 2017, Vol. 38
2017, Vol. 38



污水处理厂广泛采用生物脱氮技术,因为硝化细菌对许多环境因素比较敏感(如pH、 温度、 溶解氧浓度等),所以脱氮过程中的硝化作用被认为是整个脱氮过程的限速步骤[1]. 硝化过程中氨氧化细菌(ammonia-oxidizing bacteria,AOB)将氨氮(NH4+-N) 转化为亚硝酸盐氮(NO2--N),然后亚硝酸盐氧化细菌(nitrite-oxidizing bacteria,NOB)将亚硝酸盐氮(NO2--N)转化为硝酸盐氮(NO3--N)[2, 3]. 污水中的氨氮以游离氨(NH3)和铵根离子(NH4+)的形式存在,游离氨(free ammonia,FA)是AOB菌群生长所需要的初始基质,但是过高的FA会对其产生毒害作用[4, 5],影响AOB和NOB菌群的生长[6, 7]. 王淑莹等[8]研究了高浓度FA对絮状污泥和好氧颗粒污泥的抑制影响,发现氨氧化菌均匀分布的絮状污泥容易受到FA抑制. Chen等[9]在富集高效硝化菌的过程中发现当FA浓度增加到10~20 mg·L-1(混合反应器),FA对反应器的脱氮性能造成明显的抑制. 尤永军[10]在SBR反应器中研究FA对硝化菌的影响时,指出随FA浓度提高,AOB活性逐渐增大而NOB活性先增大后被抑制. 彭赵旭等[11]研究了在活性污泥系统中(序批式活性污泥反应器,SBR),当进水NH4+-N为65 mg·L-1时[混合液悬浮固体浓度(mixed liquid suspended solids,MLSS)为2 200~2 400 mg·L-1],反应器硝化效果明显变差,NH4+-N冲击负荷对硝化作用造成短期抑制. 目前已有的研究主要集中在FA浓度对硝化性能和活性的短期抑制影响方面,或是以亚硝酸盐积累为目的短程硝化作用影响的研究[12~14],例如孙洪伟等[15]充分利用较高FA对NOB的抑制,通过优化DO、 ORP、 pH等参数,准确实现短程硝化. 但对生物系统反复受到高浓度氨氮废水的急性冲击负荷(以下称“冲击负荷”),以及完整的生物硝化反应的抑制与活性恢复机制的研究还不够. 在连续运行模式下,不同FA浓度对硝化作用冲击的浓度特点、 抑制程度、 持续时间、 微生物菌群变化等相关信息还缺乏报道.
本文采用SBR反应器处理模拟高浓度氨氮污水,研究高氨氮废水冲击负荷条件下,FA对生物硝化作用的抑制和细菌活性恢复的影响、 硝化菌群(AOB和NOB菌群)结构的变化,以揭示高浓度FA对生物硝化过程的影响机制.
1 材料与方法 1.1 试验装置试验所采用的SBR反应器试验系统如图 1所示,SBR反应器有效容积5 L,总高0.85 m. 反应器依靠保温水层维持温度在25℃±0.5℃; 底部曝气,控制反应器内溶解氧(dissolved oxygen,DO)≥2 mg·L-1. SBR反应器自动运行,每个周期6 h(进水5 min,曝气4 h,静沉75 min,排水10 min,静置30 min),每周期出水量为2.5 L. 定期排泥,控制MLSS为6 000 mg·L-1左右,污泥龄约为10 d.

|
图 1 SBR反应器试验系统示意 Fig. 1 Schematic of SBR experiment system |
SBR反应器进水采用模拟生活污水,利用无水葡萄糖配置进水并控制化学需氧量(COD)为250 mg·L-1,利用氯化铵配置进水NH4+-N为50 mg·L-1,pH为7.8左右,具体组分为(mg·L-1):无水葡萄糖240,NH4Cl 320,KH2PO4 30,NaHCO3 350,MgSO4·7H2O 20,FeSO4·7H2O 2.5,ZnSO4·7H2O 0.25,CaCl2·2H2O 10,CoCl2·6H2O 0.000 05,MoO3 0.001 5.
