 2015, Vol. 36
2015, Vol. 36

2. 贵州师范大学地理与环境科学学院, 贵阳 550001;
3. 贵州师范大学化学与材料学院, 贵阳 550001;
4. 贵州省水文水资源局, 贵阳 550002
 , LI Qiu-hua1
, LI Qiu-hua1 , JIAO Shu-lin2, LI Yue1, XIAO Jing1, DENG Long1, SUN Rong-guo3, GAO Yong-chun4, LUO Lan4
, JIAO Shu-lin2, LI Yue1, XIAO Jing1, DENG Long1, SUN Rong-guo3, GAO Yong-chun4, LUO Lan4
2. School of Geography and Environmental Science, Guizhou Normal University, Guiyang 550001, China;
3. School of Chemistry and Material Science, Guizhou Normal University, Guiyang 550001, China;
4. Hydrology and Water Resources Bureau of Guizhou Province, Guiyang 550002, China
水库、 湖泊水体富营养化逐年加剧,目前已成为水环境保护突出的问题[1],浮游植物作为水生态系统生产过程中的最初和基础环节,其群落的变化可以反映水体的营养状况和生态环境质量的变化[2],传统浮游藻类鉴定根据物种同源性特点将其归入不同的类群,很少体现环境生态特征[3],具有一定局限性. Reynolds等[4]于2002年提出相对完整的浮游藻类功能群理论,后经Padis k等[5]补充完善迄今共鉴定并描述了39种浮游藻类功能群. 目前功能群在浮游植物生态学中已成为国内外研究的热点问题[6, 7, 8, 9, 10, 11, 12]. 然而运用功能群对小型水库进行浮游植物群落结构分析与富营养化评价的研究还比较少,小型水体更易受外界物理因子及人为活动干扰,功能群能否正确评价这类水体的富营养化特征并做出响应?这类水体出现富营养化后浮游植物功能群分布与环境因子关系有何特殊性?本研究将一一进行分析探讨. 本研究基于对小关水库夏季水质理化参数以及浮游植物的研究近况,结合功能群分类特征对其浮游植物进行功能群划分,分析在富营养化、 水动力等条件下浮游植物功能群的分布特征及其影响因子以及功能群对富营养化特征的响应问题,并采用藻类生物学评价法、营养状态指数法以及浮游植物功能群生境指示法对小关水库富营养化进行综合评价,以期为贵州高原水库浮游植物研究及水资源管理提供理论基础.
1 材料与方法 1.1 水库概况及采样点设置小关水库地处贵阳市云岩区,属亚热带湿润型季风气候,年均气温15.3℃,是一座集水面积约29.5 km2,总库容达到2.31×106 m3的小型浅水水库. 其功能是防洪以及向黔灵湖与南明河提供环境用水,自2004年以来,政府已投入上千万资金通过清淤、 对水库水面进行日常保洁、 投放鱼类、 设置人工植物浮岛等进行水生态治理,以改善小关水库的水环境状况,然而收效甚微,近年来富营养化仍然严重,已出现多次水华,其生态状况令人担忧.
于2014年7月25日至9月27日进行1周1次、 为期10周的采样,根据小关水库水域特点将采样点设在水库大坝(E106°41′26.52″,N26°37′34.27″)和上游(E106°41′30.17″,N26°37′56.71″)(图 1).
 |
图 1 小关水库样点设置示意 Fig. 1 Sampling sites in Xiaoguan Reservoir |
浮游植物定性样品用25号浮游生物网于水平及垂直方向“∞”字形缓慢拖网,浮游植物定量样品于水下0.5 m处取1.5 L水样,带回实验室用3%~5%福尔马林固定,于实验室进行镜检,藻种鉴定与分类方法参考文献[13]. 水温(WT)、 电导率(EC)、 溶解氧(DO)以及pH用YSI多参数水质检测仪现场测定,通过赛氏盘(Secchi Disk) 对透明度(SD)进行测定,按照GB 3838-2002《地表水环境质量标准方法》测定总磷(TP)、 总氮(TN)、 氨氮(NH4+-N)、 硝氮(NO3--N)、 亚硝氮(NO2--N)与正磷酸盐(PO43--P)等指标. 用0.45 μm的醋酸纤维滤膜对叶绿素a(Chla)进行抽滤,水样为500 mL,反复冻融浸提,运用丙酮萃取方法测定[14].
