 2015, Vol. 36
2015, Vol. 36



2. 挪威生命科学大学环境科学学院,奥斯 NO-1432,挪威
2. Department of Environmental Sciences, Norwegian University of Life Sciences, s NO-1432, Norway
全球范围内大气活性态氮(Nr,主要包括氮氧化物NOx和氨NH3)排放量的持续增加,导致欧洲、 北美和东亚许多地区很高的大气氮沉降量[1]. 氮素是森林生态系统的主要营养元素之一,较低的氮沉降能促进森林的生长; 但当氮沉降超过生态系统可以利用的限度时,就会发生氮饱和[2],进而导致其他严重的环境问题,包括地下水NO-3污染、 地表水富营养化、 土壤酸化、 森林生产力下降、 生物多样性减少以及温室气体N2O的排放量增加等[3, 4, 5].
欧美自20世纪80年代以来开展了许多有关森林生态系统对氮沉降响应的研究[3, 4, 5, 6, 7, 8],15 N稳定同位素技术也被广泛地应用于表征森林生态系统的氮循环特征[9, 10, 11]. 氮同位素分馏,即土壤微生物和植物在利用和转化氮素时倾向于利用更轻的14 N,从而使得流失到生态系统之外的含氮物质(比如经过硝化反应产生的NO-3和反硝化作用产生的N2O)是15 N贫化的,而被固持和离子交换到土壤中的有机氮和NH+4(进而被植物吸收)则是15 N富集的[10, 11, 12],导致土壤和植被的15 N自然丰度值(δ15 N)出现差异[11]. 不少研究发现,氮沉降量增加、 硝化作用增强和氮淋溶增加都会使生态系统的δ15 N增加,进而提出土壤和植物的δ15 N可用来表征生态系统的氮状态[9, 10, 11, 12, 13].
我国近些年Nr排放量激增[14,15],并由此导致氮沉降量的显著增加[16],尤其是我国东部和南部的氮沉降量普遍高于3 g ·(m2 ·a)-1,与欧美地区的最高值相当或者更高[15,17]. 有研究表明氮饱和在我国南方的热带森林[18]和亚热带森林[19]均有发生,但关于这些生态系统在持续高氮输入下的氮循环机制和氮转化过程仍需深入研究. 为此,本研究通过对我国西南地区典型的亚热带森林生态系统土壤和植物15 N自然丰度(δ15 N)的分析,探讨长期的模拟氮沉降(施加氮肥)对该生态系统氮饱和状态的影响.
1 材料与方法 1.1 实验样地
本研究在重庆铁山坪林场一片马尾松林内开展. 马尾松林是我国亚热带地区分布最广且资源最大的森林群落[20]. 该点(106°41.24′E,29°37.42′N)位于重庆市东北,距城中心约20 km,海拔450 m,属典型的亚热带季风气候,年均降水量为1028 mm,年均温度为18.2℃,土壤是山地黄壤. 该地区年均降水的pH值(2001~2004年)为4.0~4.2[19],氮沉降量(2001~2010年)超过4 g ·(m2 ·a)-1,其中NH+4-N的比例在一半以上[21],马尾松林可能已经发生氮饱和[19,22]. 1.2 实验设置
在同一片马尾松林内设立9块14×14 m2的样地,2005年1月至今开展施加不同氮肥(未标记15 N)的野外控制实验[23],用以模拟不同形态氮沉降的影响. 3块平行处理的样地喷洒NH4NO3溶液(记为AN样地),另外3块平行处理的样地喷洒NaNO3溶液(SN样地),剩下3块作为对照样地喷洒与施氮样地相同体积的水(REF样地). 氮肥施加的具体方法是在每月初分别将200 g NH4NO3或400 g NaNO3溶解在10 L去离子水中(相当于每月仅增加0.6 mm的降水量),并以背式喷洒器均匀喷洒在森林土壤上. 施氮样地(AN和SN)的总施加量均为4 g ·(m2 ·a)-1,这与当地氮沉降量相当. 在施加氮肥之前,各样区土壤的pH值无明显差异,在3.6~4.2之间(主要是不同土层的差异),土壤盐基交换容量和总氮量也无明显差异[24]. 在模拟氮沉降实验开展7 a后,即2011年开展不同处理样地15 N自然丰度的研究. 1.3 样品采集与分析
2011年11月初,对各样地土壤有机层和矿质层进行分层采集,每个样地内随机取5个剖面. 有机层包括表层枯落物层(L)和腐殖质层(O),厚度分别在1~2 cm左右; 矿质层用土钻(内径5 cm)进行取样,分0~10 cm、 10~20 cm和20~30 cm共3层. 将同层的5个平行样混合为一个样品进行化学分析.
