 2015, Vol. 36
2015, Vol. 36



赤潮是一种常见的海洋灾害,严重影响着中国沿岸海域[1]. 近年来,随着陆源营养物质的输入,长江口邻近海域富营养化程度越来越严重,赤潮频繁发生[2,3]. 有关赤潮的生态学研究在此相继展开[4~6],然而,赤潮发生时该海域浮游植物的光学性质研究鲜有报道. 丘仲锋等[7,8]对东海赤潮高发区水体的光谱吸收特性进行总体分析,但未将赤潮与非赤潮水体区别开来,且未判别赤潮类型. 雷惠[9]分析了东海赤潮高发区赤潮和非赤潮水体的固有光学性质,并基于固有光学参数差异对硅、 甲藻赤潮进行遥感识别,然而对造成硅、 甲藻赤潮之间和赤潮与非赤潮水体之间光吸收特性差异的原因尚未进行深入分析. 浮游植物的光吸收特性是重要的固有光学量,也是利用海洋水色遥感监测赤潮的基础参量之一[10]. 受长江径流影响,长江口及邻近海域泥沙含量大,浑浊度高,水色组分随长江径流和潮周期的季节变化呈现不均匀的时空分布特征[11],传统的叶绿素遥感反演算法在此应用受到限制[12],浮游植物光吸收特性研究已然成为提高该海域水色组分的反演精度,精确定义和表达生物-光学模型和固有光学量参数化的迫切需要.
鉴于此,本研究依据海上现场调查数据,对长江口邻近海域赤潮水体浮游植物光吸收特性进行分析,并探究赤潮与非赤潮水体浮游植物光吸收特性的差异.
1 材料与方法本研究依托国家基金委长江口共享航次,搭载“科学三号”科学考察船,对长江口邻近海域开展生物-光学调查,调查时间为2013年8月17日至28日,调查范围为122.2°~124.2°E,29.9°~32.1°N. 通过航次期间进行的现场原位测量,样品收集和过滤,以及返回陆地实验室进行的样品测试,获得了较为综合而全面的生物-光学数据集. 基于研究目标,本文着重分析航次进行期间赤潮水体浮游植物的吸收系数和基础水文生化参数.
利用集成有Seabird CTD传感器的Niskin莲座型采水器,现场采集表层水样,并按照拟测量参数的不同依次处理. 收集水样完毕后,立即在船上的实验室进行过滤操作或其他处理. 盐度和温度剖面数据由RBRconcerto多参数水质仪获得,营养盐和溶解氧浓度剖面数据由中国科学院海洋生态与环境科学重点实验室采集和提供.
1.1 颗粒物吸收系数的测定对200~1 000 mL水样进行低压抽滤,颗粒物富集在直径为25 mm、 0.7 μm的Whatman GF/F玻璃纤维滤膜上. 富集着颗粒物的滤膜被迅速置于Corning细胞培养皿中,用铝箔封闭并放入液氮中保存. 返回陆地后,利用带积分球的Lambda 1050双光束紫外-可见-近红外分光光度计测量颗粒物吸收光谱,扫描波长范围为300~1 000 nm,分辨率为2 nm. 吸收系数计算参照NASA海洋光学规范[13],光程放大因子取4.5[14,15]. 浮游植物吸收系数[phytoplankton absorption coefficients,aph(λ)]即为颗粒物总吸收系数与甲醇萃取褪掉色素后的非藻类颗粒吸收系数之差. 浮游植物比吸收系数[phytoplankton-specific absorption coefficients,aph*(λ)]定义为aph(λ)与总叶绿素a浓度(Total chlorophyll-a concentration,[TChla])(测量方法详见1.2节)[16]之比.