1.3 数据分析方法 1.3.1 单因素方差分析对采样点大坝与上游的综合营养指数、 环境因子进行单因素方差分析(one-way ANOVA),于SPSS 19.0软件下完成.
1.3.2 藻类生物学法根据湖泊营养类型评价的藻类生物学指标与标准[15],藻类丰度(D) > 108cells ·L-1时为重度富营养状态,80×106 cells ·L-1<D≤108 cells ·L-1时为富营养状态,40×106 cells ·L-1<D≤80×106 cells ·L-1时为中度富营养,10×106 cells ·L-1<D≤40×106 cells ·L-1时为中营养状态,D≤106 cells ·L-1为贫营养状态. Chla浓度 < 25 μg ·L-1时为中营养,Chla浓度 < 50μg·L-1时为中富营养状态,Chla浓度 < 500 μg ·L-1时为富营养状态.
1.3.3 综合营养状态指数采用综合营养状态指数法评价水体营养状态,其公式为:
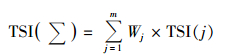
式中,TSI(Σ)为综合营养状态指数; Wj为第j种参数的营养状态指数的相关权重; TSI(j)代表第j种参数的营养状态指数; m为评价参数的个数.
1.3.4 数据处理采用SPSS计算综合营养指数TLI值并进行单因素方差分析. 小关水库的浮游植物物种数据与环境因子的RDA分析在Canoco 4.5软件包上运行,通过浮游植物常见功能群丰度指标来反映浮游植物种类与环境因子之间的关系,为获得数据正态分布,将物种数据以及环境因子(pH除外)数据进行lg(x+1)转换. 其他图表均由Excel绘制.
2 结果与分析 2.1 浮游植物群落结构及功能群划分调查中,共鉴定出浮游植物78种(包括变种、 变型),隶属于7个门. 其中绿藻门43种,蓝藻门16种; 硅藻门11种; 裸藻门3种; 甲藻门2种; 隐藻门1种; 金藻门2种. 其中绿藻门最多,占总数的55.1%; 其次为蓝藻门和硅藻门,分别占20.5%、 14.1%,其他几门相对较少. 此外,蓝藻占总丰度的15.4%~98.2%,均值为84.6%,绿藻占总丰度的1.89%~65.5%,均值为9.91%,硅藻占总丰度的0.87%~38.1%,均值为9.76%(图 2). 可知小关水库夏季浮游植物群落结构为蓝-绿-硅藻型. 参考Reynold等提出的功能群分类方法[4,5],结合水库的水质及生态状况,划分出A、 B、 D、 E、 F、 G、 H1、 J、 LM、 LO、 M、 MP、 P、 S1、 SN、 T、 W1、 W2、 X1、 X3、 Xph、 Y共计22个功能群. 频率分析发现,功能 群S1、 D、 J、 B、 G、 MP、 LO、 SN、 X1、 Y、 Xph、 F、 T、 W1在水体中出现频率达到50%以上,为这次调查中的常见功能群,W2、 E、 LM出现频率在30%~50%之间,只在适宜条件下出现,而P、 A、 H1、 X3、 M出现几率较低. 如果某个功能类群在至少1个采样点的相对丰度比例>10%,则达到优势地位,称之为优势功能群,分别为S1、D与J这3个功能群. 根据浮游植物功能群对物质和能量供给变化划分其C-R-S生长策略,C型在物质和能量均充分的条件下占优,R型对能量限制的情况下具有耐受性,S型在物质限制下占优. 各优势功能群的代表性藻种(属)及其C-R-S生长策略见表 1.
