 2016, Vol. 37
2016, Vol. 37



贡湖是太湖的重要组成部分,是无锡苏州两市的饮用水水源地和太湖鱼类重要的产卵地与索饵地,在水资源及生物多样性等方面具有重要作用,然而由于太湖富营养化水平的持续居高不下,加之周边城市的快速扩张,贡湖流域面临水生态系统结构单一,污染净化能力下降等问题,水质总体处于Ⅴ类. 而入太湖贡湖河道水体污染,水体氮素含量的升高,严重影响到太湖贡湖水生生态系统的生物多样性及稳定性[1]. 因此,控制入贡湖河道污染,改善入湖河道水体水质,对于水源地水体保护以及保护河道水环境的生态多样性具有重要意义.
目前,国内外学者对于河道氮素去除,水体富营养化控制以及脱氮机制等方面进行了广泛的研究. 蒋跃等[2]研究了美人蕉(Canna indica Linn.)、 再力花(Thalia dealbata)、 千屈菜(Lythrum salicaria)这3种浮床植物的生长特性以及其对氮、 磷吸收的优化配置[2]. 李正魁等[3]探讨了应用氮循环菌的方法对湖泊水体氮污染的修复,陈祈春等[4]研究了沉水植物床-固定化微生物技术在水源地中的修复效果. 上述技术相比较挺水与沉水植物浮床系统,微曝气生态浮床[5]等技术,在控制水体的富营养化方面具有较为显著的效果[6, 7]. 然而上述研究主要针对水体氮素的去除效果,而对水生植物-微生物具体如何影响入湖河道底泥中的脱氮微生物机制的研究尚不多见[8, 9]. 因此,本研究针对入贡湖亲水河氮素负荷较高、 河道水体流速缓慢、 水生植物逐渐死亡等特点,选取了脱氮微生物(immobilized nitrogen cycling bacteria,INCB),与伊乐藻种(Elodea nuttallii)相结合,进行模拟实验,分析原位亲水河的水体修复,通过提高亲水河氮素转化能力达到河道水体修复的目的. 同时本研究还探讨了伊乐藻与高效脱氮微生物联用下,对于河道泥水界面中脱氮微生物群落的影响,分析了河道氮素脱除的机制.
1 材料与方法 1.1 研究区域概况研究区域主要位于入贡湖亲水河(图 1),其位 于太湖新城南部、 环太湖高速公路北侧,东起蠡河 西,西至壬子港,全长13.82 km. 沟通环太湖大堤北侧的大量断头浜,在改善太湖新城南部水环境中发挥着重要的作用. 亲水河是贡湖入太湖前的缓冲河流,拦截地表径流及各类污染物质进入贡湖水域. 根据2011-10-2013-03的长期水质跟踪监测资料,贡湖湾亲水河区域生态环境严重退化,污染负荷不断增加,流速减慢,河道的自净能力降低,水质恶化,其中总磷(TP)0.1-0.2 mg ·L-1,总氮(TN) 4-5 mg ·L-1,氨氮(NH4+-N)2-3 mg ·L-1,硝氮(NO3--N)0.5-1.0 mg ·L-1,总体为劣Ⅴ类水. 具体数据见表 1.
 | 图 1 太湖贡湖湾入湖河道(亲水河)实验修复地点 Fig. 1 Test remediation site of inflow river (Qinshui River) in Gonghu Bay,Taihu Lake |
|
|
表 1 实验区域表层底泥的物理化学性质 Table 1 Physio-chemical characteristics of surface layer of sampled sediment core |
本研究采集了太湖流域底泥样品和水样,经过多次富集、 平板划线分离、 生理生化特性实验等方法分离与筛选出土著脱氮微生物.
采用甲基丙烯酸-β-羟乙酯(HEMA)、 丙烯酸羟乙酯(HEA)与蒸馏水按特定体积比均匀混合,并用氮气进行密封,在60 Co-γ射线(辐射剂量1×104 Gy),-78℃温度条件下辐照制备形成具有生物相容性的固定化脱氮微生物的载体[10]. 将固定化载体放在去离子水中浸润3 h,直至完全溶胀(此时载体的孔径大小约为20-100 μm),然后将已经分离好的菌液与载体混合,同时加入培养微生物的营养液,在8h好氧曝气培养,4 h厌氧培养(每72 h循环6次). 固定化载体具有微环境可供高效脱氮微生物生长并进行繁殖,制备成固定化脱氮微生物.