1.3 试验方法从天津某城市污水厂曝气池取得接种活性污泥,用2 mm孔径的滤网去除污泥中的杂质,然后放入SBR反应器进行驯化培养. 在进水氨氮浓度为50 mg·L-1条件下,运行SBR反应器,待出水NH4+-N稳定至0.2 mg·L-1以下时,开始进行冲击负荷试验. 冲击负荷试验中,SBR的进水NH4+-N浓度逐渐提高到150、 200、 300、 400、 500、 750、 1 000、 1 250、 1 500、 2 000 mg·L-1,每一个冲击负荷运行一个周期,然后进水NH4+-N立刻恢复到50 mg·L-1进行自然恢复过程,试验中NH4+-N稳定至0.2 mg·L-1以下所需的自然恢复时间分别为 2、 3、 4、 5、 6、 7、 9、 9、 9、 12个周期. 在冲击负荷周期中取曝气5 min、 1 h、 2 h、 3 h、 4 h时的活性污泥混合样以及静沉后的出水(定义为第6 h样品),经0.45 μm滤膜过滤后检测NH4+-N、 NO2--N、 NO3--N等指标. 在冲击负荷周期曝气4 h时取活性污泥混合液进行比耗氧速率(SOUR)和荧光原位杂交(FISH)分析.
1.4 生物活性的表征AOB、 NOB及异养菌的活性分别用氨氧化比耗氧速率(氨氧化SOUR)、 亚硝酸盐氮氧化比耗氧速率(硝化SOUR)和碳氧化比耗氧速率(碳氧化SOUR)来表征. 通过添加不同浓度的抑制选择剂[氯酸钠(NaClO3)和丙烯基硫脲(ATU)][16],可以测出氨氧化、 亚硝酸盐氮氧化和碳氧化比耗氧速率.
1.5 其它分析方法COD:密封消解和哈希COD测定仪测定; NH4+-N:纳氏试剂分光光度法; NO3--N:酚二磺酸分光光度度法; NO2--N:N-(1-萘基)-乙二胺分光光度度法; MLSS:重量法; DO和pH:仪器测定; SOUR:参见文献[16]; FISH:样品固定和杂交方法依据标准杂交培育方法[17],用两种寡核苷酸探针NSO1225和Ntspa662分别标记AOB菌[18]和NOB菌[19],并用OLYMPUS IX71(Olympus,Japan)荧光显微镜观察已制备好的玻片,分析杂交结果.
1.6 游离氨的浓度由于NH3与NH4+-N之间存在着电离平衡,因此进水FA的浓度与进水NH4+-N、 pH以及温度有着密切的关系[20],见式(1)和式(2)所示:
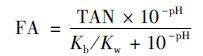
|
(1) |
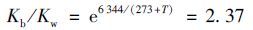
|
(2) |
式中,FA为游离氨浓度(mg·L-1); TAN为总氨氮浓度(mg·L-1); Kb 为与温度有关的氨电离常数; Kw 为与温度有关的水电离常数.
在冲击负荷试验中,当进水NH4+-N浓度为:150、 200、 300、 400、 500、 750、 1 000、 1 250、 1 500和2 000 mg·L-1时的pH分别为7.72、 7.44、 7.33、 7.32、 7.27、 7.19、 7.15、 7.10、 7.04和6.98(维持在中性左右),依上述公式计算得到,冲击负荷进水FA的浓度分别为:4.5、 3.6、 4.1、 5.0、 5.4、 6.6、 8.1、 9.3、 10.5和11.8 mg·L-1.
1.7 抑制率计算本文采用氨氮的最大比降解速率抑制率(IPA)和硝酸盐氮的最大比生成速率抑制率(IPN)[21]表征FA分别对AOB和NOB的抑制程度,如公式(3)和(4)所示:
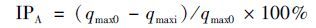
|
(3) |
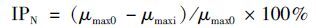
|
(4) |
式中,qmax0为 FA冲击负荷试验中最大的氨氮最大比降解速率(以N/MLSS计,下同),本试验中,qmax0=12.73 mg·(g·h)-1; qmaxi为FA冲击负荷试验中,不同FA对应的氨氮最大比降解速率; μmax0为FA冲击负荷试验中最大的硝酸盐氮最大比生成速率(以NO3--N/MLSS计,下同),本试验中,μmax0=4.26 mg·(g·h)-1; μmaxi为FA冲击负荷试验中的硝酸盐氮最大比生成速率.
