 2015, Vol. 36
2015, Vol. 36

2. 中国科学院大学, 北京 100049;
3. 加拿大农业与农业食品部温室与加工作物研究中心, 安大略 N0R 1G0
 , LIANG Ai-zhen1, YANG Xue-ming3, ZHANG Xiao-ping1
, LIANG Ai-zhen1, YANG Xue-ming3, ZHANG Xiao-ping1 , JIA Shu-xia1, CHEN Xue-wen1, ZHANG Shi-xiu1, SUN Bing-jie1,2, CHEN Sheng-long1,2
, JIA Shu-xia1, CHEN Xue-wen1, ZHANG Shi-xiu1, SUN Bing-jie1,2, CHEN Sheng-long1,2
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Greenhouse and Processing Crops Research Centre, Agriculture and Agri-Food Canada, Ontario N0R 1G0, Canada
我国每年的秸秆资源总量为7.62×108~8.41×108 t[1, 2],其中很大一部分被直接焚烧,这对大气环境产生了严重的污染[3]. 实施秸秆还田不仅可以减轻其焚烧对大气环境的污染,也是增加土壤有机碳含量,提升土壤地力的重要措施[4]. 秸秆还田后作物残体的分解是农田生态系统碳循环及养分周转平衡的一个至关重要环节[5, 6]. 探明秸秆在土壤中的分解转化特征及其影响因素不仅对全球变暖背景下的秸秆还田固碳研究具有重要意义[7],也可以作为秸秆还田后土壤养分管理的重要参考依据[8].
植物残体的化学性质对土壤中秸秆分解、 CO2排放和微生物演替有重要的影响[9, 10]. 张红等[11]对不同植物秸秆的腐解特征和微生物功能多样性进行了研究,认为C/N低、 体积小的秸秆在腐解前期分解速度快,而C/N高、 体积小的秸秆会在腐解后期加快分解. 木质素是秸秆中一类很难分解的高分子化合物[12],它的含量也会对秸秆的分解产生重要的影响[13]. 碳氮比和木质素含量低的秸秆在土壤中会有较快地分解和氮矿化速率并促进微生物量的增加; 而碳氮比和木质素含量较高秸秆的分解速率相对较慢,矿化释放的氮素相对较少[14].
除了秸秆的化学性质外,土壤质地也是秸秆分解转化的重要影响因素. 由于黏粒与有机质的黏结能力比砂壤土强,黏壤土可以提供比砂壤土更好的物理保护作用[15]. 但是Fang等[16]通过培养实验发现,混合有玉米秸秆的粉质黏土比混合玉米秸秆的砂壤土有更高的CO2排放量. 这可能是因为黏壤土具有比砂壤土更高的养分和水分保持能力,添加秸秆后,土壤中较高的养分含量提高了微生物的生物量和代谢能力[17].
玉米是东北黑土区广泛种植的粮食作物,其根、 茎、 叶等不同部位秸秆的C/N、 木质素含量等化学性质存在显著差异[18]. 本研究分析了添加不同植株部位玉米秸秆对黏壤土CO2排放和土壤微生物量的影响及其与秸秆碳、 氮含量之间关系[19],但添加不同植株部位玉米秸秆对砂壤土的CO2排放和土壤微生物量影响如何,秸秆的木质素含量与土壤的CO2排放、 微生物量之间存在何种关系,两种质地土壤的CO2排放征、 微生物量存在何种差异还有待进一步的研究. 因此,本文选择了位于东北黑土区的两种不同质地土壤,通过室内恒温培养实验重点探讨:①添加不同植株部位玉米秸秆后的砂壤土CO2排放特征和微生物量大小是否与黏壤土存在差异,②添加不同木质素和全氮含量的玉米秸秆进入土壤后,其CO2排放、 微生物量大小是否存在差异; 哪个指标最能说明土壤CO2排放的差异. 通过以上研究,旨在明晰秸秆的化学性质和土壤质地对土壤CO2排放和微生物量的影响,为东北黑土区玉米秸秆还田的实施与农田管理提供参考依据.