1.3 室内-原位实验设计底泥和上覆水均从亲水河现场进行采集. 根据沿河流域的污染程度和工业源排放情况,研究选取了河道未受干扰的段位进行样品采集,以保证模拟原位实验的准确性. 底泥通过有机玻璃管(长度为60 cm,内径9 cm)进行采集,取样的底泥厚度约为30 cm. 共采集24根完整底泥柱在室内进行模拟原位实验. 同时采集原位上覆水. 所有的底泥柱以及水样在4 h内送至实验室进行处理. 室内实验培养时间2014-06-2014-08.
24根底泥柱被分成4个不同的微环境系统,分别为:处理组A,不做任何处理的裸泥组,作为对照; 处理组B,添加了沉水植物伊乐藻(Elodea nuttallii)的处理组,选取长势良好的伊乐藻进行种植,伊乐藻长度为10 cm,株数为5株,株间距约为3 cm,每株的生物量约为10 g; 处理组C,添加了固定化脱氮微生物(INCB)的处理组,新鲜固定化载体湿重约为每包(60±1)g,每包总生物量约为4.5×105cells ·L-1; 处理组D,添加了固定化氮循环菌和沉水植物伊乐藻,添加量如上所述. 所有的处理组均放在室温约为20℃±1℃进行培养. 在为期约1个月的室内培养结束后,进行相关水质及理化数据的测定. 分别考察固定化氮循环菌、 伊乐藻、 伊乐藻-固定化脱氮微生物联用技术对河道水质改善的效果.
1.4 反硝化速率和厌氧氨氧化速率测定反硝化速率和厌氧氨氧化速率的测定是通过Risgaard-Peterenx[11]和Trimmer等[12]提出15 N同位素标记法进行的. 底泥柱在实验之前,预处理放置24 h,以去除水体中残余的NO-. 随后,将适宜浓度的同位素混合稀释液加入到底泥柱中. 分为3个处理组进行投加,即:①15 NH4+ (15 N at.%=99.11),②15 NH4++14NO3-,③ 15 NO3-(15 N at.%=99.16),使得最终每个处理组中的15 N浓度约为100 μmol ·L-1. 每个底泥柱有3个平行处理. 在添加15 N同位素之后,每个柱子上面用密封性能良好的橡胶塞进行密封,留有10 cm高度的顶空进行气体采集. 整个底泥柱在室温(25℃±1℃)下培养24 h后,向每个柱体中投加100 μL的ZnCl2溶液(质量分数50%),使柱体中的微生物停止活动,然后采集水样,通过膜接口质谱仪(MIMS,Bay Instruments,Easton,MD,USA)进行同位素28 N2、 29 N2、 30 N2的测定. 最后通过如下公式所述计算得到厌氧氨氧化速率及反硝化速率.
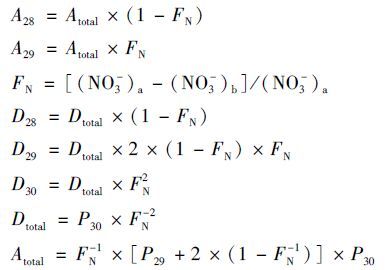
研究通过高通量测序平台(MiSeq)对4个不同的生态处理组进行微生物群落多样性的对比分析.
DNA提取:每个样品分别称取约500 mg后利用FastDNA Soil Kit (MP Biomedicals,CA,USA) 提取各样品的总DNA,使用NanoDropND-1000 (NanoDrop Technologies,Willmington,DE,USA) 对提取的总DNA样品进行浓度和纯度检测,保证各样品浓度大于20 ng ·μL-1,A260/A280介于1.80和2.00之间.
PCR扩增及纯化:利用上下游分别带有不同的9碱基标签序列的V1/V2区引物对各样品进行PCR扩增,上游引物序列结构为“G+8碱基标签序列+AGAGTTTGATYMTGGCTCAG”,下游引物序列结构为“G+8碱基标签序列+TGCTGCCTCCCG TAGGAGT”. 用TaKaRa Ex Taq(TaKaRa Bio,Japan)进行PCR扩增,单个样品做4个PCR平行. 扩增条件为:98℃ 5 min; 20循环(98℃ 30 s,50℃ 30 s,72℃ 40 s); 72℃ 10 min. PCR完成后将每个样品的4个平行PCR体系混合后利用试剂盒进行PCR产物纯化,最终洗脱体积为30 μL.
