 2017, Vol. 38
2017, Vol. 38



厌氧氨氧化反应是厌氧氨氧化菌在厌氧条件下,以NH4+作为电子供体,NO2-作为电子受体,将NH4+和NO2-转化为N2的生物氧化过程,并且不用外加有机碳源[1~3].近年来,国内外研究学者在海洋水体和沉积物以及极端环境下发现了海洋厌氧氨氧化菌的存在[5~9].到目前为止,已经发现有5属14种厌氧氨氧化菌[10, 11],其中“Candidatus Scalindua”在海洋中占主要的地位[12, 13], 同时也有很多学者利用海洋沉积物成功启动海洋厌氧氨氧化反应器[14, 15].有研究表明,海洋厌氧氨氧化菌产生的N2占海洋氮循环的30%~50%[1~4].这些发现使人们对海洋氮循环有了新的认识,对研究海洋生态学和海洋氮元素生物地球化学循环都具有重要的意义.除此之外,沿海城市由于淡水匮乏引起的海水直接利用导致这些地区生活污水盐度升高,较高的盐度对常规脱氮微生物的生理活性非常不利,从而限制了其脱氮效能的发挥.海洋厌氧氨氧化菌来自海洋环境中,能够耐受较高的盐度,因此,其在高盐度废水的脱氮处理方面有着良好的应用前景[16~19].
海洋厌氧氨氧化菌的富集与培养需要严格的反应条件,反应条件被破坏,恢复其活性则需要较长的时间.有研究表明,与生长在淡水条件下的厌氧氨氧化菌相比,海洋厌氧氨氧化菌的最适温度较低[20],尤其是在北极海洋沉积污泥中,厌氧氨氧化菌的最适活性温度低于15℃,甚至在-1.3℃的情况下也可以存活[20~22].这些研究海洋厌氧氨氧化菌具有适应低温的能力.除此之外,为了改善海洋厌氧氨氧化脱氮过程,可靠的动力学参数的获取和合适的动力学模型的建立能够提高生物反应过程的可预测性.目前,国内尚未有关于海洋厌氧氨氧化动力学研究的报道,国外有文章阐明了Stover-Kincannon模型适合模拟海洋厌氧氨氧化反应器的脱氮性能[23],对海洋厌氧氨氧化脱氮起到了指导作用.
鉴于当前海洋厌氧氨氧化研究相对较少、其影响因素和机制尚不清楚的现状,本研究采用海洋厌氧氨氧化菌处理含海水污水,通过改变反应器内温度,分析了不同温度对海洋厌氧氨氧化脱氮效能的影响,建立反应器稳定运行的调控策略,并采用Logistic模型分析了不同温度情况下海洋厌氧氨氧化反应的动力学特性,以期为海洋厌氧氨氧化菌处理含海水污水提供理论基础和技术支持.
1 材料与方法 1.1 实验装置本实验采用ASBR反应器 (如图 1),有效容积为7 L,系有机玻璃制成.其利用海洋底泥经过一年成功启动,无污染其他菌源体,并稳定运行5个月,NH4+-N和NO2--N基本可以完全去除.反应器内的温度通过温控箱控制,整个反应器用锡纸包裹,防止光对海洋厌氧氨氧化菌活性的影响.反应区内悬挂无纺滤布作为海洋厌氧氨氧化菌的生物膜载体,反应器内污泥主要以红色颗粒为主,粒径特征分布如图 2所示, 其中频率指的是某一粒径范围内,颗粒污泥数量占样品总量的比值.实验采用人工配制的模拟废水,经高纯氮气吹脱15 min后 (DO < 0.2 mg·L-1),通过流动泵进水.反应器进水pH控制在7.5±0.1,运行一个周期包括进水4 min,反应6 h,静置25 min,出水4 min.

|
1.取样口及进出水口;2.搅拌;3.储水箱;4.水浴进出水口;5.水浴恒温控制器;6.进水蠕动泵 图 1 海洋厌氧氨氧化反应器装置 Fig. 1 Schematic diagram of the marine anammox reactor |