 |
图 2 小关水库浮游植物功能群频率分布 Fig. 2 Frequency distribution of phytoplankton functional groups in Xiaoguan Reservoir |
| 表 1 浮游植物常见功能群的代表性藻种及其C-R-S生长策略 Table 1 Phytoplankton functional groups and their growth strategies |
大坝功能类群动态变化主表现为S1→S1→S1→S1→S1→S1→S1→J/D/S1→S1→S1/D,上游功能类群动态变化主要过程为S1→S1→S1→S1→S1→S1→S1→J/D/S1→S1→S1(图 3~4),两者变化趋势大体相同. 从相对丰度上看,优势功能群S1、 J、 D占总丰度的84.3%以上. 其中蓝藻门的功能群S1(以湖泊假鱼腥藻为主)占绝对优势,除了于第8周仅为15.0%之外,其余时期均在74.4%以上,最高值达到91.1%,而绿藻门的功能群J仅在第8周占绝对优势,相对丰度为65.1%,硅藻门的功能群D也在第8周占优,相对丰度为13.9%,此外于第10周的大坝再次占据优势,达到14.2%. 调查期间(图 5),绝对优势功能群S1丰度表现出一定的规律性. 采样点大坝与上游变化趋势基本保持一致,均从第1周至第4周缓慢增长,于第5周出现第一次峰值,第6周有所下降,第7周出现第二次峰值,并于第二次峰值之后的第8周滑入最低谷(此时正值秋初),稍后又有所增长,但丰度上与两次峰值相比已从108 cells ·L-1下降至107 cells ·L-1.
 |
图 3 大坝浮游植物优势功能群周变化 Fig. 3 Weekly dynamics of dominant phytoplankton functional groups in the Dam site |
 |
图 4 上游浮游植物优势功能群周变化 Fig. 4 Weekly dynamics of dominant phytoplankton functional groups in the upstream site |
 |
图 5 功能群S1周变化 Fig. 5 Weekly dynamics of S1 in Xiaoguan Reservoir |
小关水库的理化因子见表 2,对大坝与上游的环境因子进行单因素方差分析,结果均显示P > 0.05,即差异不显著,表明小关水库水体交换能力较强. 采样期间,TN、 TP营养盐平均值分别为1.785 mg·L-1与0.078 mg ·L-1,远高于国际上一般标准的水体富营养化发生的质量浓度. 此外,溶解性氮、 磷值均表现出较大的变化趋势. 由于采样时间为夏季,WT较高,其均值达到24.83℃,其变化范围相对较小. SD在0.500~0.950 m范围内变化,总体表现较低. Chla表现出较高的质量浓度,最高值达到135.3 μg ·L-1,其变化波动较明显. DO最低值、 最高值相差7.18 mg ·L-1,波动幅度也较大. 此外,EC也有较大的波动,而pH值变化幅度较小.
| 表 2 小关水库水质理化指标 Table 2 Physico-chemical parameters of water quality in Xiaoguan Reservoir |
采用Chla、 TP、 TN、 SD作为水体综合营养状态评价指标. 通过小关水库各个理化指标参数可得到各指标的营养状态指数,然后通过综合营养状态指数对水库的水质进行富营养化评价. 利用SPSS 19.0对采样点大坝与上游的综合营养指数进行单因素方差分析(one-way ANOVA),得F=2.23,P>0.05,即差异不显著,因此对两采样点的综合营养指数取平均值,不再分开讨论分析. 根据中国环境监测总站有关湖泊(水库)营养状态分级规定,50 < TLI(Σ) ≤60为轻度富营养,60 < TLI(Σ) ≤70为中度富营养,小关水库综合营养状态指数基本处于中度富营养状态(见图 6).