建库以及MiSeq测序:准确移取等质量的100 ng的PCR纯化产物混合均匀,进行建库. 建库完成后利用2100 Bioanalyzer (Agilent Technologies,Santa Clara,USA)进行芯片电泳以确认建库成功. 同时再次使用Qubit 2.0 Fluorometer对文库进行精确定量. 将建库成功的文库按比例混合、 变性并稀释至终浓度8 pmol ·L-1后进行Miseq上机测序.
数据分析:通过Sickle软件(https://github.com/najoshi/sickle)去除低质量序列,用Mothur软件包(http://www.mothur.org/wiki/Main_Page)对测序数据按样品筛分,然后在Mothur官网提供的标准流程进行降噪处理,最终对降噪数据进行比对分析,获得样品中微生物群落结构信息.
1.6 水质指标及测定方法氨氮采用纳氏试剂分光光度法测定; 硝态氮采用紫外分光光度法进行测定; 总氮采用过硫酸钾氧化紫外分光光度法(日本岛津UV-2450)测定; pH采用便携式pH计(YSI pH100)进行测定; DO采用便携式溶氧仪(YSI 550A)进行测定; 底泥表层DO采用溶解氧微电极系统进行测定(Micro TX3,Presens,德国).
1.7 数据分析方法采用了SPSS 13.0 数据处理包进行数据分析; 采用单向或双向方差分析(ANOVA)来区别不同处理组样品之间的差异性; 采用皮尔逊相关分析(Pearson correlation)进一步评价数据变化的相关性; 采用Origin 8.0进行图形绘制. 不同处理组之间的显著性差异水平设置为P<0.05.
2 结果与讨论2.1 室内模拟原位实验水质改善效果对比
2.1.1 氮素营养盐水平变化模拟实验研究于2014-06-2014-08月进行. 采用生态模拟修复柱进行,在柱内微生物和植物已经稳定生长后,用原位河道水进行更换,然后跟踪监测一个星期的水质变化,每天下午17:00进行水样的采集. 总氮、 硝氮、 氨氮、 亚硝氮的含量变化如图 2-5所示. 实验结果表明,4个处理组中,添加了伊乐藻和固定化氮循环菌的处理组总氮去除率为72.03%(图 2),硝氮的去除率为76.65%(图 3),在4个处理中均为最高,水体总氮与硝氮变化规律一致(r=0.993,P<0.05)说明沉水植物与固定化氮循环菌联用技术强化水体的氮素脱除能力. 从图 4和图 5中,水体中氨氮和亚硝氮含量本底值不高,且呈现一个波动变化. 添加了固定化高效脱氮微生物的处理组,氨氮和亚硝氮含量均表现为初期增加,而后逐渐降低的趋势; 未添加固定化脱氮微生物的处理组没有增加的现象. 由于脱氮细菌的存在,水体中存在大量的氨化反应、 硝化反应、 硝酸盐异化还原为铵以及反硝化反应等生物化学作用,氨氮在好氧状态下被氨氧化细菌转化为亚硝氮,同时在厌氧条件下,,亚硝氮可通过硝酸盐异化还原为铵(DNRA)转化为氨氮,这些过程都会导致水体中的氨氮和亚硝氮在短时间发生互相转化[13]. 因此氨氮和亚硝氮短期升高,可能是由于水体中同时存在氨化细菌的矿化作用和氨氧化细菌所造成. 随后随着硝化细菌的不断转化,使得氨氮和亚硝氮含量逐渐下降. 从图 5中可以看出,添加了伊乐藻和固定化脱氮微生物的处理组的亚硝氮的含量高于裸泥组的亚硝氮浓度(ANOVA,P<0.05). 亚硝氮作为水体中脱氮过程一个中间产物[14, 15],在柱体脱氮过程中会出现一个短暂性的累积现象.