1 材料与方法 1.1 土壤与秸秆培养实验的黏壤土采自于吉林省德惠市米沙子乡(44°12′ N,125°33′ E)中国科学院东北地理与农业生态研究所黑土农业生态试验示范基地的农田黑土表层(0~10 cm). 砂壤土采自于吉林省农安县哈拉海镇(44°42′ N,125°06′ E)农田黑土表层(0~10 cm). 地表作物均为玉米. 采集的土壤过2 mm筛后放在4℃保鲜冰箱中保存,用于培养实验和基本理化性质测定. 培养实验前土壤的基本性质见表 1.
| 表 1 培养实验土壤的基本理化性质 Table 1 Selected physical and chemical properties of soils used for incubation trial |
玉米(Zea mays)秸秆于2013年10月采自中国科学院东北地理与农业生态研究所长春综合农业试验站. 玉米品种为先玉335,采样时玉米和大豆均已经完全成熟. 将采集的玉米秸秆分成根、 茎下部(3节以下)、 茎中间部分、 茎顶部(7节以上)、 叶五部分. 由于玉米茎的性质存在垂直分布逐渐过渡现象,茎中部性质介于茎下部和茎顶部之间,而茎下部、 茎顶部秸秆之间有显著的差异,故未将茎中间部分进行培养实验. 将上述秸秆装入牛皮纸袋中在60℃条件下烘干至恒重,然后用植物粉碎机以29 000 r ·min-1的速率粉碎5 min,使它们的形状大小基本一致后用于培养实验. 使用FlashEATM 1112元素分析仪测定总碳和总氮含量,根据Vonset方法[20]使用VELP纤维素分析仪测定木质素含量(表 2).
| 表 2 玉米不同部位全碳、全氮、木质素含量及比值 Table 2 Total carbon, total nitrogen,lignin contents, C/N and lignin/N in different parts of corn residues |
培养实验采用两因素设计,每个处理设置4个重复. 实验因素为添加秸秆类型:玉米根、 茎下部、 茎顶部和叶,土壤质地:黏壤土、 砂壤土. 取烘干粉碎后的玉米根、 茎下部、 茎顶部、 叶1.5 g分别与相当于烘干土150 g的鲜土进行充分混合,然后置于500 mL广口瓶中进行培养,同时设置不加秸秆的空白对照. 调节土壤含水量至40%的孔隙含水率,然后用保鲜膜将广口瓶瓶口封住,并用注射器针头扎若干细孔保证土壤含水量和培养瓶中的氧气浓度. 将装有土壤的广口瓶置于25℃的恒温培养箱中进行培养,每两天进行称重补水. 广口瓶放入培养箱稳定2 d后,在培养的第3、 4、 5、 6、 7、 8、 9、 11、 13、 15、 17、 21、 25、 29、 33、 37、 41、 45、 49、 53、 57、 61 d对土壤的CO2排放速率进行测定,然后破坏性取样测定土壤的微生物量碳、 氮和总溶解性氮浓度.
1.3 测定项目与方法 1.3.1 土壤CO2排放速率的测定与计算利用LI-820 CO2红外气体分析仪,根据封闭动态箱原理对土壤CO2排放速率进行测定计算[21]. 将广口瓶、 LI-820构建成封闭动态体系(图 1),并对封闭动态体系内的CO2浓度进行120 s实时动态测定. 将测定的CO2浓度与测定时间进行线性回归,计算出封闭动态体系内CO2浓度的增加速率a. CO2排放速率根据公式(1)进行计算:

式中,F为培养物质的CO2产生速率,单位是mol ·(g ·s)-1; Vm是CO2摩尔体积,M是培养物质的质量; V为封闭动态系统内的体积.
 |
图 1 测定CO2排放速率时的封闭动态体系示意
Fig. 1 Diagrammatic sketch of enclosed dynamic system for CO2 emission rate measuring
|
采用氯仿熏蒸-K2SO4浸提法,用岛津TOC-VCPH分析仪测定提取液中的碳、 氮含量[22]. 根据熏蒸和未熏蒸处理的提取液中有机碳之差除以转换系数0.38为MBC[23],根据熏蒸和未熏蒸处理的提取液中全氮之差除以转换系数0.45为MBN[24].
1.3.3 土壤总溶解性氮测定土壤中的总溶解性氮(TDN)使用0.5mol ·L-1的K2SO4(土壤、 溶液比为1 ∶4)浸提,用岛津TOC-VCPH/CPN分析仪测定浸提液中的总氮含量.