1.2 色素浓度的测定及浮游植物粒级结构分析对200~2 000 mL水样进行低压抽滤,将颗粒物富集在0.7 μm Whatman GF/F玻璃纤维滤膜上,迅速对折滤膜,并用铝箔包好,制成色素样品并转移至液氮中. 少量站位取200~2 000 mL水样分别通过20 μm筛绢、 2 μm Millipore聚碳酸酯膜和0.7 μm Whatman GF/F膜,迅速折叠筛绢和滤膜,其他步骤同上,制成分级叶绿素a样品,其中>20 μm为小型浮游植物(microplankton,Micro-)叶绿素a,2~20 μm为微型浮游植物(nanoplankton,Nano-)叶绿素a,0.7~2 μm为超微型浮游植物(picoplankton,Pico-)叶绿素a. 返回陆地后,用Hitachi F-4500荧光分光光度计测量叶绿素a浓度(chlorophyll-a concentration,[Chla])和脱镁叶绿素a浓度[17]. [TChla]为[Chla]和脱镁叶绿素a浓度之和. 测量前,使用Sigma 叶绿素a标准品对荧光分光光度计定标. 少量站位的样品使用Waters 2695 高效液相色谱仪(high performance liquid chromatograph,HPLC)结合二极管阵列检测器和反相C8色谱柱测试色素浓度[13],测试的色素有岩藻黄素、 多甲藻素、 19己酰基氧化岩藻黄素、 19丁酰基氧化岩藻黄素、 别藻黄素、 叶绿素b、 玉米黄素、 叶绿素c2和叶绿素a. 色素标准品从DHI-14C公司购买. 由HPLC测量的叶绿素a浓度表示为[Chla]HPLC.
采用粒径指数[size index,SI]描述浮游植物细胞粒径的变化趋势,SI定义为[16]:
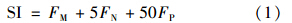
取500 mL水样于大口聚乙烯瓶中,立即加中性甲醛溶液(终浓度为1%),密封后避光保存. 测量前,将样品沉降48 h,去上清液并浓缩至100 mL后,使用Olympus BX43光学显微镜×200和×400倍下鉴别浮游植物物种和细胞计数.
2 结果与分析研究站位如图 1所示. 通过观测人员对现场水体水色的肉眼观察并结合叶绿素a数据([Chla]>10 mg ·m-3[9, 18]),确定B1~B10共10个赤潮站位. 赤潮站位主要位于长江口东部的口外海滨及其东北部浅水区,这与往年长江口邻近海域夏季叶绿素高值区的调查结果一致[19, 20, 21]. 此外,本研究还选取了O1~O10和Z1~Z14共24个非赤潮站位以方便研究结果对比. 为此,将研究区域划分为3个子区域:B1~B10称为B区,O1~O13称为O区,Z1~Z14称为Z区. 3个子区域有明显的地理位置差别:B区水深小于50 m且在30.5°N以北(B5除外); O区水深大于50 m且在123°E以东,远离海岸; Z区位于123°E以西和30.5°N以南的浙江海域.
 | 图 1 2013年8月长江口邻近海域调查站位Fig. 1 Investigation stations in adjacent waters of the Changjiang estuary during August 2013 |
| 表 1 赤潮站位及优势物种Table 1 Phytoplankton bloom stations and their dominant species |
受长江冲淡水和海水的共同影响,研究海域表层盐度(sea surface salinity,SSS)在15.48~33.64之间. 由图 2(a)可以看出,长江冲淡水分两股向外扩展,一股朝东北偏北,一股朝南,且前者扩展的强度大于后者. 由于航次季节为夏季,海表温度 (sea surface temperature,SST)较高,在23.89~29.32℃之间. 由图 2(a)和2(b)可以发现,长江口外南部海域存在一高盐冷水中心,该中心大体位于122.50°E、 30.57°N,SSS和SST达到31.41和23.89℃,其周围水体在水平方向的温度梯度十分明显,表明该海域存在上升流. 台湾暖流的北上入侵可能是该海域上升流形成的重要原因之一. 叶绿素高值区位于B区,与长江冲淡水的扩展方向和上升流的影响范围较为一致,其[Chla]和[TChla]分别在5.63~37.73 mg ·m-3和6.27~65.88 mg ·m-3范围内,最高值出现在B1站位; Z区[Chla]和[TChla]分别在0.92~2.82 mg ·m-3和1.31~3.66 mg ·m-3范围内; O区在0.24~0.54 mg ·m-3和0.39~0.74 mg ·m-3之间. 根据Bricaud等[16]对水体营养程度的划分标准,B区和Z区为富营养化水体,而O区为中等营养水体. 整体上,[Chla]由西北向东南降低,并与盐度呈负相关[Chla]=-2.28 SSS+76.66 (Pearsons r=-0.92,N=32,ANOVA P<0.000 1). 分级叶绿素a在不同水体所占的比例明显不同[图 2(d)]:B、 Z区分别以Micro-和Nano-为主,而O区除O1的Nano-和Pico-均较高外,其余站位以Pico-为主. SI在B、 Z和O区的平均值分别为40.8、 11.9和9.3 μm,也表明由B区向Z区再向O区过渡时小粒径浮游植物所占比例的抬升. SI随[Chla]的升高而呈现幂函数增大的关系(SI=2.157[Chla]0.055,R2=0.78,N=16,ANOVA P<0.000 1). 表层水体溶解氧DO浓度均大于4 mg ·L-1,并在B2和B10处超过10 mg ·L-1,这是浮游植物光合作用旺盛的结果. 与之相对,B区底层呈现缺氧状态[图 3(d)],B1站位距底11 m处发现溶解氧最低值(0.55 mg ·L-1). 低氧区的形成与赤潮水体表层浮游植物产生的大量有机物质在底层水体的氧化分解密切相关.
