 2016, Vol. 37
2016, Vol. 37



溴酸盐是一种典型的饮用水臭氧消毒副产物. 当原水中含有Br-时,Br-可以被臭氧分子直接氧化生成溴酸盐或者通过 ·OH氧化生成溴酸盐[1]. 目前,很多研究表明溴酸盐可以诱发试验动物肾脏细胞肿瘤[2, 3],另外许多遗传毒性试验也呈现阳性结果[4, 5, 6, 7]. 因此国际癌症研究机构将溴酸盐归为2B类致癌物. 我国的生活饮用水卫生标准(GB 5749-2006),首次将溴酸盐纳入饮用水国家标准,规定在使用臭氧时,水质溴酸盐含量的限值为0.01 mg ·L-1[8]. 同样,我国食品安全国家标准(GB 19298-2014)中也规定,包装饮用水中溴酸盐含量不得超过0.01 mg ·L-1[9].
然而溴酸盐不仅仅在饮用水中产生,还可以通过氯化或臭氧化电厂冷却水,次氯酸钠等消毒剂的生产和使用等过程产生[10]. 同时溴酸盐在食品加工(我国已经禁用)、 黄金提取和羊毛生产等领域中有广泛的应用[11]. 这些人类活动势必会导致溴酸盐进入自然水体,同时由于溴酸盐一旦进入水体则具有高度的稳定性,因此这会促进溴酸盐对自然水体的污染[11].
以前普遍认为溴酸盐在地表水和地下水中一般不存在,但是随着溴酸盐检测技术的发展,溴酸盐已经在许多调查研究中检出[12]. 例如,英国地下水蓄水层中溴酸盐的含量达到2 mg ·L-1[11]; 印度西南沿海喀拉拉邦自来水中浓度为5.34 μg ·L-1,但在井水中则未检出[13]; 美国洛杉矶两处水库的溴酸盐含量分别为68 μg ·L-1和106 μg ·L-1[14]; 菲律宾马尼拉自来水中溴酸盐含量为7-138 μg ·L-1,两条河流的平均溴酸盐浓度为15-80 μg ·L-1. 而地下水和废水样品中溴酸盐含量高达246 μg ·L-1和342 μg ·L-1[15]. 在我国,Wu等[16]调研发现上海水样的溴酸盐浓度最高可达28.4 μg ·L-1. 刘勇建等[17]研究发现北京有3个饮用水厂的源水中含有溴酸盐,含量为0.6-2.2 μg ·L-1. 杨永亮等[18]发现沈阳细河浅层地下水中,有两处溴酸盐的浓度存在超标的情况,分别为31 μg ·L-1和15 μg ·L-1.
由以上研究可知,溴酸盐已经对许多国家和地区的自然水体造成了污染. 但是有关溴酸盐对水生生态系统的潜在风险还不确定,对水生生物的影响还没有系统的报道,因此评估溴酸盐对水生生态系统的潜在影响,探讨其对水体不同营养级生物的毒性效应具有重要意义.
一个健全的水生生态系统主要包括单细胞生物、 浮游植物、 浮游动物和鱼类等营养级生物. 其中,发光菌、 绿藻、 水蚤和斑马鱼是几个不同营养级的代表性生物,并且在水生生物毒理学中也常常作为判断某种物质毒性的标准测试生物. 本研究通过分析溴酸盐对明亮发光杆菌、 费氏弧菌、 青海弧菌、 斜生栅藻、 普通小球藻、 大型蚤、 裸腹蚤和斑马鱼的急性毒性,探讨溴酸盐毒性作用的特征,以期为评价溴酸盐的毒性效应提供基础数据,判断其对水生生物可能造成的危害,为有关生态风险评价提供参考依据.
1 材料与方法 1.1 试验材料溴酸钾、 溴酸钠、 溴化钾购于国药集团化学试剂有限公司,分析纯.