1.4 数据处理与统计分析 1.4.1 CO2累积排放量计算参考文献[25]的方法,根据培养期间测定的CO2排放速率进行分段积分计算培养期间的CO2累积排放量. 具体计算方法见公式(2):
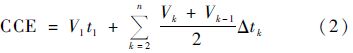
式中,CCE表示培养期间的CO2累积排放量,V1是第一次测定时的CO2排放速率,t1指从开始培养到第一次测定的间隔时间,Vk为第k次测定时的CO2排放速率,Δtk表示第k-1次测量到第k次测量的间隔时间.
1.4.2 激发效应计算秸秆添加后的激发效应(priming effect,PE)根据公式(3)计算[26]:
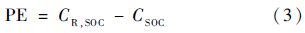
式中,CR,SOC指添加秸秆的土壤CO2累积排放量,CSOC是未添加秸秆的土壤CO2排放量.
1.4.3 统计分析使用Excel 2013进行平均值及标准差计算; 使用SAS 9.3 PROC组块的GLM过程进行双因素方差分析和Tukey多重比较进行显著性差异检验(P=0.05); 用REG过程进行线性回归,分析秸秆化学属性与与土壤激发效应之间的相关关系; 使用SigmaPlot 10.0进行绘图.
2 结果与分析 2.1 土壤的CO2排放由图 2可知,添加秸秆显著提高了两种质地土壤的CO2排放速率,其CO2排放速率[26~160 pmol ·(g ·s)-1]是未添加秸秆土壤CO2排放速率[12~18 pmol ·(g ·s)-1]的2~13倍. 在61 d的培养实验中,未添加秸秆的土壤CO2排放速率基本稳定,而添加秸秆的土壤CO2排放速率随时间均整体呈现出“快速下降-稳定-出现一较高值(在30 d前后)-持续下降”的过程. 除玉米叶外,秸秆添加处理的黏壤土CO2排放速率在培养的绝大多数时间段均显著小于对应的砂壤土CO2排放速率.
 |
图 2 玉米不同部位秸秆在培养期间的土壤CO2排放速率(均值±标准差)
Fig. 2 CO2 emission rate of clay loam and sandy loam soils amended with different crop residue samples(Mean value±standard deviation)
|
双因素方差分析显示,秸秆类型、 土壤质地及其它们的交互作用对土壤CO2累积排放量有显著影响(表 3). 由图 3可知,添加秸秆显著提高了黏壤土和砂壤土的CO2累积排放量,其数值分别是相应对照处理的4.5~5.64倍和4.8~6.6倍. 两种土壤中根和茎顶部秸秆的CO2累积排放量均显著小于对应茎下部和叶的CO2累积排放量. 对照处理、 根、 茎下部、 茎顶部在黏壤土中的CO2累积排放量均显著大于对应的砂壤土18.38%~25.64%,但是玉米叶在两种质地土壤中的CO2累积排放量没有显著性差异.
 |
不同大写字母表示黏壤土下不同秸秆添加处理的差异显著(P<0.05),不同小写字母表示砂壤土下不同秸秆添加处理的差异显著(P<0.05),*表示该秸秆添加处理下的两种质地土壤差异显著(P<0.05),**表示该秸秆添加处理下的两种质地土壤差异极显著(P<0.01),下同图 3 不同类型秸秆培养期间的CO2累积排放量(均值±标准差)
Fig. 3 Cumulative CO2 efflux from clay loam and sandy loam soils amended with different crop residues (Mean value±standard deviation)
|
| 表 3 秸秆类型、土壤质地及其交互作用对土壤 CO2累积排放量、MBC、 MBN、 MBC/MBN、 TDN、 PE值的影响1) Table 3 Effects of residue type, soil texture and their interaction on cumulative CO2 efflux, MBC, MBN, MBC/MBN, TDN and PE of soil amended with different crop residues |
由图 4可知,添加不同秸秆进入两种土壤后产生的激发效应有明显差别,其PE值介于216.53~335.17μmol ·g-1. 双因素方差分析显示,添加秸秆产生的激发效应值受到秸秆类型、 土壤质地以及土壤质地与秸秆类型交互作用的显著影响,秸秆类型对PE值的影响最显著,土壤质地以及土壤质地与秸秆类型交互作用对PE值影响的显著性依次减小(表 3). 两种土壤中根和茎顶部秸秆PE值平均值均小于茎下部和叶; 且两种土壤中茎顶部秸秆的PE值均显著小于茎下部和叶,砂壤土中根的PE值也显著小于叶. 除砂壤土中叶的PE值略大于对应的砂壤土外,其他各类型秸秆的PE值均表现为黏壤土显著大于砂壤土.