 | 图 2 SSS、 SST、 表层[Chla]和分级叶绿素a浓度的相对比例的水平分布Fig. 2 Horizontal distributions of salinity,temperature,surface [Chla] and proportions of size-fractioned chlorophyll a concentration
(a)SSS,(b)SST(℃),(c)表层[Chla](mg ·m-3),(d)分级叶绿素a浓度的相对比例 |
除夏季适宜的温度和光照条件外,由长江冲淡水携带和上升流涌升的营养盐是赤潮发生的物质基础. 赤潮水体无机氮和硅酸盐的含量较高[图 3(a)和3(c)],表层浓度范围分别为13.39~44.40 μmol ·L-1和1.81~43.23 μmol ·L-1,氮和硅不对浮游植物生长造成限制. 表层磷酸盐含量较低,浓度在0.17~0.61 μmol ·L-1之间,这与长江冲淡水的磷酸盐浓度相对偏低有关[22],叶绿素最大值处磷酸盐浓度降到0.17 μmol ·L-1的最低值,说明浮游植物生长对磷酸盐的大量消耗[23]; 而下、 底层水磷酸盐浓度的抬升[图 3(b)]可能来自台湾暖流的补充[22].
 | X轴取对数坐标,各标注刻度的数值分别代表 2×10-1、 5×10-1、 100、 2×100、 5×100、 101、 2×101和5×101图 3 营养盐和溶解氧的垂向分布随表层[Chla]的变化Fig. 3 Vertical distributions of nutrients and dissolved oxygen with variations of surface chlorophyll-a concentration |
赤潮水体的浮游植物有着不同于非赤潮水体的吸收系数. 图 4展示了aph(440)的空间分布. 可以看出,3个子区域aph(440)的大小关系为O区
 | 图 4 aph(440)的空间分布Fig. 4 Spatial distributions of aph(440) |
| 表 2 浮游植物在440 nm处的吸收系数与[Chla]的幂函数模型拟合系数及与其他模型的对比 1)Table 2 Regression coefficients at 440 nm of power law function of phytoplankton and comparison with other models |
 | X、 Y轴取对数坐标图 5 赤潮水体aph(440)与[Chla]的关系及与其他模型的比较Fig. 5 Relationships between aph(440) and [Chla] during phytoplankton blooms and comparison with other models |
与吸收系数相比,aph*(440)和aph*(675)变化较小,赤潮水体分别在0.012~0.036 m2 ·mg-1和0.009~0.023 m2 ·mg-1之间,最低值出现在B1站,平均值分别为0.023 m2 ·mg-1和0.015 m2 ·mg-1,这与Brewin等[26]提出的三粒级模型对于Micro-比吸收系数的反推结果接近(图 6). 非赤潮水体二者分别在0.021~0.066 m2 ·mg-1和0.012~0.026 m2 ·mg-1之间,最高值出现在O5站,平均值为0.035 m2 ·mg-1和0.016 m2 ·mg-1,高于赤潮水体. 总体上,aph*(440)有随[Chla]增加而减小的趋势[aph*(440)=0.034[Chla]-0.193,R2=0.43,N=34,ANOVA P<0.000 1]. 如图 6,3个子区域比吸收系数平均值在蓝光波段的大小关系为O区>Z区>B区. 比吸收系数的蓝红波段比值aph*(440)/aph*(675)(blue-to-red ratio,B/R)在B、 Z和O区的平均值分别为1.51、 1.96和2.37,且随[Chla]的增加有减小的趋势(B/R=2.084[Chla]-0.130,R2=0.43,N=34,ANOVA P<0.000 1).