同时,对各样地的主要植物进行采集,包括马尾松地上部分(针叶和当年生树芯)、 主要地表植物的地上部分和新生细根. 其中,针叶和树芯采集自每个样地内两棵最高的马尾松; 针叶包括当年生和一年生针叶,其中一年生针叶只用于生物量的统计而不进行化学分析. 细根的采集,是在各样地上述两棵马尾松之一的周围3个方向上,每个方向距离树干0.5、 1和1.5 m分别安装内生长土芯,分0~10、 10~20和20~30 cm这3层分别收集称重. 主要地表植物包括芒(Miscanthus sinensis)、 黑足麟毛蕨(Dryopteris fuscipes)、 芒萁(Dicranopteris pedata)和狗脊蕨(Woodwardia japonica),这4种植物占地表草本植物总生物量的95%以上.
土壤样品自然状态下风干,对矿质层进行过筛(<2 mm). 植物样品在70℃烘干至少48 h后称重. 所有土壤和植物样品磨细后(植物磨碎后用四分法,土壤过100目筛以保证混合均匀,减小15 N自然丰度的测量误差),分别测定总氮(TN,凯氏定氮法)和15 N/14 N的原子百分比(同位素质谱仪EA-IRMS,Thermo Scientific DELTA V Advantage,USA). 1.4 数据分析
样品中的15 N自然丰度值用δ15 N表示:
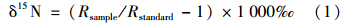
15 N富集因子(εp/s),通常是指反应产物与底物之间δ15 N的差值,这里具体指植物与土壤的δ15 N差值[26]:
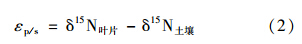
在本研究中,植物选取当年生的针叶,土壤选取0~10 cm层.
计算各种处理的平均δ15 N,以不同处理为变量,进行单因素方差分析(基于SPSS 17.0数据分析软件),P≤0.05时认为具有显著差异. 2 结果与分析 2.1 土壤TN含量与δ15 N
无论对照样地(REF)还是长期施氮样地(AN和SN),土壤各层TN均随土壤深度增加而逐渐降低,由L层的10.6~12.4 g ·kg-1减少到20~30 cm层的0.16~0.19 g ·kg-1(表 1). 但是,土壤各层δ15 N则一般随土壤深度的增加而逐渐升高:REF样地由L层的-6.52‰±0.35‰增加到20~30 cm层的4.51‰±0.00‰; AN和SN样地分别由L层的-4.33‰±0.11‰和-5.83‰±0.05‰增加到10~20 cm层的4.81‰±0.28‰和3.21‰±0.13‰,但20~30 cm层有所下降,分别为3.42‰±0.62‰和1.90‰±0.55‰(图 1).
| 表 1 各样地土壤和植物TN含量与δ15 N 1) Table 1 TN and δ15 N values of soils and plants among different treatments |
比较不同处理之间的差别,施氮样地(AN和SN)除了在20~30 cm层的δ15 N明显低于对照样地之外,其余各层均显著高于对照样地的相应值; 同时,AN样地土壤各层的δ15 N值均显著高于SN样地各层(图 1). REF和SN样地的有机层(L和O层)和矿质上层(0~10 cm层)均为15 N贫化(δ15 N<0),而AN样地仅在有机层为15 N贫化(δ15 N<0).
 | 误差线表示标准差,字母表示不同处理的 差异显著(one-way ANOVA, P≤0.05) 图 1 不同处理样地内土壤各层的δ15 N Fig. 1 δ15 N in soil horizons among different treatments |
各样地当年生针叶的TN含量范围是11.8~12.3 g ·kg-1,地表植物为16.3~17.6 g ·kg-1,新生细根分别为13.7~16.7 g ·kg-1(0~10 cm层)、 14.5~16.9 g ·kg-1(10~20 cm层)和14.5~16.8 g ·kg-1(20~30 cm层). 其中,除0~10 cm层细根AN样地显著高于SN样地之外,其余植物样品在不同的处理间没有显著差异(表 1).