 |
图 6 小关水库综合营养指数周变化 Fig. 6 Weekly dynamics of the comprehensive trophic state index(TLI) in Xiaoguan Reservoir |
对物种数据进行除趋势对应分析(DCA)选择排序模型,运用Canoco 4.5软件分析得出:最大梯度长是0.704,故对数据进行冗余分析(RDA). 对物种数据进行筛选,选择14个常见功能群用于排序. 环境因子包括TP、 TN、 NO3--N、 NO2--N、 NH4+-N、 PO43--P、 WT、 DO、 pH、 EC、 N/P、 SD、 Chla共11个,采用蒙特卡拟合方法对物种数据及环境因子进行显著性检验. 通过筛选,仅有SD、 Chla两个具有显著解释性的环境变量,与物种变量进行RDA分析(图 7).结果显示:第1轴的特征值达到0.696,说明第1轴能解释绝大部分的浮游植物分布. 环境因子SD、 Chla与轴1的相关系数分别为-0.804、 0.727,与浮游植物分布关系密切. Chla与功能群S1、 J、 SN等大部分功能群呈正相关,表明浮游植物丰度与Chla呈正相关,其中S1、 SN、 T、 F、 W1、 LO对Chla贡献较大,然而除功能群D之外,透明度与功能群S1、 J、 SN等大部分功能群呈负相关.
 |
图 7 浮游植物功能群丰度与环境因子的RDA分析 Fig. 7 Redundancy analysis of phytoplankton functional groups related to environmental factors |
湖泊富营养化评价主要有特征法、 参数法、 藻类生物学法、 营养状态指数法、 灰色系统法及应用模糊数学等系列方法,对湖泊富营养化进行单一的方法评价往往不够客观,缺乏说服力,因此本研究中主要采用藻类生物学评价中的丰度指标、 叶绿素指标,营养状态指数法以及浮游植物功能群几种方法对小关水库富营养化进行综合评价. 在10次采样中,浮游植物丰度变化范围在2.59×107~5.10×108 cells·L-1之间,其中5次为极富营养状态,4次为中富营养状态,一次为中营养状态; Chla浓度最低值为44.7 μg·L-1,最高值为128.6 μg·L-1,9次处于富营养状态,1次处于中富营养状态,富营养状态占绝大多数; 综合营养状态指数在55.76~65.81间变动,9次为中度富营养,一次为轻度富营养,表明对大多数处于中度富营养状态; 优势功能群S1、 J常生长于浑浊的富营养化水体,D在浑浊的,含有营养盐的水体中生长良好并对营养盐缺乏敏感. 功能群B生长于中营养水体,G生长于富营养水体,MP适合生长于浑浊的浅水水体,LO于各种营养等级水体均能生长,SN生长于混合水体,X1、 W1均适合于高富营养水体生长,功能群Y于静止水体生长良好,Xph生长于含钙高,碱性的小型水体,对低营养敏感,F生长于中到富营养水体,T生长于混合水体,对营养缺乏敏感[5]. 以上常见功能群基本于富营养化水体生长良好,且大都属于C、 R与C/R型策略功能群,在理想的物质和能量供给条件下能在竞争中获胜而取代其他藻种,因此由功能群可以判定小关水库处于富营养化状态,这与叶绿素指标评价以及综合营养状态指数法评价结果基本一致. 由于该水库假鱼腥藻在丰度上占据绝对优势,然而作为小型丝状藻种,假鱼腥藻的细胞体积远低于大多数藻类,在生物量上不占优势,因此以藻类丰度作为判定水体富营养化的标准在本研究中并不适用,容易出现评价结果偏大,不能够真实反映水体水质情况的问题,可以滤选掉. 通过对藻类生物学叶绿素指标、 营养状态指数以及浮游植物功能群这3种方法的综合评价,得出小关水库处于中度富营养-富营养化状态. 值得一提的是,营养盐、 丰度、 叶绿素各项指标均已超过水体发生水华的最低阈值,然而该水库并未发生水华,这可能与降雨、 水体混合以及假鱼腥藻为该水库绝对优势种相关,当然这需要更多的数据分析加以明确.