2 结果与讨论 2.1 驯化污泥的活性表征驯化阶段进水NH4+-N浓度范围为49.5~55.1 mg·L-1,出水NH4+-N均低于0.2 mg·L-1,NH4+-N去除率可达99.7% 以上,说明反应器中硝化细菌活性很高. 进水COD为250 mg·L-1左右,驯化期间出水COD可降至20 mg·L-1以下,COD去除率在91.6% 以上. 出水pH稳定在7.2~7.5的范围内. 图 2显示一个曝气周期中氮的形态转化,从中可知,在曝气的前4 h,NH4+-N浓度逐渐降低到0.01 mg·L-1,同时出水NO3--N浓度逐渐增加,最后稳定在32.3 mg·L-1左右,氨氮最大比降解速率为1.38 mg·(g·h)-1. NO2--N从曝气开始浓度逐渐增加,在1 h左右达到最大值16.0 mg·L-1,随后逐渐降低,曝气4 h后NO2--N低于检出限. 通过氮平衡的计算,88.6%的NH4+-N转化成了NO3--N,其它部分的NH4+-N可能是通过细菌的合成代谢被转化为有机氮[22].

|
图 2 SBR典型曝气周期中氮的形态转化 Fig. 2 Transformation of the nitrogen forms in a typical aeration circle |
因为SBR系统的活性污泥中存在大量的异养菌,异养菌会利用一部分NH4+-N进行自身合成代谢,因此进水NH4+-N的去除是由异养菌和硝化细菌共同来完成的. 在一定的DO和碱度条件下,异养菌能够利用有机物和NH4+-N进行合成代谢[根据合成代谢公式(5)],本试验驯化过程中进水葡萄糖浓度约为250 mg·L-1,出水COD为20 mg·L-1左右,则理论上异养菌合成代谢消耗的NH4+-N量为15.3 mg·L-1,接近实际进水和出水中总无机氮浓度之差,即20.0 mg·L-1,剩余误差可能是由于SBR中存在反硝化等反应造成的[23].
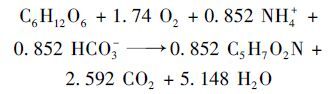
|
(5) |
在冲击负荷试验中,SBR反应器的进水NH4+-N浓度逐渐升高[图 3(a)],每一个冲击负荷运行一个周期,然后进水NH4+-N立刻恢复到50 mg·L-1进行自然恢复过程[图 3(a)]. 当NH4+-N冲击负荷为150 mg·L-1(FA=4.5 mg·L-1)时,在第一个周期运行结束时,出水NH4+-N浓度为26.6 mg·L-1[图 3(b)],说明在这个NH4+-N浓度下系统内的AOB菌群已经不能对进水的NH4+-N进行全部去除,经过2个恢复周期,出水NH4+-N才降至0.2 mg·L-1以下水平. 随着NH4+-N浓度的增加,短时出水NH4+-N浓度也逐渐增加,且出水NH4+-N恢复至0.2 mg·L-1以下所需的周期逐渐增加,当 NH4+-N冲击负荷为2 000 mg·L-1(FA=3.6 mg·L-1)时,需要12个周期才能完全恢复[图 3(b)]. 可见进水氨氮浓度越大,其短时去除率越低,出水NH4+-N降至0.2 mg·L-1以下的恢复时间越长. 虽然恢复时间增加了,但是FA 对AOB和NOB的抑制作用在试验浓度下是可恢复的[24],且硝化菌的活性并不能被完全抑制[25]. 张亮等[26]指出FA对硝化菌的抑制都是可逆的,硝化菌受FA抑制解除后,可逐渐恢复正常活性,且硝化菌对游离氨的抑制存在适应性,长期高游离氨环境下的硝化菌活性也可能逐渐恢复. 相比于重金属类污染物(反应器相同)对活性污泥系统的毒性抑制作用[27],FA冲击负荷对活性污泥的恢复速度相对较快.