无论对照样地还是施氮样地,δ15 N在各植物样品(针叶、 树芯、 地表植物和新生细根)中均呈负值. 总体来看,δ15 N呈现趋势:新生细根>地表植物>当年生针叶>当年生树芯(AN样地土壤中下层细根最低为例外). 新生细根δ15 N随土壤层深度的增加而升高(SN样地10~20 cm层最高为例外),见图 2.
 | 误差线表示标准差,字母表示不同处理的差异显著(one-way ANOVA, P≤0.05) 图 2 不同处理样地的植物δ15 N Fig. 2 δ15 N in plant species among different treatments |
施氮样地(AN和SN)的δ15 N在各植物样品均显著高于对照样地(REF); 同时,AN样地的δ15 N在土壤中层(10~20 cm)和下层(20~30 cm)的新生细根中显著低于SN样地,而在其他各植物样品中均显著地高于SN样地(图 2).
利用土壤和植物的δ15 N计算各样地的富集因子(εp/s),其中REF样地为-6.2‰±0.5‰,显著低于AN样地(-3.7‰±0.3‰)和SN样地(-5.2‰±0.1‰),而AN样地显著高于SN样地. 3 讨论 3.1 土壤与植物的δ15 N及其影响因素
通常土壤δ15 N随土壤深度的增加而升高 [9, 10, 11, 12, 27, 28, 29, 30, 31, 32],对照样地(REF)结果基本如此,但是施氮样地(AN和SN)下层土壤(20~30 cm)的δ15 N低于中层土壤(10~20 cm),原因是施氮样地已经氮饱和,下层土壤中的NO-3大多没有被植物和微生物利用,因此未经同位素分馏直接被淋溶,导致了该层δ15 N的下降[10,11].
各样地土壤有机层的δ15 N(-6.5‰~-4.0‰)普遍低于欧美的温带森林土壤(-6.0‰~3.3‰)[27, 28, 33],也低于我国东北的温带森林土壤(-4.9‰~-1.0‰)[30]和我国南部的亚热带(-1.7‰~-1.3‰)和热带森林土壤(-1.8‰~-1.0‰)[31,32]. 原因可能有两方面:一是该地区降水和穿透水中的NH+4是15 N贫化的(分别为-5.2‰和-3.3‰)[34]; 二是受林中凋落物δ15 N较低的影响,该地区的凋落物通量较高,为5.96 t ·(hm2 ·a)-1,且其中针叶的比例(48.4%)很高[35],凋落物δ15 N低于土壤表层(由于凋落物在被微生物分解过程中发生15 N分馏)[10,11]. 本研究中针叶δ15 N低于土壤表层,与之相符.
各处理样地的植物δ15 N均为负值(表 1和图 2),这与欧洲已有的一些研究结果相似[11, 12, 13, 28, 29]. 尽管国外大部分热带森林叶片δ15 N为正(3.7‰±3.5‰,n=73),并显著高于许多温带森林(-2.8‰±2.0‰,n=90)[27],但我国华南[31]和台湾[36]的亚热带森林,以及海南岛[32]的热带森林叶片δ15 N多为负值,这与我国热带和亚热带森林土壤有较低的δ15 N相一致[31,32].
对于植物在不同组织间的δ15 N差异(新生细根>地表植物叶片>针叶>树芯),与欧美已有的一些研究结果相符[12, 29, 33, 37],这一差异表明植物在输送和利用氮的过程中发生了15 N的分馏[11,37].
如图 3所示,通过对植物与土壤δ15 N的比较,发现各样地的植物δ15 N均低于土壤矿质层δ15 N,并且矿质上层(0~10 cm)细根δ15 N与该层土壤δ15 N之间有显著的正相关关系[图 3(a)],针叶和地表植物的δ15 N也分别与矿质上层(0~10 cm)土壤δ15 N呈现显著的正相关关系[图 3(b)和3(c)],这与之前的研究结果相似[9, 10, 13, 33, 37],表明植物δ15 N(特别是叶片和细根的δ15 N)能作为一种指标来表征氮来源,以及土壤的氮状态[11, 12, 13, 33, 37, 38].