3.2 浮游植物功能群对水库富营养化特征的响应水生生态系统中浮游植物种类和数量与水体环境之间存在着紧密关系[16]. 小关水库作为一座小型浅水水库,且处于富营养化状态,浮游植物功能群与环境因子的关系具有其独特性. RDA分析表示SD是浮游植物优势功能群分布主要影响因子. 虽然RDA分析中,WT与浮游植物分布相关性不显著,但由于本次调查时间在夏季,WT较高,适合蓝藻和绿藻生长,而蓝、 绿藻又为本次调查的优势藻种,因此WT也是影响浮游植物种类组成和数量的一个主要因素. Chla是浮游植物重要的组成成分之一,与浮游植物的种类、 丰度以及生物量密切相关,是反映水质的综合评价指标,一般而言,浮游植物丰度越高,Chla质量浓度越高,所以本研究中Chla与功能群丰度呈显著正相关不能说明其是影响浮游植物分布的主要影响因子,故影响浮游植物优势功能群分布的主要因子为WT与SD. 虽然该水库常见功能群大都适于富营养水体生长,但未分析出氮、 磷营养盐与浮游植物功能群分布呈显著相关关系,可见富营养化的小关水库中,较高的氮磷营养盐不再成为浮游植物功能群分布的限制因子,这与文献[17]的研究结果一致.
S1功能类群适应生境为浑浊的混合水体环境[5],且于中富营养水体中生长良好[10],主要的代表种类是湖泊假鱼腥藻. 国内外水体研究中以S1类为优势功能群的现象普遍存在[18, 19, 20, 21, 22]. 近年来贵州高原有关假鱼腥藻优势种的报道也越来越多,如贵阳市内的红枫湖水库、 百花湖以及黔东南的三板溪水库[10, 23, 24]. 最近有研究表明假鱼腥藻成为优势种的水体不容易发生蓝藻水华[21, 22]. 本研究中功能群S1在丰度上占据绝对优势,这可能是因为夏季气温较高,适于假鱼腥藻等蓝藻生长,且多强对流暴雨天气,大量外源物质对库区营养盐进行补充,同时造成水体浊度升高,透明度降低,进而光限制增强,而S1作为典型的R型策略功能群,对能量输入(光照辐射)具有较高的耐受性[25],此外该水库属于浅水水体,水体上下层水温差异较小,不易产生分层,同时易受风力,降雨等物理因子干扰,使得水体混合良好,十分适合S1生长,成为绝对优势功能群. 然而功能群S1在最后几周的秋季采样中其丰度与优势度均有所下降,第8周采样优势度已跌破至15.0%,且在镜检中,大量假鱼腥藻处于分解状态. 与前几周相比,这次水库水位明显下降,据此推测在此期间小关水库工作人员为了给下游的黔灵湖与南明河补充水源,进行了一次大规模的开闸放水,强烈的冲刷作用导致了假鱼腥藻的大量死亡. 功能群J适应生境为浅的、 混合的高富营养水体[5]. 该水库中功能群J的主要代表种类是栅藻,盘星藻与空星藻. 虽然仅于第8周采样中占据绝对优势,但其包含的种类最多,达到13种,占所有总类数的16.7%,且出现频率达到100%,为第二优势功能群. 其在该水库广泛存在可能是因为流域内的云岩区雅关村、 偏坡村,高新区的大关村境内,存在许多养猪大户、 豆腐作坊,这些场所产生的猪粪、 废水未经过处理排进库区,带入大量有机质,造成水体高度富营养化,十分利于其生长. 此外,J作为CR型策略功能群,在理想的物质和能量供给条件下能在竞争中获胜而取代其他藻种直至达到顶级状态,并且对能量输入(光照辐射)具有较高的耐受性[25]. Romo等发现[26],S1与J常共生于浑浊的富营养化水体中,这在本研究中得到了验证. D功能群耐受冲击,对营养耗尽敏感,适应生境为浅的浑浊水体[5],主要优势种为硅藻门的针杆藻. 作为R型策略功能群,对能量输入(光照辐射)具有较高的耐受性[25],其本身在夏季就具有其优越性,此外,小关水库下游为黔灵湖及南明河,为满足市民生活、 旅游、 休闲等需求,库区需要不时进行开闸放水,造成水力滞留时间短,从而利于耐冲刷的功能群D占据优势,而这种优势在大坝采样点更为明显. 值得一提的是,此次采样中在贵州高原鉴定出了一种能指示喀斯特水库多钙且碱度高的功能群Xph,代表性藻种为透镜壳衣藻. 其生境为小型碱性湖泊,钙含量丰富且光照充足[5],调查中,出现频率达到60%. 作为典型的高原喀斯特小型水库,小关水库也具有含钙高,呈碱性的特点,然而该水库夏季浊度高,透明度低,这也一定程度上影响了光照强度,限制了Xph的生长.