|
图 2 颗粒污泥粒径分布 Fig. 2 Diameter distribution of granular sludge |
实验所用海水取自胶州湾 (黄海北部),其中约70%为NaCl,约14%为MgCl2,平均盐度为32‰,经人工配制成模拟废水,其主要成分为:29 mg·L-1 KH2PO4,136 mg·L-1 CaCl2,1 200 mg·L-1 KHCO3,300 mg·L-1 MgSO4·7H2O.同时投加微生物所需的微量元素,微量元素Ⅰ:EDTA 5 000 mg·L-1,FeSO4·7H2O 5 000 mg·L-1,微量元素Ⅱ:EDTA 15 000 mg·L-1,H3BO3 11 mg·L-1,MnCl2·4H2O 990 mg·L-1,CuSO4·5H2O 250 mg·L-1,ZnSO4·7H2O 430 mg·L-1,NiCl2·6H2O 190 mg·L-1,Na2MoO4·2H2O 220 mg·L-1,CoCl2·6H2O 240 mg·L-1,NaSeO4·10H2O 210 mg·L-1. NH4+-N和NO2--N分别用NH4Cl和NaNO2提供.
1.3 分析方法NH4+-N:纳氏试剂比色法[24];NO2--N:N-(1-萘基)-乙二胺光度法[24];NO3--N:麝香草酚分光光度法[24];pH值:WTW pH/Oxi 340i;温度:水银温度计.总氮容积负荷 (TNLR) 根据进水总氮浓度与HRT关系得到,总氮去除负荷 (TNRR) 根据进、出水总氮浓度与HRT关系得到,即:
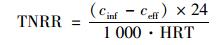
|
(1) |

|
(2) |
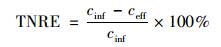
|
(3) |
式中,cinf为进水总氮浓度 (mg·L-1),ceff为出水总氮浓度 (mg·L-1),HRT为水力停留时间 (HRT=6 h,采样间隔t=0.5 h),TNRE为总氮去除率 (%).
Arrhenius方程:
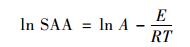
|
(4) |
式中,SAA为最大比反应速率,g·(m·d)-1;A为Arrhenius常数;E为活化能,J·mol-1;R为气体常数,8.314 J·(K·mol)-1;T为热力学温度,K.
1.4 动力学模型修正的Logistic模型[25]如下:

|
(5) |
把式 (3)、(5) 合并得到:
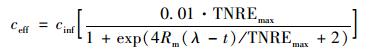
|
(6) |
式中,TNREmax为最大总氮去除率,%;Rm为最大脱氮速率,%·h-1,λ为海洋厌氧氨氧化菌脱氮的延迟时间,h;t为周期内某时刻,h.
2 结果与讨论 2.1 温度对海洋厌氧氨氧化菌脱氮效能的影响为了防止温度对海洋厌氧氨氧化菌产生不可逆的影响,本实验研究了35、30、25、20、15、10℃下海洋厌氧氨氧化菌的脱氮效能,结果如图 3所示.在25~35℃时,NH4+-N和NO2--N的去除率保持在99%左右,TNRR基本保持在 (0.62±0.01) kg·(m3·d)-1,TNRE的平均值为83.46%,此时的温度范围对反应器的脱氮效能影响不大.
本实验进行到第38 d,温度下降到20℃,反应器的脱氮效能开始受到微弱影响.出水NH4+-N和NO2--N浓度出现较小的上升趋势,同时TNRR由最初平均值为0.62 kg·(m3·d)-1下降至0.53 kg·(m3·d)-1,TNRE下降了6.01%.随后,反应器在20℃恒温下运行了13 d,出水NH4+-N和NO2--N浓度降低至稳定,最终基本可以全部去除,TNRR提高到0.62 kg·(m3·d)-1,TNRE也提高至79.24%.实验结果表明:温度在20℃时,TNRE和TNRR均出现了提高的现象,说明海洋厌氧氨氧化菌在20℃时仍具有较强的脱氮能力, 这一结果与Kawagoshi等[26]的研究结果相当.同时,与淡水厌氧氨氧化菌相比,其在较低温度处理含海水污水更具有实际意义.于德爽等[27]的实验结果显示,温度在20~25℃时,反应器内的厌氧氨氧化与反硝化耦合反应开始受到明显的抑制,并且未出现脱氮效能和TNRR提高的现象.
第52 d时,温度降到15℃,出水NH4+-N、NO2--N的浓度突然明显上升,其平均值分别为34.83 mg·L-1和69.00 mg·L-1,TNRE的平均值为39.44%,而且TNRR从最初的0.62 kg·(m3·d)-1下降至0.30 kg·(m3·d)-1.由此可以看出,厌氧氨氧化反应已受到抑制.李祥等[28]的研究表明,温度在20℃时,反应器氮去除率开始迅速下降,低于15℃时,出现了NO2--N的积累从而造成亚硝酸盐抑制反应器脱氮作用的现象,而本实验中,此温度下并未出现明显的NO2--N积累现象,这可能与海洋厌氧氨氧化菌能够耐较低温有关,此时只有温度影响其脱氮能力.当温度下降至10℃时,出水NH4+-N、NO2--N浓度的平均值分别提高至54.69 mg·L-1和90.93 mg·L-1,TNRE的平均值为10.71%,而且TNRR下降至0.08 kg·(m3·d)-1,海洋厌氧氨氧化菌的脱氮效能出现恶化,此温度条件不利于反应器的运行.
由图 3(d)可以看出,随着温度的变化,ΔNO2--N/ΔNH4+-N,ΔNO2--N/ΔNH4+-N也出现较小的变化.在25~35℃时,ΔNH4+-N:ΔNO2--N:ΔNO3--N的平均值为1:1.24:0.20,在10~15℃时,ΔNH4+-N:ΔNO2--N:ΔNO3--N的平均值为1:1.06:0.31,在20℃时, ΔNH4+-N/ΔNO2--N/ΔNO3--N的平均值为1:1.28:0.24,这与Strous等[3]研究的理论比值1:1.32:0.26是具有一定偏差的.但不难看出,在20~35℃时,计量比与理论比值偏差较小,反应器内厌氧氨氧化反应起到主要作用.而在10~15℃时,化学计量比的偏差较大,可能是反应器内存在其他少量同源自养菌与海洋厌氧氨氧化菌所适应的温度不同,从而其脱氮能力被抑制的程度不同.