发光菌(明亮发光杆菌502、 费氏弧菌、 青海弧菌Q67)冻干粉,由北京滨松光子技术股份有限公司提供.
绿藻(斜生栅藻 Scenedesmus obliquus、 普通小球藻 Chlorella vulgaris),购于中国科学院水生生物研究所淡水藻种库,编号分别为FACHB417,FACHB1068. 培养条件为:温度25℃±1℃,光照强度约3 000 lx,光暗比为12 ∶12,在无菌条件下用BG11培养基进行培养,每天定时人工振荡若干次.
水蚤(大型蚤 Daphnia magna、 裸腹蚤 Moina),大型蚤购买于中国科学院生态环境研究中心,裸腹蚤捕捞于松花江. 两者均在实验室条件下培养3代以上,形成单克隆纯品系. 培养条件为:温度20℃±1℃,光照强度800-1 000 lx,光暗比为16 ∶8. 培养用水为曝气3 d以上的自来水,每天用去除培养基的普通小球藻喂食.
斑马鱼,购买于中科院水生生物研究所,平均体长3 cm±0.5 cm,体重0.3 g±0.1 g. 实验室驯化1个月以上. 培养条件为:雌雄比1 ∶1,水温25℃±1℃、 光暗比14 ∶10,光照强度约1 000 lx. 培养用水为曝气3 d以上的自来水,每日喂食两次冷冻赤虫.
1.2 试验方法 1.2.1 发光菌发光抑制试验首先按照各发光菌的使用说明书对菌种进行复苏. 具体操作步骤为:将明亮发光杆菌502、 费氏弧菌、 青海弧菌Q67冻干粉置于室温下平衡15 min,分别加入一定量的复苏稀释液,溶解后的冻干粉置于室温下10 min后即可用于样品的测定.
取调节好渗透压的样品2 mL于测试管中,以复苏稀释液作为空白对照,加入0.05 mL发光菌复苏液,轻轻振荡,使之充分混匀,室温放置15 min后,用BHP9514型饮用水安全快速检测仪测定发光菌的发光强度.
按照以下公式计算样品相对发光强度:

参照文献[19]进行试验. 根据预试验结果,在无菌条件下,取一定量处于对数生长期的藻液及污染物储备液,用新鲜无菌BG11培养基稀释,使斜生栅藻和普通小球藻的初始藻密度约为12.5×105 ind ·mL-1和12.5×105 cells ·mL-1,污染物浓度按等对数间距设置为0、 0.5、 1、 2、 4、 8 mmol ·L-1. 记录第0、 24、 48、 72、 96 h藻液在波长为680 nm的光密度,通过藻细胞血球计数板显微计数与光密度之间的线性方程计算藻细胞密度,进而计算生长抑制率,通过抑制率与污染物浓度对数之间的线性方程计算96 h半抑制浓度EC50.
藻的生长速率及生长抑制百分率按下式计算:
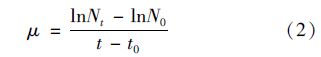
参照文献[20]进行试验. 根据预试验结果,按照等对数间距设置浓度梯度,每个浓度设置5组,每组放5只个体均匀,出生6-24 h的水蚤. 试验容器为50 mL烧杯,试验过程不换试验溶液,中间不喂食. 记录大型蚤和裸腹蚤24 h和48 h累计运动抑制数及死亡数. 通过寇氏法计算24 h和48 h半抑制浓度EC50和半致死浓度LC50及相应的95%置信区间.
计算公式以LC50为例,如下:
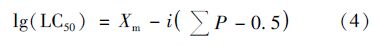
95%置信区间的计算公式如下.