 |
图 4 不同类型秸秆在培养期间的激发效应值(均值±标准差)
Fig. 4 Priming effect of soil amended with different crop residues(Mean value±standard deviation)
|
由图 5可知,PE值与秸秆的木质素/N之间的线性关系明显好于PE值与秸秆木质素含量、 C/N、 含氮量之间的线性关系. 两种土壤中PE值与木质素/N之间线性回归方程的截距比较接近,但是砂壤土的斜率(-7.320 79)比黏壤土的斜率(-4.660 95)小57.07%. 当秸秆木质素/N较低时,两种土壤的PE值比较接近; 而随着木质素/N增大,两种土壤的PE值之差也逐渐增大.
 |
图 5 秸秆化学属性与砂壤土、黏壤土PE值的相关关系
Fig. 5 Correlation of priming effect with residue chemistry of clay loam and sandy loam soils amended with different crop residues
|
双因素方差分析表明,土壤MBC、 TDN受到秸秆类型、 土壤质地及其交互作用的显著影响,而MBN、 MBC/MBN只受到秸秆类型的显著影响(表 4). 添加秸秆的土壤MBC、 MBN显著大于未添加秸秆的对照处理,而MBC/MBN、 TDN则显著小于未添加秸秆的对照处理. 在两种土壤中,茎下部和叶的MBC和MBN均显著大于根和茎顶部的MBC和MBN; 而TDN则表现为根和叶的含量显著大于茎下部和茎顶部,茎下部的含量显著大于茎顶部. 虽然同种秸秆添加下的两种质地土壤MBC、 MBN没有显著差异,但添加秸秆后砂壤土中微生物量增加的幅度大于黏壤土的微生物量增加幅度. 黏壤土中添加秸秆的土壤MBC、 MBN分别比对照处理大105%~146%、 185%~299%; 砂壤土中添加秸秆的土壤MBC、 MBN分别比对照处理大150%~295%、 423%~552%. 各处理下的砂壤土中TDN含量都显著小于对应处理黏壤土的矿质态氮含量11.77%~31.33%.
| 表 4 不同部位玉米秸秆在土壤中培养后的微生物量碳、微生物量氮、 矿质态氮含量(均值±标准差)1) Table 4 Concentration of soil microbial biomass and mineral N in soil amended with different parts of corn residues |
添加秸秆的土壤CO2排放速率随时间表现出 “快速下降-稳定-出现较高值-持续下降”的过程,这与土壤中易分解有机物质耗竭和微生物类群的增长有关. 秸秆与土壤混合后CO2排放速率由较高的排放速率迅速下降,是因为秸秆进入土壤后易分解的短链有机物质(单糖、 淀粉和简单蛋白质等)被耗竭所致; 在培养30 d左右出现的“较高值”,与能分解长链或环状有机物微生物类群的增长有关; 之后可分解的有机物质继续减少,这导致了CO2排放速率的进一步衰减[27,28].
文中玉米秸秆各部位木质素含量表现为:根>茎>叶,与Abiven等[18]观察到的结果一致. 茎下部和茎顶部秸秆的木质素含量基本一致,而茎顶部秸秆的含氮量只有茎下部秸秆的一半. 这说明玉米秸秆的全氮含量在玉米茎中存在垂直分布的差异,而木质素含量则没有. 由于具有最高的木质素含量,根的CO2累积排放量显著小于茎下部和叶的CO2累积排放量. 但是,由于茎顶部秸秆的含氮量很低,尽管它的木质素含量平均值只有根的54%,CO2累积排放量却并没有显著小于根. 这说明根的CO2累积排放量小是由于高的木质素含量,而茎顶部的CO2累积排放量小是受限于较低氮的含量. 显然,玉米秸秆的分解与CO2排放受到自身木质素及氮含量的共同影响,根部和茎顶部秸秆的共同特点是有较高的木质素/N. 所以,木质素/N比木质素含量、 C/N等更好地说明秸秆分解和CO2排放的差异. Johnson等[29]和Yanni等[30]也认为木质素全氮比高的秸秆在土壤中的分解速率会相对缓慢,这与本研究结果一致.