 | 图 3 细根、 针叶和地表植物δ15 N与土壤矿质上层δ15 N的相关关系 Fig. 3 Correlation of δ15 N in fine roots,current needle,and ground vegetation respectively with δ15 N in upper mineral soil |
长期施氮样地(AN和SN)的土壤和植物δ15 N均显著地高于只接受了自然氮沉降的对照样地(REF),这与很多针对森林的长期施氮研究结论相符[12, 13, 31, 33, 38],即氮输入的增加会提高土壤和植物中的δ15 N. 这主要是两方面的原因:一是无机氮(主要是NH+4)输入的增加,提高了硝化速率(表 2),造成更多15 N贫化的NO-3的淋溶,以及更多15 N富集的NH+4被固持在土壤中,进而被植物利用,因此较高的硝化速率和淋溶导致了土壤和植物中15 N的富集[11, 12, 13, 29, 31]; 二是在高氮输入下,反硝化速率增加[11, 13, 31],反应产生15 N贫化的N2O被排放到系统外,而15 N富集的NO-3被留在土壤内,造成土壤的15 N富化[11, 13, 31],同样导致植物δ15 N的增加.
| 表 2 各处理样地的氮通量与富集因子 1) Table 2 N fluxes and 15 N enrichment factor in different treatments |
同样是接受长期持续的高氮输入,土壤和植物δ15 N在施加NH4NO3(同时增加NH+4和NO-3)和NaNO3(仅增加NO-3)下却存在显著差异. 这说明该生态系统对不同形式的氮输入具有不同的响应. NH+4沉降导致各层土壤δ15 N均显著高于同剂量(按氮计)的 NO-3沉降,原因是土壤中有更多的NH+4硝化(表 2)产生15 N贫化的NO-3被淋溶(AN和SN样地的NO-3淋溶通量相当),而15 N富集的NH+4被留在土壤中. 由于植物在吸收氮的过程中与土壤的氮状态有密切的关系(图 3),因此不同形态氮沉降造成的土壤δ15 N的差异进一步导致了植物δ15 N的显著差异[11, 13, 38]. AN样地植物更多利用的是土壤中15 N富集的NH+4,而SN样地植物利用的更多是15 N相对贫化的NO-3,从而导致了植物δ15 N的差异[37]. 3.3 15 N富集因子及其影响因素
各处理样地的15 N富集因子(εp/s)与其氮输入量、 净硝化速率和氮淋溶量(主要是NO-3淋溶)均呈显著的正相关关系(表 2),而这些通量又与生态系统是否氮饱和有着密切关系(例如,大部分氮输入被淋溶,表明这些样地已经氮饱和)[2]. 不少研究尝试用15 N富集因子来表征森林系统的氮状态,因为不论是相对封闭的系统[9, 11, 12, 28, 38],还是新近研究的氮淋溶很高的亚热带、 热带地区的开放森林生态系统[31,32],都证实了富集因子与氮通量之间的显著相关性. Emmett 等[12] 通过总结欧洲4个地区温带森林的15 N自然丰度值,发现了富集因子(εp/s)与氮沉降量之间存在显著的正相关关系(R2=0.98,P<0.01; 图 4),但是该研究只是针对欧洲的温带森林系统,且氮沉降量在1~6 g ·(m2 ·a)-1之间.
本研究得到REF、 AN和SN样地的εp/s分别为-6.2‰±0.5‰、 -3.7‰±0.3‰和-5.2‰±0.1‰(表 2). 对中外其他地区的研究结果进行分析,这些地区包括美国北部的温带森林[28,38]和南部的热带森林[9],中国东北大兴安岭地区的寒温带森林[30],以及中国南部的亚热带森林[31,39]和热带森林[32],发现所有马尾松林(包括本研究TSP、 广东鼎湖山DHS和贵州鹿冲关LCG)的富集因子和氮沉降量有着显著的线性相关关系(R2=0.74,P≤0.05),不同于欧洲森林的结果[12],而我国和美国的另外一些森林类型则可能满足其他的关系(图 4). 以上不同研究结果的对比初步表明,富集因子可用来表征生态系统的氮状态,不过还要同时考虑森林类型等其他因素,这需更多观测结果的支持.
 | 图 4 15 N富集因子与氮沉降量之间的关系 Fig. 4 Relationship between 15 N enrichment factor and N deposition |
(1)通过对我国西南地区典型马尾松林的土壤和植物15 N自然丰度进行分析和比较,表明长期持续的高氮输入会增加土壤和植物的15 N自然丰度值. 氮沉降的增加同样会提高森林生态系统的富集因子. 15 N富集因子有可能作为表征生态系统的氮状态的指标,不过还需更多的观测结果的支持.
