是WSOC、有机质和温度, 含水率和TN的影响稍弱.温度主要与9个菌属呈正相关关系.
| [1] | Wu C D, Li W G, Wang K, et al. Usage of pumice as bulking agent in sewage sludge composting[J]. Bioresource Technology, 2015, 190: 516-521. DOI:10.1016/j.biortech.2015.03.104 |
| [2] |
张陇利, 刘青, 徐智, 等. 复合微生物菌剂对污泥堆肥的作用效果研究[J]. 环境工程学报, 2008, 2(2): 266-269. Zhang L L, Liu Q, Xu Z, et al. Research on effect of compound microbial inoculant on sewage sludge composting[J]. Chinese Journal of Environmental Engineering, 2008, 2(2): 266-269. |
| [3] | Sarkar S, Banerjee R, Chanda S, et al. Effectiveness of inoculation with isolated Geobacillus strains in the thermophilic stage of vegetable waste composting[J]. Bioresource Technology, 2010, 101(8): 2892-2895. DOI:10.1016/j.biortech.2009.11.095 |
| [4] | Hachicha R, Rekik O, Hachicha S, et al. Co-composting of spent coffee ground with olive mill wastewater sludge and poultry manure and effect of Trametes versicolor inoculation on the compost maturity[J]. Chemosphere, 2012, 88(6): 677-682. DOI:10.1016/j.chemosphere.2012.03.053 |
| [5] | Zhang J C, Zeng G M, Chen Y N, et al. Impact of Phanerochaete chrysosporium inoculation on indigenous bacterial communities during agricultural waste composting[J]. Applied Microbiology and Biotechnology, 2013, 97(7): 3159-3169. DOI:10.1007/s00253-012-4124-y |
| [6] | Varma V S, Ramu K, Kalamdhad A S. Carbon decomposition by inoculating Phanerochaete chrysosporium during drum composting of agricultural waste[J]. Environmental Science and Pollution Research, 2015, 22(10): 7851-7858. DOI:10.1007/s11356-014-3989-y |
| [7] |
徐智, 张陇利, 张发宝, 等. 接种内外源微生物菌剂对堆肥效果的影响[J]. 中国环境科学, 2009, 29(8): 856-860. Xu Z, Zhang L L, Zhang F B, et al. Effects of indigenous and exogenous microbial inocula on composting in a bioreactor[J]. China Environmental Science, 2009, 29(8): 856-860. |
| [8] |
宋云龙, 张金松, 朱佳, 等. 基于高通量测序的微生物强化污泥减量工艺中微生物群落解析[J]. 中国环境科学, 2016, 36(7): 2099-2107. Song Y L, Zhang J S, Zhu J, et al. Analysis of microbial community in in-situ sludge reduction process by bioaugmentation using high-throughput sequencing technology[J]. China Environmental Science, 2016, 36(7): 2099-2107. |
| [9] | Ren G M, Xu X H, Qu J J, et al. Evaluation of microbial population dynamics in the co-composting of cow manure and rice straw using high throughput sequencing analysis[J]. World Journal of Microbiology and Biotechnology, 2016, 32(6): 101. DOI:10.1007/s11274-016-2059-7 |
| [10] | Jiang J S, Liu X L, Huang Y M, et al. Inoculation with nitrogen turnover bacterial agent appropriately increasing nitrogen and promoting maturity in pig manure composting[J]. Waste Management, 2015, 39: 78-85. DOI:10.1016/j.wasman.2015.02.025 |
| [11] | Zucconi F, Pera A, Forte M, et al. Evaluating toxicity of immature compost[J]. BioCycle, 1981, 22(2): 54-57. |
| [12] |
GB 7959-1987, 粪便无害化卫生标准[S]. GB 7959-1987, Sanitary standard for the non-hazardous treatment of night soil[S]. |
| [13] |
张雪辰, 邓双, 杨密密, 等. 畜禽粪便堆腐过程中有机碳组分与腐熟指标的变化[J]. 环境科学学报, 2014, 34(10): 2559-2565. Zhang X C, Deng S, Yang M M, et al. Changes of organic carbon fractions and maturity index during composting of manure[J]. Acta Scientiae Circumstantiae, 2014, 34(10): 2559-2565. |
| [14] | Zhang L, Sun X Y. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar[J]. Bioresource Technology, 2014, 171: 274-284. DOI:10.1016/j.biortech.2014.08.079 |