除玉米叶外,添加其他秸秆的黏壤土CO2排放速率和CO2累积排放量均显著大于对应的砂壤土,这意味着黏粒保护作用不是两种土壤中秸秆分解差异的主要原因. 添加秸秆后,土壤中的有效氮等营养物质成为分解植物残体微生物活性的限制因素; 而黏壤土的营养物质丰富,有利于土壤中微生物数量和活性的提高,从而促进了秸秆的分解和CO2的排放[17]. 而叶在两种土壤中的CO2累积排放量没有差异,是因为叶的C/N、 木质素/N比较小(表 2). 一方面,是因为叶的氮素含量高、 木质素含量低,微生物对它的分解作用较少受到土壤中矿质氮等营养物质含量的限制[14]; 另一方面,这类秸秆与黏粒的结合能力比含氮少、 木质素多的秸秆强,可以减少一部分微生物的分解作用[31]. 由此可以认为,秸秆木质素含量越高、 总氮越低受到土壤质地的影响就越大; 而类似玉米叶等木质素含量较低、 总氮含量较高的秸秆在两种土壤中的分解速率差异就很小.
3.2 秸秆化学属性与两种质地土壤激发效应的关系由于两种质地土壤的CO2排放本底值存在显著差异,计算添加秸秆的PE值可以更好确定不同处理下CO2排放的差异的来源[32]. 双因素方差分析的结果显示,秸秆类型对PE值有极显著影响,而土壤质地以及土壤质地与秸秆类型的交互作用对PE值影响的显著性依次降低,说明秸秆类型比土壤质地对CO2排放差异的贡献更大. 不同秸秆添加下PE值差异与土壤CO2累积排放量的差异基本一致,PE值与秸秆木质素/N的呈负的线性关系,且明显好于其他指标与PE值的线性关系. 这进一步证明了前文关于秸秆类型和土壤质地对秸秆分解、 CO2排放影响的论断,木质素/N确实比木质素含量、 含氮量、 C/N等更好地说明秸秆分解和CO2排放的差异.
两种质地土壤的PE值与木质素/N线性回归方程的截距比较接近,但砂壤土的斜率却明显小于黏壤土的斜率. 两种质地土壤的激发效应对不同木质素/N的响应不一致,随着秸秆木质素/N的增大,砂壤土PE值降低的速率更快,与相应黏壤土的差值更大. 正如前文论断,由于氮素限制作用和黏粒结合能力的差异[14,31],随着秸秆木质素/N的增加,质地对秸秆分解、 CO2排放的影响逐渐增大. 这意味着,在黑土区实施秸秆还田时应更加注意木质素/N较高的秸秆在不同质地土壤中的分解、 CO2排放的差异.
3.3 秸秆类型对两种质地土壤微生物量和TDN含量的影响添加秸秆的土壤微生物量显著大于未添加秸秆的对照处理,而TDN含量显著小于未添加秸秆的对照处理. 这是因为添加秸秆提供了物质和能源促进了微生物的生长繁殖,为了满足生长繁殖和分解秸秆的需要,微生物必须从土壤中吸收含氮底物,从而降低了土壤中的TDN含量[33]. 添加玉米秸秆增加了MBC、 MBN含量,但MBC/MBN降低,说明更多的含氮底物被微生物固持,微生物群落氮固持的速率高于碳固持. 同时,MBC/MBN还可以大致反映出微生物的种群结构信息[34],添加秸秆的土壤MBC/MBN显著减小,是因为添加的秸秆在分解早期具有较多可利用碳源(糖类、 半纤维素、 纤维素等)和营养物质更有利于细菌、 放线菌等具有较低C/N微生物的生长繁殖,而这类微生物往往以r 策略生存(r-strategy)[27, 28].
植物残体的化学性质在影响植物残体分解速率的同时也会对土壤的TDN含量、 微生物量及微生物的群落结构产生一定影响. 有研究认为木质素含量和C/N较低的残体在土壤中比较容易分解,有较高的CO2排放、 矿质氮含量以及微生物量; 而较高C/N和木质素含量的植物残体更难分解,其CO2排放、 矿质氮含量、 微生物量相对较小,同时还会促使真菌等适应营养贫乏环境的微生物丰度增加[14,35]. 本研究结果也部分证实了上述观点. 在本研究中,玉米根和茎顶部秸秆的木质素/N显著大于茎下部和叶,它们的MBC、 MBN也和CO2累积排放量一样显著小于茎下部和叶; 黏壤土中玉米根和茎顶部秸秆的MBC/MBN显著大于茎下部和叶,说明根和茎顶部的真菌比例显著大于茎下部和叶[34].