先求标准差:
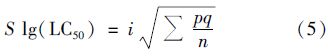
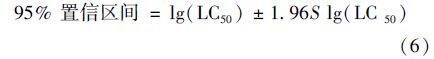
参照文献[21]进行试验. 根据预试验,按照等对数间距设置浓度梯度,每个浓度设置3组,每组放10尾,个体差异不大,游动灵活的斑马鱼于2 000 mL烧杯中进行试验,试验过程不换试验溶液,中间不喂食. 试验开始后,记录24、 48、 72、 96 h斑马鱼死亡数目及任何不正常的行为. 通过寇氏法计算24、 48、 72、 96 h半致死浓度LC50及相应的95%置信区间.
1.3 数据处理试验数据中试验组和对照组之间的显著性差异采用SPSS 20.0统计分析软件进行分析,以P<0.05表示差异具有统计学意义.
2 结果与讨论 2.1 溴酸盐对发光菌的急性毒性效应发光菌毒性测试方法具有快速、 灵敏等特点,广泛用于饮用水及废水的急性毒性测定[22],通过测定发光菌发光强度的变化来反映水体的污染情况. 图 1是不同浓度的溴酸钾、溴酸钠和溴化钾分别作用于明亮发光杆菌502、 费氏弧菌、 青海弧菌Q67 15 min后,对各个发光菌的相对发光强度的影响. 从中可以看出,在如此大的测定浓度范围内,无论是溴酸钾、 溴酸钠还是溴化钾对3种发光菌的相对发光强度均在97%-120%之间,除个别数据外,整体上试验组与空白组相比没有显著性差异,并且随着污染物浓度的升高,发光菌的相对发光强度不呈规律性变化. 因此说明发光菌对溴酸盐的毒性效应反应不明显,不适于检测和评价溴酸盐的毒性效应.
 | *表示与空白组相比具有显著性差异P<0.05,**表示P<0.01图 1 污染物对发光菌相对发光强度的影响 Fig. 1 Effects of KBrO3,NaBrO3 and KBr on the relative luminous intensity of luminescent bacteria |
绿藻是水生食物链的初级生产者[23],外源化合物对绿藻产生的毒性效应将影响水生食物链的能量传递,进而影响高营养级生物和整个水生生态系统[24]. 藻类因其个体小、 繁殖快、 对毒物敏感[25]、 在较短时间内就可得到化学物质对其世代及种群水平的影响[26],并可直接观察细胞水平上的中毒症状等特点而得到广泛的应用.
图 2为藻细胞密度与光密度之间的线性关系. 从中可知,在试验所考察的不同藻密度下,显微计数得到的藻细胞密度和680 nm的光密度之间呈现良好的线性关系,斜生栅藻的线性回归方程为y=112.82x-0.022 6 (R2=0.998 6),普通小球藻的线性回归方程为y=193.88x-0.621 8 (R2=0.998 3),根据相应的线性回归方程,可以通过测定藻液在680 nm处的吸光度代替显微计数来确定藻细胞密度,简化试验操作.
 | 图 2 斜生栅藻和普通小球藻与光密度之间的线性关系 Fig. 2 Linear relationship between cell density and optical density of Scenedesmus obliquus and Chlorella vulgaris |
图 3为斜生栅藻暴露在不同浓度的溴酸钾、 溴酸钠和溴化钾中的生长曲线. 从中可以看出,当暴露在溴酸钾和溴酸钠条件下,斜生栅藻都受到了不同程度的影响,随着污染物浓度的升高,斜生栅藻的生长速度逐渐变慢,当污染物浓度达到8 mmol ·L-1时,斜生栅藻几乎停止了生长. 但是当暴露在溴化钾时,斜生栅藻的生长几乎没有受到影响.