|
图 3 NH4+-N抑制及其恢复阶段的运行效果 Fig. 3 Performance of the SBR in the stages of NH4+-N inhibition and recovery |
在150~500 mg·L-1 的NH4+-N冲击负荷试验中(即进水FA浓度为3.6~5.4 mg·L-1),出水中几乎没有NO2--N的积累[图 3(c)],而当NH4+-N投加浓度大于500 mg·L-1(即FA浓度大于5.4 mg·L-1),出水NO2--N出现明显的积累,最大积累量为2.4 mg·L-1. 当NH4+-N大于750 mg·L-1(即FA浓度大于6.6 mg·L-1)的冲击负荷结束时,出水NO2--N浓度分别为3.9、 26.3、 17.0、 24.3和38.4 mg·L-1,说明NH4+-N冲击浓度大于等于750 mg·L-1(FA浓度6.6 mg·L-1)时,NOB开始受到FA的抑制. 并且,从图 3可以明显看出,NOB相对于AOB较易受FA冲击负荷的影响.
2.3 FA冲击负荷对硝化速率的影响根据各个冲击负荷曝气阶段的NH4+-N、 NO2--N和NO3--N浓度变化,得到不同FA冲击负荷下氨氮去除量[图 4(a)]和氨氮最大比降解速率[图 4(b)]. 随着进水FA浓度的增加,SBR反应器对NH4+-N去除量以及氨氮最大比降解速率呈现先升高后降低的趋势,当FA浓度为9.3 mg·L-1时,NH4+-N去除量最大为156.5 mg·L-1,是驯化阶段NH4+-N去除量的3.4倍; FA浓度增加至11.8 mg·L-1时,NH4+-N去除量又下降至113.0 mg·L-1,说明3.6~9.3 mg·L-1的FA促进AOB菌群对NH4+-N的去除,而FA浓度大于9.3 mg·L-1时抑制AOB菌群对NH4+-N的去除.

|
图 4 SBR系统不同FA冲击负荷下氨氮去除与硝酸盐氮生成速率 Fig. 4 Consumption rates of NH4+-N and formation rates of NO3--N under different FA loading concentrations in SBR system |
出水NO2--N的浓度也随着FA的增加逐渐增大. FA浓度低于6.6 mg·L-1时,系统中NO2--N浓度较低,最高浓度为1.2 mg·L-1; 当FA增加至11.8 mg·L-1时,系统中NO2--N发生明显积累,出水NO2--N浓度达到38.4 mg·L-1,说明NOB菌群受到FA的严重抑制,NO2--N积累量增加了96.5%,与Chen等[9]在活性污泥反应器(CSTR)的试验得到的FA在11.3 mg·L-1时,NO2--N积累量增加了93.8% 相近. 试验出水NO3--N的浓度以及硝酸盐氮最大比生成速率呈现先升高后降低的变化趋势,与NH4+-N去除量的变化趋势相一致,且硝酸盐氮最大比生成速率的最大值出现在FA浓度为10.5 mg·L-1时,为4.26 mg·(g·h)-1. 当FA为11.8 mg·L-1时,硝酸盐氮最大比生成速率降至1.73 mg·(g·h)-1,NO3--N生成量的降低反映了FA对NOB菌群的抑制作用. Chung等[28]发现在FA逐渐升高到10 mg·L-1时氨氮比降解速率逐渐增至最大,大于10 mg·L-1时氨氮比降解速率开始降低; FA为5 mg·L-1 时亚硝酸氧化作用的速率达到最大,随后降低,该研究的峰值与本文的研究结论相似.
随着FA浓度的增大,氨氮的最大比降解速率的抑制率IPA=80.70%,硝酸盐氮的最大比生成速率的抑制率IPN=59.46%,IPA大于IPN,说明NOB比AOB菌群更敏感,更易受到高浓度FA的抑制作用.
2.4 FA冲击负荷对SOUR的影响图 5显示了FA冲击周期内曝气阶段的亚硝化SOUR(表征AOB菌群的活性)、 硝化SOUR(表征NOB菌群的活性)和碳氧化SOUR(表征异养菌的活性). 随着进水FA浓度从3.6 mg·L-1增加到8.1 mg·L-1,亚硝化SOUR呈现增加的趋势. FA浓度为8.1 mg·L-1时,亚硝化SOUR达到最大,即0.20 mg·(g·min)-1(以O2/MLSS计,下同),之后随着FA浓度的继续升高(大于8.1 mg·L-1),亚硝化SOUR急剧下降至0.06 mg·(g·min)-1,FA对亚硝化SOUR的抑制率为70%. 此抑制结果与前述最大比降解速率的峰值结果一致(图 4). 硝化SOUR的变化趋势与亚硝化SOUR的变化趋势相同,当FA浓度为6.6 mg·L-1时,硝化SOUR达到最大值0.40 mg·(g·min)-1(以O2/MLSS计,下同),随后硝化SOUR急剧降低至0.10 mg·(g·min)-1,FA对硝化SOUR的抑制率达到75%. 亚硝化SOUR比硝化SOUR达到峰值时的FA浓度高,说明AOB比NOB菌群抗FA抑制的能力更强. 此外,FA对异养菌的SOUR基本没有太明显的影响,表明FA冲击负荷对异养菌的活性没有明显的抑制作用.