|
图 3 不同温度下反应器内脱氮指标的变化 Fig. 3 Changes of nitrogen removal index in reactor at different temperature |
图 4为不同温度下,反应器在单个周期内NH4+-N、NO2--N、NO3--N以及pH的变化.由图 4(a)~4(c)可以看出,NH4+-N和NO2--N在不同温度的周期内均可同步去除,且伴有NO3--N同步生成.在25~35℃时,NH4+-N和NO2--N在5 h内均可以完全去除,温度对海洋厌氧氨氧化菌的脱氮效能影响较小,当温度降至20℃时,反应器受到轻微的影响,NH4+-N在5.5 h后完全去除,NO2--N在6 h后可基本去除.随后温度在15℃时,NH4+-N和NO2--N的反应速度明显减小.而在10℃时,6 h内的NH4+-N和NO2--N去除率仅为18.97%和8.82%,此时海洋厌氧氨氧化菌的活性受到严重抑制.由于厌氧氨氧化是产碱反应,脱氮能力受到不同温度的影响,pH的变化范围也有所差别,如图 4(d)可知,温度在25~35℃时,pH变化范围基本一致,均由7.49±0.01提高至8.04±0.01.在20℃时,pH变化过程出现波动,此温度下海洋厌氧氨氧化菌受到轻微影响,导致脱氮效能开始不稳定,经过6 h后,pH由初始值7.48提高至7.96,NH4+-N和NO2--N去除率的平均值分别为81.50%、68.60%.随着温度的降低,pH变化范围也开始减小.当温度降至15℃时,pH由7.5提高至7.75.然而在10℃时,pH仅由7.5提高至7.56,此时海洋厌氧氨氧化菌的脱氮效能受到严重削弱.

|
图 4 不同温度下反应器周期内脱氮指标的变化 Fig. 4 Changes of the nitrogen removal index in the reactor during the operation cycle at different temperatures |
温度对海洋厌氧氨氧化比反应速率的影响如图 5所示.以ln SAA为纵坐标,1/T为横坐标得到拟合曲线.在反应周期内,以NH4+-N为标准计算不同温度下的最大比反应速率,再根据Arrhenius方程得到反应所需的活化能.通过图 5可以看出,在10~35℃范围内,存在两个阶段的活化能.因为厌氧氨氧化反应进行的过程中,可能涉及两种不同最适温度的酶,而且厌氧氨氧化菌种类的丰富度也会随温度的变化而变化,这可能也是在不同温度范围内存在两个活化能的原因[29].除此之外,从图 5亦可看出,温度在25~35℃时,活化能为26.280 kJ·mol-1,随着温度的降低,海洋厌氧氨氧化的最大比反应速率也随之降低,在10~25℃时,活化能为76.613 kJ·mol-1.这一结果低于Isaka等[29]和Lotti等[30]的研究结论,Isaka认为淡水厌氧氨氧化在28~37℃时,活化能为33 kJ·mol-1;在28~22℃时的活化能为93 kJ·mol-1;在6~22℃时的活化能为94 kJ·mol-1.而Lotti分析得出,温度在30~25、25~20、20~15、15~10℃时,活化能分别为 (46.6±2.7)、(67.6±7.1)、(104.9±13.3)、(230.5±8.3) kJ·mol-1, 由此可以看出,与淡水厌氧氨氧化相比,海洋厌氧氨氧化在低温下更容易进行.