 | 图 3 污染物对斜生栅藻生长的影响 Fig. 3 Effects of KBrO3,NaBrO3 and KBr on the growth of Scenedesmus obliquus |
图 4是在不同浓度的污染物下暴露96 h后,斜生栅藻的藻密度. 从中可以看出,在溴酸钾和溴酸钠的影响下,随着污染物浓度的升高,藻密度逐渐下降. 当溴酸钾和溴酸钠浓度为8 mmol ·L-1时,相应的藻密度分别为空白值的50.5%和55.2%,与空白值相比具有显著性差异. 而在溴化钾的暴露下,试验组与空白组相比没有显著性差异. 表 1是在暴露96 h后,各污染物对斜生栅藻的半抑制浓度. 从中可以看出溴酸钾对斜生栅藻的96 h EC50为4.42 mmol ·L-1,换算成质量浓度为738.18 mg ·L-1; 溴酸钠对斜生栅藻的96 h EC50为3.58 mmol ·L-1,换算成质量浓度为540.26 mg ·L-1; 由于溴化钾没有对斜生栅藻的生长产生抑制效果,所以没有半抑制浓度数值. 对比可知,虽然溴酸钾和溴酸钠的96 h EC50有一定差别,但与溴化钾相比差别更大. 通过以上结果及分析可以得出,溴酸钾与溴酸钠对斜生栅藻的毒性效应很相似,其差别可能是由阳离子的不同而引起的. 但两者与溴化钾对斜生栅藻的影响差别非常显著,这是由阴离子的不同而引起的,因此可以说明造成斜生栅藻生长受到抑制的主要原因是溴酸盐(即BrO3-)的毒性作用的影响.
 | *表示与空白组相比具有显著性差异P<0.05,**表示P<0.01图 4 暴露96 h后污染物对斜生栅藻生长的影响 Fig. 4 Effects of KBrO3,NaBrO3 and KBr on the growth of Scenedesmus obliquus after 96 h exposure |
| 表 1 污染物对斜生栅藻的急性毒性 Table 1 Acute toxicity of KBrO3,NaBrO3 and KBr on Scenedesmus obliquus |
图 5是试验开始96 h后,在溴酸盐暴露下斜生栅藻藻液的宏观照片. 从中可以看出,在溴酸盐的作用下斜生栅藻藻液颜色逐渐黄化失绿,说明藻细胞密度逐渐降低,死亡藻细胞逐渐增多. 显微观察发现经溴酸盐暴露后,藻细胞出现粘连和聚集现象,沉淀现象明显,同时形成了一些8细胞群体,并逐渐处于优势地位,而对照组中依然是4细胞群体处于优势,这可能是由于为了抵抗外来毒物的侵袭而自发形成的一种抵御状态. 另外死细胞及藻细胞分解碎片随着溴酸盐浓度的增加和培养时间的延长逐渐增多. 图 6分别是暴露在0 mmol ·L-1和8 mmol ·L-1溴酸盐96 h后斜生栅藻的细胞形态. 从中可以发现在溴酸盐的作用下,藻细胞变形,干瘪,细胞分裂畸形,出现异型胞,藻细胞处于濒临死亡的状态.
 | 图 5 斜生栅藻藻液的照片 Fig. 5 Photo of the culture solution of Scenedesmus obliquus |
 | (a) 0 mmol ·L-1; (b) 8 mmol ·L-1 图 6 暴露96 h后斜生栅藻的细胞形态 Fig. 6 Cell morphology of Scenedesmus obliquus after 96 h exposure |
为了考察溴酸盐对其他藻细胞是否同样存在抑制作用,以普通小球藻为受试生物考察溴酸盐的毒性效应. 图 7分别为普通小球藻暴露在不同浓度的溴酸钾、 溴酸钠、 溴化钾96 h内的生长曲线. 从中可以看出,普通小球藻的生长并没有出现像斜生栅藻那样随着溴酸盐浓度的升高生长速度明显减慢的现象,即使在溴酸盐为8 mmol ·L-1时,也没有出现非常明显的抑制效果. 图 8为暴露96 h后普通小球藻的藻密度. 从中可以发现,只有当溴酸钾和溴酸钠的浓度大于4 mmol ·L-1时,试验组才与空白组出现显著性差异,而溴化钾试验组与空白组则始终没有显著性差异. 当溴酸钾和溴酸钠浓度为8 mmol ·L-1时,相应的藻密度为空白值的77.3%和79.4%. 由于96 h暴露后,普通小球藻的半抑制率在最高浓度下也没有超过50%,因此没有给出对普通小球藻的半抑制浓度. 因此通过上述结果可以发现溴酸盐对普通小球藻的生长抑制作用要比对斜生栅藻的生长抑制作用小得多.