|
图 5 不同FA冲击负荷下系统微生物活性的变化 Fig. 5 Microbial activities under different FA loading concentrations |
氨氮既是作为基质被微生物利用,也是作为去除标准来评价冲击负荷对硝化作用的影响,随着进水FA浓度从3.6 mg·L-1增加到8.1 mg·L-1,氨氮的去除率虽然降低并且恢复时间增加[图 3(b)],但亚硝化SOUR呈现上升的趋势(图 5),即AOB菌群的活性不断升高. 系统受该浓度范围的FA冲击负荷后,出水氨氮出现积累,可见进水氨氮并不能完全被微生物代谢利用,说明氨氮投加量大于硝化菌自身代谢的量,才会出现出水氨氮的积累. 当进水氨氮浓度继续提高时才会对硝化菌活性造成活性抑制作用(图 5). 尤永军[10]在SBR反应器中研究FA浓度(0.5、 5、 10 mg·L-1)对硝化菌的影响时,指出随FA浓度提高,AOB活性逐渐增大,NOB活性先增大后被抑制(10 mg·L-1). 由此可见,FA抑制条件行下,系统氨氮去除率的下降并不一定表示菌群活性的降低.
2.5 FISH结果分析本试验在同一个反应器中经过连续的不同负荷的冲击及其恢复过程的运行(161周期),活性污泥中的硝化菌群有足够时间发生变化. 因此,对驯化后以及冲击负荷曝气4 h的活性污泥混合液进行FISH分析观察菌群分布的变化特征,用荧光显微镜观察已制备好的玻片,得到泥样中总细菌、 AOB和NOB细菌的分布比例(图 6和图 7). 观察发现,驯化后的原污泥样中AOB和NOB菌群所占比例分别为13.7%和9.0%(图 6),进水受到3.6 mg·L-1的FA冲击负荷后,AOB和NOB菌群所占比例分别为14.5%和9.7%(图 7). 对不同冲击负荷结束后的FISH图进行数据化分析(图 8),结果表明,在FA的浓度小于8.1 mg·L-1时,随着进水FA浓度不断升高,冲击负荷结束后AOB菌群的数量相对于前一个冲击负荷有略微升高的趋势,在FA的浓度大于 8.1 mg·L-1时,冲击负荷结束后AOB菌群的数量相对于前一个冲击负荷有略微下降的趋势,在11.8 mg·L-1冲击负荷结束时,AOB菌群数量降至最低. NOB菌群的数量也有相同趋势(FA浓度在3.6~5.4 mg·L-1阶段增加,大于5.4 mg·L-1开始降低,图 8). 当FA冲击负荷的浓度小于5.4 mg·L-1时,不同冲击负荷结束后,AOB菌群的丰度基本保持不变,但是AOB菌群的活性升高(图 5 中硝化SOUR),NH4+-N的去除量也在不断增加,说明这个范围内的FA浓度提高了AOB菌群的活性,而对AOB菌群的丰度并没有太大影响; 在FA冲击负荷为5.4~8.1 mg·L-1的范围时,AOB菌群的丰度和活性均有上升趋势,说明这个浓度范围的FA不仅增强AOB菌群的活性,而且可以促进AOB菌群的繁殖; 当FA浓度大于8.1 mg·L-1时,AOB菌群受到了明显的抑制,丰度逐渐下降,且NH4+-N的去除量明显下降,表明了高浓度FA对AOB菌群造成了毒害作用. Villaverde等[29]的研究中,在生物过滤装置中,利用较低的不同浓度的FA对硝化菌进行驯化,发现硝化菌对不同浓度的FA有很好的适应性,可能是由于硝化菌的逐渐增殖或者是硝化菌本身的活性增加. 在整个FA冲击负荷过程中,AOB菌群始终是优势菌群(图 8),且AOB和NOB菌群的比例基本不变,这与彭赵旭等[11]提出的氨氮冲击负荷对硝化作用的短期影响中,AOB和NOB菌群比例不变的发现相一致.