然而,根的TDN含量和砂壤土MBC/MBN的数据与已有的研究结果不一致. 根的TDN含量没有显著小于其它类型秸秆可能是因为,土壤中TDN含量受到土壤中微生物同化固定和秸秆矿化作用的共同发影响[36, 37]. 一方面,根的木质素含量比较高,较难被微生物分解利用,土壤中微生物增加量相对较低,微生物从土壤中吸收固定的含氮底物数量比其它类型秸秆少; 另一方面,由于根的全氮含量较高,虽然根的腐解速率比较慢,依然可以通过矿化作用向土壤中补充较多的矿质氮. 而砂壤土中MBC/MBN反映的微生物群落结构信息与黏壤土有较大差异,这说明秸秆性质对土壤微生物群落结构影响因土壤种类而异. 土壤微生物遗传背景以及土壤机械组成的差异可能是不同土壤中微生物群落结构对秸秆添加响应不同的重要原因[36,38]. 必须指出的是,MBC/MBN只能大致反映土壤微生物群落结构的变化,需要PLFA、 高通量测序等方法才能更好解读出土壤中微生物群落结构信息.
砂壤土各处理的土壤TDN含量均显著小于对应的黏壤土,这与砂壤土的CO2累积排放量小于对应处理黏壤土的结果一致. 而尽管砂壤土MBC平均含量比黏壤土小,但相同秸秆添加处理下的砂壤土MBC、 MBN没有显著性差异. 这意味着土壤有效氮含量导致的微生物代谢能力差异可能是砂壤土中秸秆分解速率小于对应黏壤土的重要原因.
3.4 对秸秆还田实施与管理的探讨秸秆还田被很多研究人员认为是提高土壤地力、 增加土壤碳库储量的重要措施[39]. 本研究通过两种质地土壤的秸秆添加实验发现,砂壤土中秸秆的分解矿化速率、 CO2排放量显著小于黏壤土; 砂壤土中微生物量增加的幅度比黏壤土中微生物量增加的幅度大. 这意味着,与黏壤土相比,在砂壤土中实施秸秆还田可以取得更好的土壤碳固存和微生物量保持效果. 而土壤微生物量既是土壤有机质和养分转化的动力,又是土壤中植物有效养分的储备库,对土壤养分的循环和有效性有着至关重要的作用[40]. 与此同时,因为黏粒含量较少,砂壤土中的TDN更容易被降雨、 径流淋失,这不仅降低了农田土壤的氮肥利用率,也会对地下水和地表水产生污染[41]. 而添加秸秆后砂壤土中的TDN可以被微生物吸收同化固定,有利于土壤氮素的保存和周转. 陈维梁等也认为秸秆还田能够有效缓解施肥带来的氮淋溶效应,降低地下水氮素污染风险,从而达到改善土壤肥力从而增加农作物产量的目的[42]. 当然,在田间实践时还需要适当补充速效氮肥,来防止秸秆分解过程中微生物与作物争夺土壤有效态氮引起的土壤有效养分短期供给不足的问题. 由此可以认为,就土壤碳固存、 微生物量保持和氮保存而言,在砂壤土中实施秸秆还田的效果可能比黏壤土更好.
4 结论(1)不同植株部位秸秆的C/N、 木质素含量等有较大差异,其分解的难易程度也不同. 玉米根和茎顶部秸秆具有较高的C/N或木质素含量相对较难分解,其CO2排放、 激发效应和微生物量均显著小于茎下部和叶. 秸秆的木质素和氮含量均会对它的分解和CO2排放产生影响,木质素/N比木质素含量、 C/N等更好地说明秸秆分解和CO2排放的差异.
(2) 除玉米叶外,添加其他秸秆的黏壤土在培养期间的CO2排放速率、 CO2累积排放量和激发效应均显著大于对应的砂壤土. 黏粒保护的作用不是两种土壤中秸秆分解差异的主要原因,土壤中的有效氮等营养物质可能是植物残体分解的限制因素. 砂壤土中微生物量增加的幅度比黏壤土中微生物量增加的幅度大. 与黏壤土相比,在砂壤土中实施秸秆还田更有利于土壤碳固存、 微生物量保持和氮保存.