 | 图 7 污染物对普通小球藻生长的影响 Fig. 7 Effects of KBrO3,NaBrO3 and KBr on the growth of Chlorella vulgaris |
 | *表示与空白组相比具有显著性差异P<0.05,**表示P<0.01图 8 暴露96 h后污染物对普通小球藻生长的影响 Fig. 8 Effects of KBrO3,NaBrO3 and KBr on the growth of Chlorella vulgaris after 96 h exposure |
相比而言,斜生栅藻对溴酸盐的毒性更为敏感,同时说明并非所有浮游藻类都对溴酸盐的毒性敏感. 这主要是由于不同藻类自身生理差异造成的,这种差异还影响了不同藻类对污染物的敏感性和耐受性[27]. Fisher等[28]以球等鞭金藻为试验生物考察了溴酸钠的毒性效应,结果表明球等鞭金藻对溴酸钠的耐受性非常强,72 h EC50为15 954 mg ·L-1,最大无影响浓度为5 277 mg ·L-1,最低有影响浓度为10 554 mg ·L-1,这与本研究结果差异较大,由此也可进一步说明不同藻类对溴酸盐毒性的敏感性和耐受性存在差异.
对于普通小球藻,仍然选取在溴酸盐溶液中暴露96 h后的藻液进行观察. 图 9和图 10分别为普通小球藻藻液的宏观照片和光学显微镜观察照片. 从图 9可以看出各浓度组藻液的颜色差别不大,并且图 10中对照组和溴酸盐暴露组的藻细胞大都比较饱满、 完整,细胞表面光滑,因此可以说明溴酸盐对普通小球藻的生长和细胞形态影响不大,与斜生栅藻相比普通小球藻对溴酸盐的毒性效应不敏感.
 | 图 9 普通小球藻藻液的照片 Fig. 9 Photo of the culture solution of Chlorella vulgaris |
 | (a) 0 mmol ·L-1; (b) 8 mmol ·L-1 图 10 暴露96 h后普通小球藻的细胞形态 Fig. 10 Cell morphology of Chlorella vulgaris after 96 h exposure |
水蚤既是藻类、 细菌、 有机残渣的捕食者,又是鱼类等一些水生生物的饵料,是水生食物链的重要环节[29],也是对污染物较为敏感的关键性生物[30],对水生生态系统的健康起着重要作用. 它广泛地被用作标准化的测试方法和基本的生态毒性研究.
当外界有毒物质进入水环境后,会对大型蚤等浮游动物产生多方面的影响,首先会表现在行为上的变化[31]. 水蚤在溴酸盐暴露后首先出现快速游动,无方向性,在烧杯底部打转,随后游动缓慢、 触角摆动变慢等现象,最后失去游动能力,沉入烧杯底部,直至死亡. 并且随着溴酸盐浓度的增加,死亡率逐渐升高,而对照组水蚤则未出现任何不正常现象.