从小关水库优势功能群的出现及其演变上看,整体上变化不大. 从第1周到第7周两采样点均以S1为绝对优势功能群,第8周起开始出现大的变化,功能群J/D/S1共同占据优势地位,其后又回到以S1为绝对优势功能群的局面. 采样期间,均以功能群S1(第8周除外)为绝对优势功能群,而导致优势功能群于第8周出现大的变化则主要是因为这次大规模的开闸放水,水力冲刷作用变强,对冲刷敏感的功能群S1大量死亡,而这却为适宜浅水水体的功能群J以及耐冲刷的功能群P提供了条件. 据Padisk等学者研究表明[5],功能群J适宜于低梯度变化的河流,这是否也能说明功能群J对冲刷也具有一定的耐受性,这在文中得到了一定的论证. 总体来说,功能群S1、 J以及D的适宜生境均为浑浊或混合的富营养性水体,小关水库在夏季恰好具备了这样的生境条件. 小关水库夏季处于中度富营养-富营养化状态,而该时期降水丰富且多暴雨,小型水体更易受降雨等物理因子影响,上游带来的小量洪水也足以使库区水体变得浑浊. 因此,功能群可以对浅水水体的富营养化特征以及水动力条件做出有效响应.
4 结论(1)小关水库共检出22个功能类群,分别为: A、 B、 D、 E、 F、 G、 H1、 J、 LM、 LO、 M、 MP、 P、 S1、 SN、 T、 W1、 W2、 X1、 X3、 Xph、 Y. 其中功能群S1、 D、 J、 B、 G、 MP、 LO、 SN、 X1、 Y、 Xph、 F、 T、 W1为水库常见功能群,S1、 J、 D为优势功能群. 其生长策略主要表现为C/R/CR型.
(2)经浮游植物功能群、藻类生物学以及综合营养指数综合评价得出小关水库处于中度富营养-富营养化状态,运用浮游植物功能群对小型水库进行富营养化评价同样适用.
(3)该水库营养盐含量丰富,不再成为影响浮游植物功能群分布的限制因子,反而是透明度、 水温成为影响浮游植物功能群分布的关键因子.
(4)浮游植物功能群可以对浅水水体的富营养化特征做出有效响应.
| [1] | Zhu W, Wan L, Zhao L F. Effect of nutrient level on phytoplankton community structure in different water bodies[J]. Journal of Environmental Sciences, 2010, 22 (1): 32-39. |
| [2] | Liu C Q, Liu L S, Shen H T. Seasonal variations of phytoplankton community structure in relation to physico-chemical factors in Lake Baiyangdian, China[J]. Procedia Environmental Sciences, 2010, 2 : 1622-1631. |
| [3] | 董静, 李艳晖, 李根保, 等. 东江水系浮游植物功能群季节动态特征及影响因子[J]. 水生生物学报, 2013, 37 (5): 836-843. |
| [4] | Reynolds C S, Huszar V, Kruk C, et al. Towards a functional classification of the freshwater phytoplankton[J]. Journal of Plankton Research, 2002, 24 (5): 417-428. |
| [5] | Padisák J, Crossetti L O, Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates[J]. Hydrobiologia, 2009, 621 (1): 1-19. |
| [6] | Becker V, Huszar V L M, Crossetti L O. Responses of phytoplankton functional groups to the mixing regime in a deep subtropical reservoir[J]. Hydrobiologia, 2009, 628 (1): 137-151. |
| [7] | Abonyi A, Leitão M, Lançon A M, et al. Phytoplankton functional groups as indicators of human impacts along the River Loire (France)[J]. Hydrobiologia, 2012, 698 (1): 233-249. |
| [8] | Demir A N, Fakio Dğ lu Ö, Dural B. Phytoplankton functional groups provide a quality assessment method by the Q assemblage index in Lake Mogan (Turkey)[J]. Turkish Journal of Botany, 2014, 38 (1): 169-179. |