| [15] | Rashad F M, Saleh W D, Moselhy M A. Bioconversion of rice straw and certain agro-industrial wastes to amendments for organic farming systems: 1.composting, quality, stability and maturity indices[J]. Bioresource Technology, 2010, 101(15): 5952-5960. DOI:10.1016/j.biortech.2010.02.103 |
| [16] | Li Y B, Li W G. Nitrogen transformations and losses during composting of sewage sludge with acidified sawdust in a laboratory reactor[J]. Waste Management & Research, 2015, 33(2): 139-145. |
| [17] | Awasthi M K, Pandey A K, Bundela P S, et al. Co-composting of gelatin industry sludge combined with organic fraction of municipal solid waste and poultry waste employing zeolite mixed with enriched nitrifying bacterial consortium[J]. Bioresource Technology, 2016, 213: 181-189. DOI:10.1016/j.biortech.2016.02.026 |
| [18] | Wu D L, Liu P, Luo Y Z, et al. Nitrogen transformations during co-composting of herbal residues, spent mushrooms, and sludge[J]. Journal of Zhejiang University-Science B, 2010, 11(7): 497-505. DOI:10.1631/jzus.B0900271 |
| [19] | Banegas V, Moreno J L, Moreno J I, et al. Composting anaerobic and aerobic sewage sludges using two proportions of sawdust[J]. Waste Management, 2007, 27(10): 1317-1327. DOI:10.1016/j.wasman.2006.09.008 |
| [20] |
张晶, 孙照勇, 钟小忠, 等. 奶牛粪条垛式模拟堆肥腐熟度及微生物群落结构变化[J]. 应用与环境生物学报, 2016, 22(3): 423-429. Zhang J, Sun Z Y, Zhong X Z, et al. Maturity and microbial community structure dynamics during simulated windrow composting of solid fraction of dairy manure[J]. Chinese Journal of Applied and Environmental Biology, 2016, 22(3): 423-429. |
| [21] |
党秋玲, 李鸣晓, 席北斗, 等. 堆肥过程多阶段强化接种对细菌群落多样性的影响[J]. 环境科学, 2011, 32(9): 2689-2695. Dang Q L, Li M X, Xi B D, et al. Effects of multi-stage strengthening inoculation on bacterial community diversity during composting[J]. Environmental Science, 2011, 32(9): 2689-2695. |
| [22] | Tian W, Sun Q, Xu D B, et al. Succession of bacterial communities during composting process as detected by 16S rRNA clone libraries analysis[J]. International Biodeterioration & Biodegradation, 2013, 78: 58-66. |
| [23] | Ziganshina E E, Belostotskiy D E, Ilinskaya O N, et al. Effect of the organic loading rate increase and the presence of zeolite on microbial community composition and process stability during anaerobic digestion of chicken wastes[J]. Microbial Ecology, 2015, 70(4): 948-960. DOI:10.1007/s00248-015-0635-2 |
| [24] | Zhang J Y, Cai X, Qi L, et al. Effects of aeration strategy on the evolution of dissolved organic matter (DOM) and microbial community structure during sludge bio-drying[J]. Applied Microbiology and Biotechnology, 2015, 99(17): 7321-7331. DOI:10.1007/s00253-015-6640-z |
| [25] | Nakasaki K, Tran L T H, Idemoto Y, et al. Comparison of organic matter degradation and microbial community during thermophilic composting of two different types of anaerobic sludge[J]. Bioresource Technology, 2009, 100(2): 676-682. DOI:10.1016/j.biortech.2008.07.046 |
| [26] | Sorokin D Y, Zhilina T N, Lysenko A M, et al. Metabolic versatility of haloalkaliphilic bacteria from soda lakes belonging to the Alkalispirillum-Alkalilimnicola group[J]. Extremophiles, 2006, 10(3): 213-220. DOI:10.1007/s00792-005-0487-7 |
| [27] | Xu S W, Reuter T, Gilroyed B H, et al. Microbial communities and greenhouse gas emissions associated with the biodegradation of specified risk material in compost[J]. Waste Management, 2013, 33(6): 1372-1380. DOI:10.1016/j.wasman.2013.01.036 |