表 2和表 3为不同暴露时间下溴酸钾、 溴酸钠、 溴化钾对大型蚤和裸腹蚤的EC50和LC50. 对比3种污染物对这两种水蚤的EC50和LC50,可以发现溴酸钾和溴酸钠对水蚤的EC50和LC50远远小于溴化钾的数值,这说明阳离子的差异所产生的影响远小于阴离子差异的影响,因此造成水蚤运动受抑制和死亡的原因是由溴酸盐(即BrO3-)所引起的. Hutchinson等[32]对前人研究的综述发现溴酸盐对大型蚤48 h LC50为179 mg ·L-1(以BrO3-计),此结果与本研究结果相近,而Fisher等[28]的报道为溴酸钠对大型蚤的24 h和48 h LC50分别为112.7 mg ·L-1和55.3 mg ·L-1,此结果与本研究结果有一定差异,这可能是由试验方法的差异造成的. 从表 2和表 3中可以发现溴酸钾和溴酸钠对大型蚤和裸腹蚤EC50和LC50数值相差不大,这说明大型蚤和裸腹蚤对溴酸盐的毒性敏感度相近. 同时对于两种水蚤而言,48 h的EC50和LC50均小于24 h的EC50和LC50,表明这两种蚤在溴酸盐的暴露下,毒性效应随着时间的延长越大,蚤的耐受程度越低.
| 表 2 污染物对大型蚤的急性毒性 Table 2 Acute toxicity of KBrO3,NaBrO3 and KBr on Daphnia magna |
| 表 3 污染物对裸腹蚤的急性毒性 Table 3 Acute toxicity of KBrO3,NaBrO3 and KBr on Moina |
斑马鱼基因与人类基因相似度高达87%,有关化学物质在斑马鱼体内得到的毒理学研究结果在多数情况下也适用于人体环境[33],同时作为水生生态系统的消费者,水体中的污染物也会严重影响到鱼类. 因此,在研究水生生态毒理学时开展对鱼类毒性试验也是非常必要的.
当斑马鱼与溴酸盐接触后,中毒症状表现为首先快速游动,呼吸加快,随后失去平衡,侧翻,摆动频率和游动速度减慢,最后沉入水底死亡,而对照组斑马鱼则未出现任何不正常现象.
表 4为不同暴露时间下溴酸钾、 溴酸钠、 溴化钾对斑马鱼的LC50. 通过对斑马鱼的LC50的对比,可以发现虽然溴酸钾和溴酸钠对斑马鱼的LC50有所区别,但都远远小于溴化钾的数值,因此说明造成斑马鱼毒性效应的原因是由溴酸盐(即BrO3-)所引起的. 同时随着暴露时间的增加,LC50降低,说明溴酸盐对斑马鱼的暴露时间越长,所表现出的毒性效应就越大,斑马鱼的耐受程度越低.
| 表 4 污染物对斑马鱼的急性毒性 Table 4 Acute toxicity of KBrO3,NaBrO3 and KBr on zebrafish |
通过以上结果和分析表明,造成水生生物生长速度变慢、 运动受抑制或死亡率增加的原因主要是由溴酸根离子,即溴酸盐引起的,并且随着溴酸盐浓度的升高,相应的生长速度或存活率也随之下降. 同时溴酸盐暴露的时间越久,表现出的毒性效应越明显. 对比可知,在所考察的水生生态系统中的几个营养级的受试生物对溴酸盐的毒性效应的敏感性差别很大,即使是同一营养级也会有差别. 其中大型蚤和裸腹蚤最敏感,斜生栅藻次之,然后是斑马鱼,而普通小球藻和发光菌最不敏感.
一定浓度的化学物质可能对某一营养级某种生物的生存没有毒害作用,但可能对另一个营养级的某种生物的生存存在强烈的毒害作用[34],因为水生态系统的稳定性和功能与每一个营养级上的生物都密切相关,因此仅依赖一种生物的毒性研究结果对溴酸盐进行毒性分级和探讨其对水生生态系统的毒性效应是片面的. 因此,本研究综合考虑包括发光菌、 浮游植物、 浮游动物、 鱼等多营养级水生生物来测定和评估溴酸盐的毒性效应,为环境中溴酸盐的毒性作用特征和风险评价提供了更有效、 全面的信息.