(2)该森林系统对于不同形式的氮沉降(NH+4或NO-3)有着不同的响应,植物和土壤的δ15 N在同剂量(按氮计)的NH+4 沉降下显著高于NO-3沉降情况. 考虑到NH+4沉降的效应更明显,而我国目前的氮沉降控制主要针对NOx排放削减(降低NO-3沉降),因此建议同时重视NH3排放的削减,从而降低NH+4沉降.
| [1] | Galloway J N, Townsend A R, Erisman J W, et al. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions [J]. Science, 2008, 320 (5878): 889-892. |
| [2] | Aber J D, McDowell W H, Nadelhoffer K J, et al. Nitrogen saturation in temperate forest ecosystems: hypotheses revisited [J]. BioScience, 1998, 48 (11): 921-934. |
| [3] | Aber J D, Magill A H. Chronic nitrogen additions at the Harvard Forest (USA): the first 15 years of a nitrogen saturation experiment [J]. Forest Ecology and Management, 2004, 196 (1): 1-5. |
| [4] | Kreutzer K, Butterbach-Bahl K, Rennenberg H, et al. The complete nitrogen cycle of an N-saturated spruce forest ecosystem [J]. Plant Biology, 2009, 11 (5): 643-649. |
| [5] | 袁颖红, 樊后保, 李燕燕, 等. 模拟氮沉降对土壤酸化和土壤盐基离子含量的影响 [J]. 应用与环境生物学报, 2011, 17 (4): 461-466. |
| [6] | Gundersen P, Emmett B A, Kjφnaas O J, et al. Impact of nitrogen deposition on nitrogen cycling in forests: a synthesis of NITREX data [J]. Forest Ecology and Management, 1998, 101 (1-3): 37-55. |
| [7] | Emmett B A, Boxman D, Bredemeier M, et al. Predicting the effects of atmospheric nitrogen deposition in conifer stands: evidence from the NITREX ecosystem-scale experiments [J]. Ecosystems, 1998, 1 (4): 352-360. |
| [8] | Magill A H, Aber J D, Berntson G M, et al. Long-term nitrogen additions and nitrogen saturation in two temperate forests [J]. Ecosystems, 2000, 3 (3): 238-253. |
| [9] | Garten C T. Variations in foliar 15N abundance and the availability of soil nitrogen on Walker Branch Watershed [J]. Ecology, 1993, 74 (7): 2098-2113. |
| [10] | Nadelhoffer K J, Fry B. Nitrogen isotope studies in forest ecosystems[A]. In: Michener K L A R H, (ed). Stable Isotopes in Ecology and Environmental Science [M]. Oxford: Blackwell Scientific Publishers, 1994. 22-44. |
| [11] | Högberg P. Tansley review No.95: 15N natural abundance in soil-plant systems [J]. New Phytologist, 1997, 137 (2): 179-203. |
| [12] | Emmett B A, Kjφnaas O J, Gundersen P, et al. Natural abundance of 15N in forests across a nitrogen deposition gradient [J]. Forest Ecology and Management, 1998, 101 (1-3): 9-18. |
| [13] | Pardo L H, Templer P H, Goodale C L, et al. Regional assessment of N saturation using foliar and root δ15N [J]. Biogeochemistry, 2006, 80 (2): 143-171. |
| [14] | Zhao Y, Duan L, Xing J, et al. Soil acidification in China: is controlling SO2 emissions enough? [J]. Environmental Science and Technology, 2009, 43 (21): 8021-8026. |
| [15] | Liu X J, Duan L, Mo J M, et al. Nitrogen deposition and its ecological impact in China: an overview[J]. Environmental Pollution, 2011, 159 (10): 2251-2264. |
| [16] | Liu X J, Zhang Y, Han W X, et al. Enhanced nitrogen deposition over China [J]. Nature, 2013, 494 (7438): 459-462. |
| [17] | Lü C Q, Tian H Q. Spatial and temporal patterns of nitrogen deposition in China: synthesis of observational data [J]. Journal of Geophysical Research, 2007, 112 (D22): D22S05. |
| [18] | Lu X K, Mao Q G, Gilliam F S, et al. Nitrogen deposition contributes to soil acidification in tropical ecosystems [J]. Global Change Biology, 2014, 20 (12): 3790-3801. |