|
图 5 海洋厌氧氨氧化反应的Arrhenius拟合关系 Fig. 5 Arrhenius fitting of the marine ANAMMOX reaction |
采用修正的Logistic模型分析不同温度下海洋厌氧氨氧化反应的动力学特性.以周期内某时刻t为横坐标,周期内TNRE为纵坐标进行拟合,结果如图 6所示.从中可以看出,修正的Logistic模型比较适合分析不同温度下海洋厌氧氨氧化菌脱氮过程的动力学特性,同时能够较好地预测反应器内总氮去除率以及出水总氮浓度.通过拟合得到了TNREmax、Rm、λ这3个动力学参数,以及TNRE和ceff的预测公式,如表 1所示.

|
图 6 修正后Logistic模型的拟合曲线 Fig. 6 Fitting curves of modified Logistic model |
|
|
表 1 不同温度下动力学参数及公式的对比 Table 1 Comparison of kinetic parameters and formulas at different temperatures |
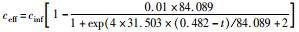


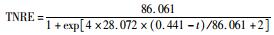
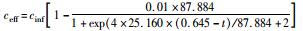
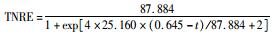

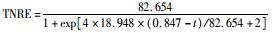
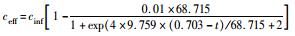
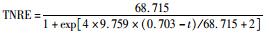
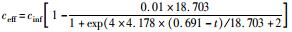
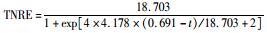
有报道指出,动力学参数会受到反应器的结构和性能、微生物种类和生理状态的影响[31],而且,污泥颗粒的大小也会明显影响动力学参数[32].在本实验中,动力学参数直接受到温度的影响而改变.温度在25~35℃范围内,TNREmax、Rm、λ这3个参数的变化并不明显,根据表 1比较此温度范围内预测TNRE的值与实际测得的值,其平均相对误差分别为 (84.089±2.670)%、(86.061±7.492)%、(87.884±12.480)%,说明在此温度范围内,厌氧氨氧化的反应速率与温度之间的关系可以用该模型描述.当温度为20℃时,虽然厌氧氨氧化反应的延迟时间略有增加,但这并不影响其脱氮能力,反应器在低温下处理含海水污水的潜能需要进一步开发.当温度降到10~15℃时,厌氧氨氧化反应的延迟时间明显增加,尤其在10℃时,TNREmax和Rm出现了大幅度的降低,TNRE的预测值与实际值相对误差较大,低温的抑制作用可能是不适合Logistic模型描述海洋厌氧氨氧化脱氮过程的原因,由此可知,对于低温度条件下,Logistic模型不适合描述其脱氮行为,在对海洋厌氧氨氧化反应器处理含海水污水的运行策略进行调控时,可将温度控制在15℃以上,防止低温的抑制作用.
3 结论(1) 温度25~35℃时,温度对反应器的脱氮效能较小.当温度降到10~15℃时,反应器的脱氮效能受到明显的抑制,TNRR由起始的0.62 kg·(m3·d)-1分别下降至0.30 kg·(m3·d)-1和0.08 kg·(m3·d)-1,TNRE分别为39.44%和10.71%.温度为20℃时,TNRE由初始值0.53 kg·(m3·d)-1提高到0.62 kg·(m3·d)-1,说明反应器在较低温度下处理含海水污水具有较大的潜能,对实际应用有着重要的意义.
(2) 根据Arrhenius方程得到不同温度下所需的活化能,温度在25~35℃时,活化能为26.280 kJ·mol-1;在10~25℃时,活化能为76.613 kJ·mol-1.与淡水厌氧氨氧化反应相比,海洋厌氧氨氧化反应在低温条件下更容易进行.
(3) 修正的Logistic模型比较适合拟合海洋厌氧氨氧化的脱氮过程,同时得到了不同温度下TNREmax、Rm、λ的值和TNRE、ceff的预测公式.在25~35℃之间,该模型可以较好地描述反应速率与温度之间的关系,但在10~20℃之间,实测值与预测值相对误差较大.
| [1] | Mulder A, van de Graaf A A, Robertson L A, et al. Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor[J]. FEMS Microbiology Ecology, 1995, 16(3): 177–183. DOI: 10.1111/fem.1995.16.issue-3 |
| [2] | Jetten M S M, van Niftrik L, Strous M, et al. Biochemistry and molecular biology of anammox bacteria[J]. Critical Reviews in Biochemistry and Molecular Biology, 2009, 44(2-3): 65–84. DOI: 10.1080/10409230902722783 |
| [3] | Strous M, Heijnen J J, Kuenen J G, et al. The sequencing batch reactor as a powerful tool for the study of slowly growing anaerobic ammonium-oxidizing microorganisms[J]. Applied Microbiology and Biotechnology, 1998, 50(5): 589–596. DOI: 10.1007/s002530051340 |
| [4] | Capone D G, Knapp A N. Oceanography:a marine nitrogen cycle fix?[J]. Nature, 2007, 445(7124): 159–160. DOI: 10.1038/445159a |
| [5] | Fu B B, Liu J W, Yang H M, et al. Shift of anammox bacterial community structure along the Pearl Estuary and the impact of environmental factors[J]. Journal of Geophysical Research:Oceans, 2015, 120(4): 2869–2883. DOI: 10.1002/2014JC010554 |