| [1] | 高利伟, 马林, 张卫峰, 等. 中国作物秸秆养分资源数量估算及其利用状况[J]. 农业工程学报, 2009, 25 (7): 173-179. |
| [2] | 谢光辉, 王晓玉, 任兰天. 中国作物秸秆资源评估研究现状[J]. 生物工程学报, 2010, 26 (7): 855-863. |
| [3] | 唐喜斌, 黄成, 楼晟荣, 等. 长三角地区秸秆燃烧排放因子与颗粒物成分谱研究[J]. 环境科学, 2014, 35 (5): 1623-1632. |
| [4] | 王如芳, 张吉旺, 董树亭, 等. 我国玉米主产区秸秆资源利用现状及其效果[J]. 应用生态学报, 2011, 22 (6): 1504-1510. |
| [5] | Parton W, Silver W, et al. Global-scale similarities in nitrogen release patterns during long term decomposition [J]. Science, 2007, 315 (5184): 361-364. |
| [6] | Randlett D L, Zak D R, Pregitzer K S, et al. Elevated atmospheric carbon dioxide and leaf litter chemistry: Influences on microbial respiration and net nitrogen mineralization [J]. Soil Science Society of America Journal, 1996, 60 (5): 1571-1577. |
| [7] | 韩冰, 王效科, 逯非, 等. 中国农田土壤生态系统固碳现状和潜力[J]. 生态学报, 2008, 28 (2): 612-619. |
| [8] | 马超, 周静, 刘满强, 等. 秸秆促腐还田对土壤养分及活性有机碳的影响[J]. 土壤学报, 2013, 50 (5): 915-921. |
| [9] | Baumann K, Marschner P, Smernik R J, et al. Residue chemistry and microbial community structure during decomposition of eucalypt, wheat and vetch residues [J]. Soil Biology and Biochemistry, 2009, 41 (9): 1966-1975 |
| [10] | 史学军, 潘剑君, 陈锦盈, 等. 不同类型凋落物对土壤有机碳矿化的影响[J]. 环境科学, 2009, 30 (6): 254-259. |
| [11] | 张红, 吕家珑, 曹莹菲, 等. 不同植物秸秆腐解特性与土壤微生物功能多样性研究[J]. 土壤学报, 2014, 51 (4): 743-752. |
| [12] | Austin A T, Ballare C L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107 (10): 4618-4622. |
| [13] | Walela C, Daniel H, Wilson B, et al. The initial lignin: nitrogen ratio of litter from above and below ground sources strongly and negatively influenced decay rates of slowly decomposing litter carbon pools [J]. Soil Biology and Biochemistry, 2014, 77: 268-275. |
| [14] | Gul S M, Whalen J K, Ellis B E, et al. Plant residue chemistry impacts soil processes and microbial community structure: A study with Arabidopsis thaliana cell wall mutants [J]. Applied Soil Ecology, 2012, 60: 94-91 |
| [15] | Chivenge P, Vanlauwe B, Gentile R, et al. Comparison of organic versus mineral resource effects on short-term aggregate carbon and nitrogen dynamics in a sandy soil versus a fine textured soil [J]. Agriculture Ecosystems and Environment, 2011, 140 (3-4): 361-371 |
| [16] | Fang M, Motavalli P P, Kremer R J, et al. Assessing changes in soil microbial communities and carbon mineralization in Bt and non-Bt corn residue-amended soils [J]. Applied Soil Ecology, 2007 37 (1-2): 150-160. |
| [17] | Gul S M, Whalen J K. Plant life history and residue chemistry influences emissions of CO2 and N2O from soil perspectives for genetically modified cell wall mutants [J]. Plant Sciences, 2013, 32 (5): 344-368. |
| [18] | Abiven S, Heim A, Schmidt M W I. Lignin content and chemical characteristics in maize and wheat vary between plant organs and growth stages: consequences for assessing lignin dynamics in soil [J]. Plant and Soil, 2011, 343 (1-2): 369-378. |
| [19] | 刘四义, 贾淑霞, 张晓平, 等. 玉米和大豆秸秆还田对黑土微生物量及呼吸的影响[J]. 土壤与作物, 2014, 3 (3): 105-111. |
| [20] | Van Soest P J, Robertson J B, Lewis B A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition[J]. Journal of Dairy Science, 1991, 74 (10): 3583-3597. |
| [21] | 刘四义, 张晓平, 贾淑霞, 等. 一种室内培养条件下采用封闭动态系统测定培养物质的CO2产生速率的方法[P]. 中国专利: CN 103776971, 2014-05-07. |