| [19] | Larssen T, Duan L, Mulder J. Deposition and leaching of sulfur, nitrogen and calcium in four forested catchments in China: implications for acidification [J]. Environmental Science & Technology, 2011, 45 (4): 1192-1198. |
| [20] | 吴征镒. 中国植被 [M]. 北京: 科学出版社, 1980. 235. |
| [21] | 余德祥, 马萧萧, 谭炳全, 等. 重庆市铁山坪2001-2010年酸沉降变化 [J]. 环境科学, 2014, 35 (1): 60-65. |
| [22] | Chen X Y, Mulder J. Indicators for nitrogen status and leaching in subtropical forest ecosystems, South China [J]. Biogeochemistry, 2007, 82 (2): 165-180. |
| [23] | 段雷, 周益, 杨永森, 等. 酸化及化学修复剂对森林土壤有机物淋溶的影响 [J]. 环境科学, 2008, 29 (2): 440-445. |
| [24] | 杨永森, 段雷, 靳腾, 等. 石灰石和菱镁矿对酸化森林土壤修复作用的研究 [J]. 环境科学, 2006, 27 (9): 1878-1883. |
| [25] | Mariotti A. Atmospheric nitrogen is a reliable standard for natural δ15N abundance measurements [J]. Nature, 1983, 303 (5919): 685-687. |
| [26] | Mariotti A, Germon J C, Hubert P, et al. Experimental determination of nitrogen kinetic isotope fractionation: some principles; illustration for the denitrification and nitrification processes [J]. Plant and Soil, 1981, 62 (3): 413-430. |
| [27] | Martinelli L A, Piccolo M C, Townsend A R, et al. Nitrogen stable isotopic composition of leaves and soil: tropical versus temperate forests [J]. Biogeochemistry, 1999, 46 (1-3): 45-65. |
| [28] | Templer P H, Arthur M A, Lovett G M, et al. Plant and soil natural abundance δ15N: indicators of relative rates of nitrogen cycling in temperate forest ecosystems [J]. Oecologia, 2007, 153 (2): 399-406. |
| [29] | Koopmans C J, Van Dam D, Tietema A, et al. Natural 15N abundance in two nitrogen saturated forest ecosystems [J]. Oecologia, 1997, 111 (4): 470-480. |
| [30] | Cheng S L, Fang H J, Yu G R, et al. Foliar and soil 15N natural abundances provide field evidence on nitrogen dynamics in temperate and boreal forest ecosystems [J]. Plant and Soil, 2010, 337 (1-2): 285-297. |
| [31] | Fang H J, Yu G R, Cheng S L, et al. Nitrogen-15 signals of leaf-litter-soil continuum as a possible indicator of ecosystem nitrogen saturation by forest succession and N loads [J]. Biogeochemistry, 2011, 102 (1-3): 251-263. |
| [32] | Wang A, Fang Y T, Chen D X, et al. Variations in nitrogen-15 natural abundance of plant and soil systems in four remote tropical rainforests, southern China [J]. Oecologia, 2014, 174 (2): 567-580. |
| [33] | Högberg P, Högbom L, Schinkel H, et al. 15N abundance of surface soils, roots and mycorrhizas in profiles of European forest soils [J]. Oecologia, 1996, 108 (2): 207-214. |
| [34] | 余龙飞, 李炳文, 张逸, 等. 我国一些站点降雨和森林穿冠水中铵态氮的同位素丰度 [J]. 环境化学, 2012, 32 (9): 1599-1603. |
| [35] | 王轶浩, 王彦辉, 于澎涛, 等. 重庆酸雨区马尾松林凋落物特征及对干旱胁迫的响应 [J]. 生态学报, 2013, 33 (6): 1842-1851. |
| [36] | Liu C P, Yeh H W, Sheu B H. N isotopes and N cycle in a 35-year-old plantation of the Guandaushi subtropical forest ecosystem, central Taiwan [J]. Forest Ecology and Management, 2006, 235 (1-3): 84-87. |
| [37] | Pardo L H, Semaoune P, Schaberg P G, et al. Patterns in δ15N in roots, stems, and leaves of sugar maple and American beech seedlings, saplings, and mature trees [J]. Biogeochemistry, 2013, 112 (1-3): 275-291. |
| [38] | Pardo L H, Hemond H F, Montoya J P, et al. Natural abundance 15N in soil and litter across a nitrate output gradient in New Hampshire [J]. Forest Ecology and Management, 2007, 251 (3): 217-230. |
| [39] | 蒋春来, 张晓山, 肖劲松, 等. 我国西南地区氮沉降量不同的森林小流域中土壤自然15N丰度的分布特征 [J]. 岩石学报, 2009, 25 (5): 1291-1296. |
| [40] | 马萧萧. 化学修复与模拟氮沉降对酸化森林土壤的影响 [D]. 北京: 清华大学, 2011. |