|
图 6 原泥样中硝化菌群FISH图片 Fig. 6 FISH assay pictures of bacteria flora in original sample |

|
图 7 FA冲击负荷下(3.6 mg·L-1)硝化菌群FISH图片 Fig. 7 FISH assay pictures of bacteria flora in the sample after the shock loading of FA (3.6 mg·L-1) |
本试验中将生物硝化过程(氨氧化过程和亚硝酸盐氮氧化过程)开始受FA抑制的浓度定义为临界抑制浓度,用来表征AOB和NOB菌群等硝化菌群对不同进水条件下的高浓度FA的耐受性. 临界抑制浓度由菌群丰度变化(FISH)以及活性变化(SOUR)共同决定. 当FA小于等于临界抑制浓度时,FA对硝化细菌的硝化能力有一定促进作用; 反之,当FA大于临界抑制浓度时,FA开始对硝化细菌活性和丰度造成抑制作用. 在试验的冲击负荷条件下,AOB菌群的FA临界抑制浓度为8.1 mg·L-1,而NOB菌群的FA临界抑制浓度为6.6 mg·L-1,说明NOB 比 AOB 菌群对FA冲击负荷影响更敏感,更易受到抑制作用.

|
图 8 不同FA冲击负荷下AOB和NOB菌群分布比例 Fig. 8 Relative proportions of AOB and NOB population under different FA loading concentrations |
(1) 本试验表明FA冲击负荷在小于8.1 mg·L-1的范围内时,增加FA负荷能够促进硝化活性,而当FA冲击负荷大于8.1 mg·L-1时,会对硝化作用造成抑制. AOB和NOB菌群活性的增强,体现在菌群数量增加和比耗氧速率SOUR增加; AOB和NOB菌群活性减弱,体现在菌群数量减少和比耗氧速率SOUR降低.
(2) FA的冲击负荷不改变AOB和NOB菌群的菌群比例,AOB和NOB菌群的丰度在进水FA浓度为3.6~8.1 mg·L-1时都略微升高,当FA浓度大于8.1 mg·L-1时都会受到抑制,菌群丰度明显降低.
(3) FA对硝化作用的抑制体现在对AOB和NOB菌群的丰度和活性(SOUR)的共同抑制,且菌群丰度和活性受抑制的变化趋势基本一致; 氨氮去除率的变化与菌群的活性变化情况并不一致,当基质远远过量时,活性的提高仍然不能避免基质去除率的降低.
(4) FA冲击负荷对硝化作用的抑制作用是可恢复的,硝化菌活性恢复时间随着FA浓度的升高而延长. 较低浓度时(FA=3.6 mg·L-1),2个周期即可恢复,较高浓度时(FA=11.8 mg·L-1),则需要12个周期才能恢复.
| [1] | Hu Z Q, Chandran K, Grasso D, et al. Effect of nickel and cadmium speciation on nitrification inhibition[J]. Environmental Science &Technology, 2002, 36(14) : 3074–3078. |
| [2] | Leyva-Díaz J C, González-Martínez A, González-López J, et al. Kinetic modeling and microbiological study of two-step nitrification in a membrane bioreactor and hybrid moving bed biofilm reactor-membrane bioreactor for wastewater treatment[J]. Chemical Engineering Journal, 2015, 259 : 692–702. DOI: 10.1016/j.cej.2014.07.136 |
| [3] | Cho K, Shin S G, Lee J, et al. Nitrification resilience and community dynamics of ammonia-oxidizing bacteria with respect to ammonia loading shock in a nitrification reactor treating steel wastewater[J]. Journal of Bioscience and Bioengineering, 2016, 122(2) : 196–202. DOI: 10.1016/j.jbiosc.2016.01.009 |
| [4] | Puigagut J, Salvadó H, García J. Short-term harmful effects of ammonia nitrogen on activated sludge microfauna[J]. Water Research, 2005, 39(18) : 4397–4404. DOI: 10.1016/j.watres.2005.08.008 |
| [5] | 刘宏波, 杨昌柱, 濮文虹, 等. 进水氨氮浓度对好氧颗粒污泥的影响研究[J]. 环境科学, 2009, 30(7) : 2030–2034. Liu H B, Yang C Z, Pu W H, et al. Effect of ammonia concentration on the characteristic of aerobic granular sludge[J]. Environmental Science, 2009, 30(7) : 2030–2034. |
| [6] | Kim J H, Guo X J, Park H S. Comparison study of the effects of temperature and free ammonia concentration on nitrification and nitrite accumulation[J]. Process Biochemistry, 2008, 43(2) : 154–160. DOI: 10.1016/j.procbio.2007.11.005 |