 | 图 2 实验柱中不同处理组总氮变化对比 Fig. 2 Comparison of variation of TN in different groups |
 | 图 3 实验柱中不同处理组硝氮浓度变化 Fig. 3 Comparison of variation of NO3--N in different groups |
 | 图 4 实验柱中不同处理组氨氮变化对比 Fig. 4 Comparison of variation of NH4+-N in different groups |
 | 图 5 实验柱中不同处理组亚硝氮变化对比 Fig. 5 Comparison of variation of NO2--N in different groups |
模拟原位河道的反应柱中种植了伊乐藻的处理组与未种植伊乐藻的处理组,溶解氧的变化有明显的差异. 从图 6可以看出,种植了伊乐藻的处理组,最高溶解氧浓度为23.7mg ·L-1,时间为14:00左右,由于此时一般为一天光照强度最强的时候,因此沉水植物伊乐藻的光合作用也最强,使得水体中的溶解氧浓度达到过饱和状态. 同时,在16:00以后,由于植物光合作用减弱,水体中的溶解氧浓度也逐渐下降. 通过Pearson相关性分析发现,添加了伊乐藻的处理组中总氮与溶解氧呈负相关(P<0.05, r=-0.894)实验结果表明,伊乐藻的稳定生长增加柱体底泥的溶解氧,改变了泥水界面处的溶解氧情况,这与之前的一些研究具有一致性[16]. 白天期间,沉水植物伊乐藻通过光合作用会产生氧气[17]. 同时,伊乐藻的根系会释放氧气,从而改变根系周围的氧气分布. 氧气侵蚀深度的变化[18],使得原本厌氧或缺氧的底泥变成了缺氧-好氧间歇分布的区域,这样的变化为泥水界面处的脱氮微生物提供了适宜的生存环境,从而进一步提高了脱氮效率.
 | 图 6 实验柱中不同处理组溶解氧变化 Fig. 6 Variation of oxygen profiles in different treatment groups |
4个处理组中,厌氧氨氧化的氮气产量,反硝化过程的氮气产生速率以及柱体总的氮气产生速率均得到了增强(图 7),仅添加了固定化脱氮微生物的处理组的N2产生速率在4个处理组中最高,说明脱氮微生物活动得到了明显的增强,对于河道脱氮过程具有明显的影响,这些过程与河道生态系统中的氮素循环以及生态修复都有着密切的关系[19]. 对比添加了固定化脱氮微生物的两个处理组的N2产生速率,虽然两者的氮气产生速率均高于未添加固定化脱氮微生物的处理组,但是添加了伊乐藻的处理组(处理组B和处理组D)的N2产生速率要显著低于没有添加伊乐藻的处理组(处理组A高于处理组C,处理组C要高于处理组D)(ANOVA,P<0.05). 由于沉水植物的存在,可能会在一定程度上抑制厌氧氨氧化过程以及反硝化过程的氮气产生速率,从而导致了底泥柱中总的N2产量的下降. 脱氮微生物将水体中的氨氮和硝氮转化为N2的过程,是一个较为复杂的生物化学过程,其中会经过N2O的一个中间产气过程,而将N2O转化为N2的过程,需要一个在相对缺氧的条件下进行[20, 21]. 沉水植物伊乐藻的存在,会丰富泥水界面处的氧气含量,改变氧气含量(图 6),因此会在一定程度上抑制N2O转化为N2的过程. 同时,沉水植物通过根部释放氧气,在改变沉积物的氧气侵蚀深度后,还会引起根区土壤的氧化还原电位的改变[22]. 固定化脱氮微生物从载体释放到水体及泥水界面中,因为其中氨化细菌、 氨氧化细菌、 硝化细菌以及反硝化细菌的存在,会引发一系列的生物化学过程,从而改变水体中的NH4+-N和NO2--N的含量. 反硝化过程与厌氧氨氧化过程互相之间有着紧密的联系,有研究发现厌氧氨氧化过程需要的NO2-可以来自于反硝化细菌还原NO3-的过程[23]. 同时,有报道已经发现厌氧氨氧化过程和反硝化过程在低C/N比的情况下,具有良好的协同作用[24, 25],同时,在微溶解氧区域(OMZ)内,反硝化过程和厌氧氨氧化过程可以同时发生[26]. 在表层的泥水界面中,氨氮的氧化导致孔隙水中有更多能被反硝化菌利用的硝氮的生成[27],同时硝氮的还原也使得孔隙水中更多的亚硝酸盐可以被厌氧氨氧化细菌利用. 因为研究区域的NO3--N含量要远高于NO2--N的含量,因此更多的硝酸盐氧化过程对于厌氧氨氧化过程更加有利. 因此,添加了INCB的实验柱体沉积物中厌氧氨化过程和反硝化过程大大提高,表现为N2产生速率明显提高.