| [9] | Stevi Dć F, Mihaljevi Dć M, Špoljari Dć D. Changes of phytoplankton functional groups in a floodplain lake associated with hydrological perturbations[J]. Hydrobiologia, 2013, 709 (1): 143-158. |
| [10] | 卢金锁, 胡亚潘. 深水型水库藻类功能组时空演替及生境变化的影响[J]. 环境科学, 2013, 34 (7): 2611-2617. |
| [11] | 黄国佳, 李秋华, 陈椽, 等. 贵州高原红枫湖水库浮游植物功能分组及其时空分布特征[J]. 生态学报, 2015, 35 (17): 1-14. |
| [12] | 胡韧, 蓝于倩, 肖利娟, 等. 淡水浮游植物功能群的概念、划分方法与应用[J]. 湖泊科学, 2015, 27 (1): 11-23. |
| [13] | 胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [14] | Lin S J, He L J, Huang P S, et al. Comparison and improvement on the extraction method for chlorophyll a in phytoplankton[J]. Ecologic Science, 2005, 24 (1): 9-11. |
| [15] | 况琪军, 马沛明, 胡征宇, 等. 湖泊富营养化的藻类生物学评价与治理研究进展[J]. 安全与环境学报, 2005, 5 (2): 87-91. |
| [16] | Götzenberger L, de Bello F, Bråthen K A, et al. Ecological assembly rules in plant communities—approaches, patterns and prospects[J]. Biological Reviews, 2012, 87 (1): 111-127. |
| [17] | 方丽娟, 刘德富, 杨正健等. 三峡水库香溪河库湾夏季浮游植物演替规律及其原因[J]. 生态与农村环境学报, 2013, 29 (2): 234-240. |
| [18] | Soylu E N, Gönülol A. Functional classification and composition of phytoplankton in Liman Lake[J]. Turkish Journal of Fisheries and Aquatic Sciences, 2010, 10 (1): 53-60. |
| [19] | Borics G, Tóthmérész B, Lukács B A, et al. Functional groups of phytoplankton shaping diversity of shallow lake ecosystems[J]. Hydrobiologia, 2012, 698 (1): 251-262. |
| [20] | 张婷, 李林, 宋立荣. 熊河水库浮游植物群落结构的周年变化[J]. 生态学报, 2009, 29 (6): 2971-2979. |
| [21] | Tian C, Lu X T, Pei H Y, et al. Seasonal dynamics of phytoplankton and its relationship with the environmental factors in Dongping Lake, China[J]. Environmental Monitoring and Assessment, 2013, 185 (3): 2627-2645. |
| [22] | Lu X T, Tian C, Pei H Y, et al. Environmental factors influencing cyanobacteria community structure in Dongping Lake, China[J]. Journal of Environmental Sciences, 2013, 25 (11): 2196-2206. |
| [23] | 黄志敏, 陈椽, 刘之威, 等. 贵州百花湖夏季浮游植物昼夜垂直分布特征[J]. 生态学报, 2014, 34 (19): 5389-5397. |
| [24] | 黄国佳, 李秋华, 陈椽, 等. 贵州高原三板溪水库浮游植物功能群时空分布特征[J]. 环境科学学报, 2015, 35 (2): 418-428. |
| [25] | Reynolds C S. The ecology of freshwater phytoplankton[M]. Cambridge: Cambridge University Press, 2006. |
| [26] | Romo S, Villena M J. Phytoplankton strategies and diversity under different nutrient levels and planktivorous fish densities in a shallow Mediterranean lake[J]. Journal of Plankton Research, 2005, 27 (12): 1273-1286. |