| [6] | Castro-González M, Molina V, Rodríguez-Rubio E, et al. The first report of a microdiverse anammox bacteria community in waters of Colombian Pacific, a transition area between prominent oxygen minimum zones of the eastern tropical Pacific[J]. Environmental Microbiology Reports, 2014, 6(6): 595–604. DOI: 10.1111/emi4.2014.6.issue-6 |
| [7] | Li M, Gu J D. Community structure and transcript responses of anammox bacteria, AOA, and AOB in mangrove sediment microcosms amended with ammonium and nitrite[J]. Applied Microbiology and Biotechnology, 2013, 97(22): 9859–9874. DOI: 10.1007/s00253-012-4683-y |
| [8] | Ward B B, Devol A H, Rich J J, et al. Denitrification as the dominant nitrogen loss process in the Arabian Sea[J]. Nature, 2009, 461(7260): 78–81. DOI: 10.1038/nature08276 |
| [9] | Shenzad A, Liu J W, Yu M, et al. Diversity, community composition and abundance of Anammox bacteria in sediments of the north Marginal seas of China[J]. Microbes and Environments, 2016, 31(2): 111–120. DOI: 10.1264/jsme2.ME15140 |
| [10] | Sonthiphand P, Hall M W, Neufeld J D. Biogeography of anaerobic ammonia-oxidizing (anammox) bacteria[J]. Frontiers in Microbiology, 2014, 5: 399. |
| [11] | 陆慧锋. 耐盐厌氧氨氧化工艺性能及其机理研究[D]. 杭州: 浙江大学, 2015. 5-6. |
| [12] | Li M, Hong Y G, Cao H L, et al. Community structures and distribution of anaerobic ammonium oxidizing and nirS-encoding nitrite-reducing bacteria in surface sediments of the South China Sea[J]. Microbial Ecology, 2013, 66(2): 281–296. DOI: 10.1007/s00248-012-0175-y |
| [13] | Dang H Y, Zhou H X, Zhang Z N, et al. Molecular detection of Candidatus scalindua pacifica and environmental responses of sediment anammox bacterial community in the Bohai Sea, China[J]. PLoS One, 2013, 8(4): e61330. DOI: 10.1371/journal.pone.0061330 |
| [14] | Kawagoshi Y, Nakamura Y, Kawashima H, et al. Enrichment of marine anammox bacteria from seawater-related samples and bacterial community study[J]. Water Science and Technology, 2010, 61(1): 119–126. DOI: 10.2166/wst.2010.796 |
| [15] | Kindaichi T, Awata T, Suzuki Y, et al. Enrichment using an up-flow column reactor and community structure of marine anammox bacteria from coastal sediment[J]. Microbes and Environments, 2011, 26(1): 67–73. DOI: 10.1264/jsme2.ME10158 |
| [16] | Kartal B, Koleva M, Arsov R, et al. Adaptation of a freshwater anammox population to high salinity wastewater[J]. Journal of Biotechnology, 2006, 126(4): 546–553. DOI: 10.1016/j.jbiotec.2006.05.012 |
| [17] | Yang J C, Zhang L, Hira D, et al. Anammox treatment of high-salinity wastewater at ambient temperature[J]. Bioresource Technology, 2011, 102(3): 2367–2372. DOI: 10.1016/j.biortech.2010.10.101 |
| [18] | Chen Y P, Ma T F, Hu X, et al. Start-up of a combined anaerobic/partial nitritation/ANAMMOX process for high-salt mustard wastewater treatment[J]. Applied Biochemistry and Biotechnology, 2015, 175(1): 119–134. DOI: 10.1007/s12010-014-1247-x |