| [7] | Vadivelu V M, Keller J, Yuan Z. Free ammonia and free nitrous acid inhibition on the anabolic and catabolic processes of Nitrosomonas and Nitrobacter[J]. Water Science and Technology, 2007, 56(7) : 89–97. DOI: 10.2166/wst.2007.612 |
| [8] | 王淑莹, 刘旭, 吴蕾. 温度和游离氨对颗粒与絮状污泥硝化性能影响的对比研究[J]. 北京工业大学学报, 2013, 39(2) : 297–303. Wang S Y, Liu X, Wu L. Effects of temperature and free ammonia concentration on nitrification between the aerobic granular and the floccular sludges[J]. Journal of Beijing University of Technology, 2013, 39(2) : 297–303. |
| [9] | Chen J W, Zheng P, Yu Y, et al. Enrichment of high activity nitrifers to enhance partial nitrification process[J]. Bioresource Technology, 2010, 101(19) : 7293–7298. DOI: 10.1016/j.biortech.2010.04.080 |
| [10] | 尤永军. 游离氨(FA)对硝化菌活性的抑制影响试验研究[D]. 兰州:兰州交通大学, 2015. 25-29. You Y J. Study on the effect of free ammonia (FA) on the inhibition of nitrifying bacteria activity[D]. Lanzhou:Lanzhou Jiaotong University, 2015. 25-29. |
| [11] | 彭赵旭, 彭永臻, 桂丽娟, 等. 氨氮冲击负荷对硝化过程的短期影响[J]. 中国给水排水, 2010, 26(11) : 9–12. Peng Z X, Peng Y Z, Gui L J, et al. Short-term influence of ammonia nitrogen impact load on nitrification process[J]. China Water &Wastewater, 2010, 26(11) : 9–12. |
| [12] | 吴莉娜, 彭永臻, 王淑莹, 等. 游离氨对城市生活垃圾渗滤液短程硝化的影响[J]. 环境科学, 2008, 29(12) : 3428–3432. Wu L N, Peng Y Z, Wang S Y, et al. Effect of free ammonia on the short-cut nitrification of the municipal landfill leachate[J]. Environmental Science, 2008, 29(12) : 3428–3432. |
| [13] | 孙洪伟, 尤永军, 白宝霞, 等. 游离氨对硝化菌活性的短期抑制影响试验研究[J]. 中国给水排水, 2015, 31(3) : 19–23. Sun H W, You Y J, Bai B X, et al. Study on short-term inhibition of free ammonia on activity of nitrifying bacteria[J]. China Water &Wastewater, 2015, 31(3) : 19–23. |
| [14] | 孙洪伟, 吕心涛, 魏雪芬, 等. 游离氨(FA)耦合曝气时间对硝化菌活性的抑制影响[J]. 环境科学, 2016, 37(3) : 1075–1081. Sun H W, Lü X T, Wei X F, et al. Synergetic inhibitory effect of free ammonia and aeration phase length control on the activity of nitrifying bacteria[J]. Environmental Science, 2016, 37(3) : 1075–1081. |
| [15] | 孙洪伟, 杨庆, 董国日, 等. 游离氨抑制协同过程控制实现渗滤液短程硝化[J]. 中国科学:化学, 2010, 40(8) : 1156–1162. Sun H W, Yang Q, Dong G R, et al. Achieving the nitrite pathway using FA inhibition and process control in UASB-SBR removing nitrogen from landfill leachate[J]. Scientia Sinica Chimica, 2010, 40(8) : 1156–1162. |
| [16] | 王建龙, 吴立波, 齐星, 等. 用氧吸收速率(OUR)表征活性污泥硝化活性的研究[J]. 环境科学学报, 1999, 19(3) : 225–229. Wang J L, Wu L B, Qi X, et al. Characterization of nitrification activity of activated sludge by oxygen uptake rate (OUR)[J]. Acta Scientiae Circumstantiae, 1999, 19(3) : 225–229. |
| [17] | Yusof N, Hassan M A, Yee P L, et al. Nitrification of high-strength ammonium landfill leachate with microbial community analysis using fluorescence in situ hybridization (FISH)[J]. Waste Management &Research, 2011, 29(6) : 602–611. |