 | 图 7 实验柱中不同处理组氮气产生速率变化对比 Fig. 7 Comparison of N2 production rate in different treatment groups |
4个不同处理组的底泥柱中,添加了固定化脱氮微生物的处理组与固定化脱氮微生物与伊乐藻联用的处理中微生物多样性不断增加,其中以变形菌门(Proteobacteria)最多,在裸泥组、 沉水植物组、 固定化脱氮微生物组以及固定化脱氮微生物与伊乐藻联用组所占比例分别为96.98%、 95.03%、 58.95%和63.24%(图 8). 变形菌门中常见的脱氮微生物有氨化细菌、硝化细菌以及反硝化细菌[28]. 同时,添加了固定化脱氮微生物的处理组中浮霉菌门(Planctomycetes)的丰度得到明显提高,而多数厌氧氨氧化细菌多见于浮霉菌门[29]. 从微生物的多样性的分析上看,添加了固定化脱氮微生物后,柱体中的反硝化细菌和厌氧氨氧化细菌具有更高的丰度. 酸杆菌门(Acidobbacteria)和拟杆菌门(Bacteroidetes)同样具有较高的丰度,这些细菌在生态系统中也具有重要的作用. 4个处理组中微生物群落结构的改变主要是因为受固定化微生物和沉水植物伊乐藻的影响. 固定化脱氮微生物带来了大量的氨化细菌、 硝化细菌和反硝化细菌. 同时,由于沉水植物伊乐藻的存在,植物根系会释放碳、 O2,会改变根系周围土壤底泥的理化条件[30],从而使得底泥柱中的微生物的群落结构发生改变. 同时,温度、 营养盐、 光照等条件也会对脱氮等微生物的群落结构造成影响.
 | 图 8 实验柱中不同处理组的微生物群落变化情况 Fig. 8 Variation of microbial communities in different treatment groups |
(1)伊乐藻与固定化脱氮微生物联合作用改变了泥水界面的溶解氧浓度,为脱氮微生物提供了缺氧-好氧的生存环境,促进微生物氮素转化过程.
(2)伊乐藻与固定化脱氮微生物联用技术提高了沉积物的反硝化速率,同时厌氧氨氧化速率也得到提高. E-INCB技术可以利用反硝化细菌与厌氧氨氧化细菌的协同促进关系,同时提高反硝化速率与厌氧氨氧化速率.
(3)添加了伊乐藻和固定化脱氮微生物的处理组,变形菌门(Proteobacteria)以及浮霉菌门(Planctomycetes)类菌群均得到了显著的提高,说明加入固定化脱氮微生物以后,显著增强了河道底泥中的脱氮微生物的多样性.
(4)室内模拟实验表明,伊乐藻与固定化脱氮微生物联用技术可以有效去除河道水体中的氮素,提高亲水河的水体自净能力.
| [1] | Binnerup S J, Sørensen J. Nitrate and nitrite microgradients in barley rhizosphere as detected by a highly sensitive denitrification bioassay[J]. Applied and Environmental Microbiology, 1992, 58 (8): 2375-2380. |
| [2] | 蒋跃, 童琰, 由文辉, 等. 3种浮床植物生长特性及氮、磷吸收的优化配置研究[J]. 中国环境科学, 2011, 31 (5): 774-780. |
| [3] | 李正魁, 濮培民. 秋冬季环境下固定化氮循环细菌净化湖泊水体氮污染动态模拟[J]. 湖泊科学, 2000, 12 (4): 321-326. |
| [4] | 陈祈春, 李正魁, 王易超, 等. 沉水植物床-固定化微生物技术在水源地修复中的应用研究[J]. 环境科学, 2012, 33 (1): 83-87. |
| [5] | 李海英, 冯慕华, 李玲, 等. 微曝气生态浮床净化入湖河口污染河水原位模型实验[J]. 湖泊科学, 2009, 21 (6): 782-788. |
| [6] | 刘丹丹, 李正魁, 叶忠香, 等. 伊乐藻和氮循环菌技术对太湖氮素吸收和反硝化的影响[J]. 环境科学, 2014, 35 (10): 3764-3768. |
| [7] | Wang Y C, Li Z K, Zhou L, et al. Effects of macrophyte-associated nitrogen cycling bacteria on denitrification in the sediments of the eutrophic Gonghu Bay, Taihu Lake[J]. Hydrobiologia, 2013, 700 (1): 329-341. |
| [8] | Tao W D, Wang J. Effects of vegetation, limestone and aeration on nitritation, anammox and denitrification in wetland treatment systems[J]. Ecological Engineering, 2009, 35 (5): 836-842. |
| [9] | Zhao Y Q, Xia Y Q, Kana T M, et al. Seasonal variation and controlling factors of anaerobic ammonium oxidation in freshwater river sediments in the Taihu Lake region of China[J]. Chemosphere, 2013, 93 (9): 2124-2131. |
| [10] | Chen F Z, Song X L, Hu Y H, et al. Water quality improvement and phytoplankton response in the drinking water source in Meiliang Bay of Lake Taihu, China[J]. Ecological Engineering, 2009, 35 (11): 1637-1645. |
| [11] | Risgaard-Petersen N, Nielsen L P, Rysgaard S, et al. Erratum: Application of the isotope pairing technique in sediments where anammox and denitrification co-exist[J]. Limnology and Oceanography: Methods, 2004, 2 : 315. |