| [19] | Wei Q Y, Kawagoshi Y, Huang X W, et al. Nitrogen removal properties in a continuous marine anammox bacteria reactor under rapid and extensive salinity changes[J]. Chemosphere, 2016, 148: 444–451. DOI: 10.1016/j.chemosphere.2016.01.041 |
| [20] | Nakajima J, Sakka M, Kimura T, et al. Enrichment of anammox bacteria from marine environment for the construction of a bioremediation reactor[J]. Applied Microbiology and Biotechnology, 2008, 77(5): 1159–1166. DOI: 10.1007/s00253-007-1247-7 |
| [21] | Thamdrup B, Dalsgaard T. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments[J]. Applied and Environmental Microbiology, 2002, 68(3): 1312–1318. DOI: 10.1128/AEM.68.3.1312-1318.2002 |
| [22] | Rysgaard S, Glud R N, Risgaard-Petersen N, et al. Denitrification and anammox activity in Arctic marine sediments[J]. Limnology and Oceanography, 2004, 49(5): 1493–1502. DOI: 10.4319/lo.2004.49.5.1493 |
| [23] | Huang X W, Wei Q Y, Urata K, et al. Kinetic study on nitrogen removal performance in marine anammox bacterial culture[J]. Journal of Bioscience and Bioengineering, 2014, 117(3): 285–291. DOI: 10.1016/j.jbiosc.2013.08.004 |
| [24] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [25] | Mu Y, Yu H Q, Wang G. A kinetic approach to anaerobic hydrogen-producing process[J]. Water Research, 2007, 41(5): 1152–1160. DOI: 10.1016/j.watres.2006.11.047 |
| [26] | Kawagoshi Y, Fujisaki K, Tomoshige Y, et al. Temperature effect on nitrogen removal performance and bacterial community in culture of marine anammox bacteria derived from sea-based waste disposal site[J]. Journal of Bioscience and Bioengineering, 2012, 113(4): 515–520. DOI: 10.1016/j.jbiosc.2011.11.024 |
| [27] | 于德爽, 魏思佳, 李津, 等. 温度对厌氧氨氧化与反硝化耦合脱氮除碳的影响[J]. 中国环境科学, 2016, 36(5): 1384–1391. Yu E S, Wei S J, Li J, et al. Effect of temperature on simultaneous carbon and nitrogen removal by anaerobic ammonium oxidation and denitrification[J]. China Environmental Science, 2016, 36(5): 1384–1391. |
| [28] | 李祥, 黄勇, 郑宇慧, 等. 温度对厌氧氨氧化反应器脱氮效能稳定性的影响[J]. 环境科学, 2012, 33(4): 1288–1292. Li X, Huang Y, Zheng Y H, et al. Effect of temperature on stability of nitrogen removal in the ANAMMOX reactor[J]. Environmental Science, 2012, 33(4): 1288–1292. |
| [29] | Isaka K, Date Y, Kimuar Y, et al. Nitrogen removal performance using anaerobic ammonium oxidation at low temperatures[J]. FEMS Microbiology Letters, 2008, 282(1): 32–38. DOI: 10.1111/fml.2008.282.issue-1 |
| [30] | Lotti T, Kleerebezem R, van Loosdrecht M C. Effect of temperature change on anammox activity[J]. Biotechnology and Bioengineering, 2015, 112(1): 98–103. DOI: 10.1002/bit.25333 |
| [31] | Strous M, Kuenen J G, Jetten M S M. Key physiology of anaerobic ammonium oxidation[J]. Applied and Environmental Microbiology, 1999, 65(7): 3248–3250. |
| [32] | Tang C J, Zheng P, Chai L Y, et al. Thermodynamic and kinetic investigation of anaerobic bioprocesses on ANAMMOX under high organic conditions[J]. Chemical Engineering Journal, 2013, 230: 149–157. DOI: 10.1016/j.cej.2013.06.047 |