3 结论(1)溴酸钾和溴酸钠对斜生栅藻、 大型蚤、 裸腹蚤和斑马鱼的EC50和LC50数值远小于溴化钾的数值,造成绿藻生长受到抑制,大型蚤和斑马鱼活动受到抑制或死亡的原因主要是由溴酸盐(即BrO3-)的毒性效应引起的.
(2)斜生栅藻、 大型蚤、 裸腹蚤和斑马鱼的EC50或LC50随着暴露在溴酸盐中时间的增加呈下降趋势,反映溴酸盐的毒性效应随时间增大,斜生栅藻、 大型蚤、 裸腹蚤和斑马鱼对溴酸盐毒性效应的耐受程度随时间降低.
(3) 不同受试生物对溴酸盐的毒性效应的敏感性不同. 其中大型蚤和裸腹蚤最敏感,斜生栅藻次之,然后是斑马鱼,而普通小球藻和发光菌最不敏感.
| [1] | 安东, 李伟光, 崔福义, 等. 溴酸盐的生成及控制[J]. 水处理技术, 2005, 31 (6): 54-55, 78. |
| [2] | 李宁. 溴酸钾的安全性毒理学研究进展[J]. 国外医学卫生学分册, 2006, 33 (1): 44-46. |
| [3] | Nakajima M, Kitazawa M, Oba K, et al. Effect of route of administration in the micronucleus test with potassium bromate[J]. Mutation Research/Genetic Toxicology, 1989, 223 (4): 399-402. |
| [4] | 王伟, 蒋颂辉, 朱惠刚, 等. 溴酸盐的遗传毒性[J]. 环境与健康杂志, 2003, 20 (3): 137-138. |
| [5] | 沈明浩, 王月娇, 李嘉, 等. 溴酸钾的毒理学研究[J]. 食品科学, 2008, 29 (8): 591-593. |
| [6] | Ishidate Jr M, Sofuni T, Yoshikawa K, et al. Primary mutagenicity screening of food additives currently used in Japan[J]. Food and Chemical Toxicology, 1984, 22 (8): 623-636. |
| [7] | Suzuki T, Hayashi M, Hakura A, et al. Combination effects of clastogens in the mouse peripheral blood micronucleus assay[J]. Mutagenesis, 1995, 10 (1): 31-36. |
| [8] | GB 5749-2006, 生活饮用水卫生标准[S]. |
| [9] | GB 19298-2014, 食品安全国家标准 包装饮用水[S]. |
| [10] | Weinberg H S, Delcomyn C A, Unnam V. Bromate in chlorinated drinking waters: Occurrence and implications for future regulation[J]. Environmental Science & Technology, 2003, 37 (14): 3104-3110. |
| [11] | Butler R, Godley A, Lytton L, et al. Bromate environmental contamination: Review of impact and possible treatment[J]. Critical Reviews in Environmental Science and Technology, 2005, 35 (3): 193-217. |
| [12] | Kruithof J C, Meijers R T. Bromate formation by ozonation and advanced oxidation and potential options in drinking water treatment[J]. Water Supply, 1995, 13 (2): 93-103. |
| [13] | Anupama V N, Kannan K, Prajeesh P V G, et al. Perchlorate, chlorate and bromate in water samples from the South-West coast of India[J]. Water Science and Technology-Water Supply, 2012, 12 (5): 595-603. |
| [14] | Kemsley J. Bromate in Los Angeles water[J]. Chemical & Engineering News, 2007, 85 (52): 9. |
| [15] | Genuino H C, Espino M P B. Occurrence and sources of bromate in chlorinated tap drinking water in Metropolitan Manila, Philippines[J]. Archives of Environmental Contamination and Toxicology, 2012, 62 (3): 369-379. |