| [18] | Degrange V, Bardin R. Detection and counting of Nitrobacter populations in soil by PCR[J]. Applied and Environmental Microbiology, 1995, 61(6) : 2093–2098. |
| [19] | Han D W, Chang J S, Kim D J. Nitrifying microbial community analysis of nitrite accumulating biofilm reactor by fluorescence in situ hybridization[J]. Water Science and Technology, 2003, 47(1) : 97–104. |
| [20] | Anthonisen A C, Loehr R C, Prakasam T B S, et al. Inhibition of nitrification by ammonia and nitrous acid[J]. Journal (Water Pollution Control Federation), 1976, 48(5) : 835–852. |
| [21] | Park S, Bae W. Modeling kinetics of ammonium oxidation and nitrite oxidation under simultaneous inhibition by free ammonia and free nitrous acid[J]. Process Biochemistry, 2009, 44(6) : 631–640. DOI: 10.1016/j.procbio.2009.02.002 |
| [22] | Bruce E R, Perry L M C. Environmental biotechnology:principles and applications[M]. New York: MeGraw Hill Publishing Co, 2001. |
| [23] | 许德超, 陈洪波, 李小明, 等. 静置/好氧/缺氧序批式反应器(SBR)脱氮除磷效果研究[J]. 环境科学学报, 2014, 34(1) : 152–159. Xu D C, Chen H B, Li X M, et al. Biological phosphorus and nitrogen removal in a sequencing batch reactor (SBR) operated in static/aerobic/anoxic regime[J]. Acta Scientiae Circumstantiae, 2014, 34(1) : 152–159. |
| [24] | 孙洪伟, 尤永军, 赵华南, 等. 游离氨对硝化菌活性的抑制及可逆性影响[J]. 中国环境科学, 2015, 35(1) : 95–100. Sun H W, You Y J, Zhao H N, et al. Inhibitory effect of free ammonia on the activity of nitrifying bacteria and recoverability[J]. China Environmental Science, 2015, 35(1) : 95–100. |
| [25] | 魏琛, 罗固源. 游离氨对稳定生物亚硝化的影响分析[J]. 重庆环境科学, 2003, 25(12) : 50–52. Wei C, Luo G Y. The influence of free ammonium on stable short-cut nitritying system[J]. Chongqing Environmental Science, 2003, 25(12) : 50–52. |
| [26] | 张亮, 张树军, 彭永臻. 污水处理中游离氨对硝化作用抑制影响研究[J]. 哈尔滨工业大学学报, 2012, 44(2) : 75–79. Zhang L, Zhang S J, Peng Y Z. Review of study on the effects of free ammonia inhibitions on nitrifying bacteria in wastewater treatment[J]. Journal of Harbin Institute of Technology, 2012, 44(2) : 75–79. |
| [27] | 王越, 翟洪艳, 季民, 等. Cr(Ⅵ)对活性污泥硝化活性的抑制及在污泥中的分布特点[J]. 环境科学学报, 2015, 35(5) : 1435–1443. Wang Y, Zhai H Y, Ji M, et al. Inhibitory effects of Cr(Ⅵ) on nitrification activity and Cr(Ⅵ) distribution in activated sludge[J]. Acta Scientiae Circumstantiae, 2015, 35(5) : 1435–1443. |
| [28] | Chung J, Shim H, Park S J, et al. Optimization of free ammonia concentration for nitrite accumulation in shortcut biological nitrogen removal process[J]. Bioprocess and Biosystems Engineering, 2006, 28(4) : 275–282. DOI: 10.1007/s00449-005-0035-y |
| [29] | Villaverde S, Fdz-Polanco F, García P A. Nitrifying biofilm acclimation to free ammonia in submerged biofilters. Start-up influence[J]. Water Research, 2000, 34(2) : 602–610. DOI: 10.1016/S0043-1354(99)00175-X |