| [22] | 鲁如坤. 土壤农化分析[M]. 北京: 中国农业科技出版社, 2000. 228-235. |
| [23] | Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C [J]. Soil Biology and Biochemistry, 1987, 19 (6): 703-707. |
| [24] | Jenkinson D S. Determination of microbial biomass carbon and nitrogen in soil [A]. In: Wilson J R, (ed.). Advances in Nitrogen Cycling in Agricultural Ecosystems. Wallingford: C. A. B International, 1988. 368-386. |
| [25] | Shi X H, Zhang X P, Yang X M, et al. Contribution of winter soil respiration to annual soil CO2 emission in a Mollisol under different tillage practices in northeast China [J]. Global Biogeochemical Cycles, 2012, 26 (2): GB2007. |
| [26] | Fang Y, Singh B, Singh B P. Effect of temperature on biochar priming effects and its stability in soils[J]. Soil Biology and Biochemistry, 2015, 80: 136-145. |
| [27] | 王晓玥, 孙波. 植物残体分解过程中微生物群落变化影响因素研究进展[J]. 土壤, 2012, 44 (3): 353-359. |
| [28] | Rui J P, Peng J J, Lu Y H. Succession of bacterial populations during plant residue decomposition in rice field soil [J]. Applied and Environmental Microbiology, 2009, 75 (14): 4879-4886 |
| [29] | Johnson J M F, Barbour N W, Weyers S L. Chemical composition of crop biomass impacts its decomposition [J]. Soil Science Society of America Journal, 2007, 71 (1): 155-162. |
| [30] | Yanni S F, Whalen J K, Simpson M J, et al. Plant lignin and nitrogen contents control carbon dioxide production and nitrogen mineralization in soils incubated with Bt and non-Bt corn residues[J]. Soil Biology and Biochemistry, 2011, 43 (1): 63-69. |
| [31] | Kleber M, Sollins P, Sutton R. A conceptual model of organo-mineral interactions in soils: self-assembly of organic molecular fragments into zonal structures on mineral surfaces[J]. Biogeochemistry, 2007, 85 (1): 9-24. |
| [32] | Kuzyakov Y, Bol R. Sources and mechanisms of priming effect induced in two grassland soils amended with slurry and sugar [J]. Soil Biology and Biochemistry, 2006, 38 (4): 747-758. |
| [33] | Ros G H, Hoffland E, Temminghoff E J M. Dynamics of dissolved and extractable organic nitrogen upon soil amendment with crop residues [J]. Soil Biology and Biochemistry, 2010, 42 (12): 2094-2101. |
| [34] | 李娟, 赵秉强, 李秀英, 等. 长期有机无机肥料配施对土壤微生物学特性及土壤肥力的影响[J]. 中国农业科学, 2008, 41 (1): 144-152. |
| [35] | Eskelinen A, Stark S, Männistö M. Links between plant community composition, soil organic matter quality and microbial communities in contrasting tundra habitats [J]. Oecologia, 2009, 161 (1):113-123. |
| [36] | 黄昌勇, 徐建明. 土壤学[M]. (第三版). 北京: 中国农业出版社, 2010. 195-202. |
| [37] | Chen B Q, Liu E K, Tian Q, et al. Soil nitrogen dynamics and crop residues. A review [J]. Agronomy for Sustainable Development, 2014, 34 (2): 429-442. |
| [38] | Henriksen T M, Breland T A. Carbon mineralization, fungal and bacterial growth, and enzyme activities as affected by contact between crop residues and soil [J]. Biology and Fertility of Soils, 2002, 35 (1): 41-48. |
| [39] | 潘剑玲, 代万安, 尚占环, 等. 秸秆还田对土壤有机质和氮素有效性影响及机制研究进展[J]. 中国生态农业学报, 2013, 21 (5): 526-535. |
| [40] | 林先贵, 胡君利. 土壤微生物多样性的科学内涵及其生态服务功能[J]. 土壤学报, 2008, 45 (5): 892-900. |
| [41] | 黄云凤, 张珞平, 洪华生, 等. 小流域氮流失特征及其影响因素[J]. 水利学报, 2006, 37 (7): 801-806, 812. |
| [42] | 陈维梁, 高扬, 林勇明, 等. 紫色土坡耕地氮淋溶过程及其环境健康效应[J]. 环境科学, 2014, 35 (6): 2129-2138. |