| [16] | Wu Q, Zhang T, Sun H W, et al. Perchlorate in tap water, groundwater, surface waters, and bottled water from China and its association with other inorganic anions and with disinfection byproducts[J]. Archives of Environmental Contamination and Toxicology, 2010, 58 (3): 543-550. |
| [17] | 刘勇建, 牟世芬, 林爱武, 等. 北京市饮用水中溴酸盐、 卤代乙酸及高氯酸盐研究[J]. 环境科学, 2004, 25 (2): 51-55. |
| [18] | 杨永亮, 刘崴, 刘晓端, 等. 辽宁省西部和沈阳地区河水及地下水中溴的分布与污染特征[J]. 环境化学, 2009, 28 (6): 924-928. |
| [19] | GB/T 21805-2008, 化学品 藻类生长抑制试验[S]. |
| [20] | GB/T 13266-1991, 水质 物质对蚤类(大型蚤)急性毒性测定方法[S]. |
| [21] | GB/T 13267-1991, 水质 物质对淡水鱼(斑马鱼)急性毒性测定方法[S]. |
| [22] | 张泽光, 黄满红, 陈东辉, 等. 十溴联苯醚对大型蚤和发光菌的毒性研究[J]. 环境工程, 2013, 31 (增刊): 299-302. |
| [23] | Wen Y Z, Yuan Y L, Chen H, et al. Effect of chitosan on the enantioselective bioavailability of the herbicide dichlorprop to Chlorella pyrenoidosa[J]. Environmental Science & Technology, 2010, 44 (13): 4981-4987. |
| [24] | 徐冬梅, 王艳花, 饶桂维. 四环素类抗生素对淡水绿藻的毒性作用[J]. 环境科学, 2013, 34 (9): 3386-3390. |
| [25] | Parr L B, Perkins R G, Mason C F. Reduction in photosynthetic efficiency of Cladophora glomerata, induced by overlying canopies of Lemna spp.[J]. Water Research, 2002, 36 (7): 1735-1742. |
| [26] | Ma J Y, Zheng R Q, Xu L G, et al. Differential sensitivity of two green algae, Scenedesmus obliqnus and Chlorella pyrenoidosa, to 12 pesticides[J]. Ecotoxicology and Environmental Safety, 2002, 52 (1): 57-61. |
| [27] | 冯鸣凤, 赵娜, 王轲, 等. 不同pH下对硝基酚(p-NP)对小球藻和斜生栅藻的毒性[J]. 环境科学研究, 2011, 24 (2): 210-215. |
| [28] | Fisher D, Yonkos L, Ziegler G, et al. Acute and chronic toxicity of selected disinfection byproducts to Daphnia magna, Cyprinodon variegatus, and Isochrysis galbana[J]. Water Research, 2014, 55 : 233-244. |
| [29] | 王良韬, 吴永贵, 廖芬, 等. Cu2+对水生食物链关键环节生物的毒性效应[J]. 贵州农业科学, 2011, 39 (5): 226-230. |
| [30] | 吴永贵, 黄建国, 袁玲. 利用水溞的趋光行为监测水质[J]. 中国环境科学, 2004, 24 (3): 336-339. |
| [31] | Fernandezcasalderrey A, Ferrando M D, Andreumoliner E. Effect of sublethal concentrations of pesticides on the feeding behavior of Daphnia magna[J]. Ecotoxicology and Environmental Safety, 1994, 27 (1): 82-89. |
| [32] | Hutchinson T H, Hutchings M J, Moore K W. A review of the effects of bromate on aquatic organisms and toxicity of bromate to oyster (Crassostrea gigas) embryos[J]. Ecotoxicology and Environmental Safety, 1997, 38 (3): 238-243. |
| [33] | 王佳佳, 徐超, 屠云杰, 等. 斑马鱼及其胚胎在毒理学中的实验研究与应用进展[J]. 生态毒理学报, 2007, 2 (2): 123-135. |
| [34] | 牧辉, 彭新晶, 戴宁, 等. 离子液体[C8mim]PF6对水生生物的毒性作用[J]. 中国环境科学, 2009, 29 (11): 1196-1201. |