| [12] | Trimmer M, Risgaard-Petersen N, Nicholls J C, et al. Direct measurement of anaerobic ammonium oxidation (anammox) and denitrification in intact sediment cores[J]. Marine Ecology Progress Series, 2006, 326 : 37-47. |
| [13] | Kumar M, Lin J G. Co-existence of anammox and denitrification for simultaneous nitrogen and carbon removal-Strategies and issues[J]. Journal of Hazardous Materials, 2010, 178 (1-3): 1-9. |
| [14] | Seitzinger S P. Denitrification in freshwater and coastal marine ecosystems: ecological and geochemical significance[J]. Limnology and Oceanography, 1988, 33 (4): 702-724. |
| [15] | 彭剑峰, 宋永会, 袁鹏, 等. 循环流廊道湿地中氮归趋过程模拟研究[J]. 环境科学, 2014, 35 (4): 1311-1317. |
| [16] | Risgaard-Petersen N, Jensen K. Nitrification and denitrification in the rhizosphere of the aquatic macrophyte Lobelia dortmanna L.[J]. Limnology and Oceanography, 1997, 42 (3): 529-537. |
| [17] | Laskov C, Horn O, Hupfer M. Environmental factors regulating the radial oxygen loss from roots of Myriophyllum spicatum and Potamogeton crispus[J]. Aquatic Botany, 2006, 84 (4): 333-340. |
| [18] | Jespersen D N, Sorrell B K, Brix H. Growth and root oxygen release by Typha latifolia and its effects on sediment methanogenesis[J]. Aquatic Botany, 1998, 61 (3): 165-180. |
| [19] | Mortensen E, Jeppesen E, Søndergaard M, et al. Nutrient dynamics and biological structure in shallow freshwater and brackish lakes[M]. Netherlands: Springer, 1994. |
| [20] | Lam P, Kuypers M M M. Microbial nitrogen cycling processes in oxygen minimum zones[J]. Annual Review of Marine Science, 2011, 3 (1): 317-345. |
| [21] | 陈诺, 廖婷婷, 王睿, 等. 碳底物含量对厌氧条件下水稻土N2, N2O, NO, CO2和CH4排放的影响[J]. 环境科学, 2014, 35 (9): 3595-3604. |
| [22] | Christensen P B, Revsbech N P, Sand-Jensen K. Microsensor analysis of oxygen in the rhizosphere of the aquatic macrophyte Littorella uniflora (L.) ascherson[J]. Plant Physiology, 1994, 105 (3): 847-852. |
| [23] | Risgaard-Petersen N, Meyer R L, Revsbech N P. Denitrification and anaerobic ammonium oxidation in sediments: effects of microphytobenthos and NO3-[J]. Aquatic Microbial Ecology, 2005, 40 (1): 67-76. |
| [24] | Long A, Heitman J, Tobias C, et al. Co-occurring anammox, denitrification, and codenitrification in agricultural soils[J]. Applied and Environmental Microbiology, 2013, 79 (1): 168-176. |
| [25] | Lotti T, Van Der Star W R L, Kleerebezem R, et al. The effect of nitrite inhibition on the anammox process[J]. Water Research, 2012, 46 (8): 2559-2569. |
| [26] | Ward B B, Devol A H, Rich J J, et al. Denitrification as the dominant nitrogen loss process in the Arabian Sea[J]. Nature, 2009, 461 (7260): 78-81. |
| [27] | An S, Joye S B. Enhancement of coupled nitrification-denitrification by benthic photosynthesis in shallow estuarine sediments[J]. Limnology and Oceanography, 2001, 46 (1): 62-74. |
| [28] | 方芳, 陈少华. 功能基因在反硝化菌群生态学研究中的应用[J]. 生态学杂志, 2010, 29 (9): 1836-1845. |