 | 3个分区(实测)和3个分级(Brewin等[26]模型)的平均比吸收系数图 6 平均比吸收系数的比较Fig. 6 Comparison of phytoplankton mean specific absorption coefficients |
而在B区内部,赤潮类型不同,浮游植物比吸收系数也不尽相同. 如图 7,从光谱形状上看,赤潮站位除在440 nm 和675 nm处存在吸收峰外,在495 nm附近存在较不明显的吸收肩峰. 同时,B7、 B9和B10站在465 nm附近存在另一个吸收肩峰,对应东海原甲藻赤潮; 而硅藻赤潮和其余未知类型赤潮站位在465 nm附近没有吸收峰. 从数值上看,甲藻赤潮的aph*(λ)在460~680 nm内均大于其他类型赤潮.
 | 图 7 赤潮站位浮游植物比吸收光谱Fig. 7 Phytoplankton specific absorption spectra of bloom stations |
从图 2(d)及SI的变化可知,由B、 Z向O区过渡时,大粒径浮游植物所占的比例减小,浮游植物由Micro-到Nano-到Pico-过渡,比吸收系数相应升高. 与Brewin等[26]对三粒级比吸收系数的模拟结果进行比较(图 6):B区的Micro-占绝对优势,其aph*(λ)的平均值与Micro-比吸收系数的模拟值几乎一致; Z区Micro-所占比例较B区有所降低,而Nano-比例升高,因此Z区aph*(λ)的平均值在蓝光和红光波段的数值接近但低于Nano-比吸收系数的模拟值; 而在O区小粒径浮游植物比例进一步升高,且Nano-占有不可忽视的比例,因而其aph*(λ)的平均值整体上大于Nano-而远小于Pico-的比吸收系数模拟值. aph*(440)和aph*(675)随SI的增加而呈现幂函数衰减的趋势,R2分别为0.43和0.25. 可见,浮游植物粒径指数的变化对440 nm和675 nm处的比吸收系数变化的贡献可分别达到43%和25%.
细胞粒径是影响色素打包效应的重要因素[24, 27, 28, 29, 30]. 打包效应对aph*(λ)的影响可用打包效应指数[package effect index,Qa*(λ)]衡量. Qa*(λ)为aph*(λ)与同等色素物质在溶解状态下的比吸收系数[phytoplankton-specific absorption coefficient of cellular matter dispersed into solution,a*sol(λ)]之比[16]. 叶绿素a在675 nm处的吸收峰受辅助色素干扰最小[28,31],也就是说,aph*(675)的变化几乎不受色素组成的影响而只源于叶绿素a打包效应[32],取a*sol(675)为0.020 7 m2 ·mg-1[29],计算得到B、 Z和O区Qa*(675)的范围分别为0.44~1.12、 0.59~1.20 和0.65~1.24,在剔除数值超过1的情况后[16],3个分区Qa*(675)的均值分别为0.65、 0.70和0.82. 这表明,赤潮水体的色素打包效应强于非赤潮水体,总体上,B、 Z和O区Qa*(675)变化约有35%、 30%和18%是由打包效应造成的. Qa*(675)随SI的增大而具有幂函数减小的趋势(R2=0.25). 在远离近岸的O区,随浮游植物细胞粒径减小而减小的打包效应比B区赤潮水体和Z区近岸水体小1.5~2倍,因此相应的比吸收系数较大.
事实上,浮游植物粒径与色素浓度和组成在某种程度上存在共变,因此,它对比吸收系数的影响往往可以代表打包效应和色素组成对比吸收系数的共同作用. 一般地,在高叶绿素浓度的水体,大粒径浮游植物占总生物量比例较高[33, 34, 35],打包效应较强,辅助色素占总色素浓度的比例较小[36,37],这些因素共同造成大粒径浮游植物具有低aph*(λ)[28],反之亦然.
3.2 浮游植物色素组成对aph*(λ)的影响B区各站位浮游植物粒径范围一致,均为Micro-占优,具有非常接近的SI,但比吸收光谱形状仍有各自明显的特征(图 7),肩峰的差异与辅助色素有关. 东海常见赤潮硅藻和甲藻的特征色素分别为岩藻黄素和多甲藻素[18, 38, 39],这两种色素均在490 nm附近存在吸收峰,但后者还在460 nm附近有吸收峰[18, 40, 41]. 同时,东海常见赤潮硅藻和甲藻的辅助色素——硅甲藻黄素和叶绿素c2也在460 nm处存在吸收峰[40],然而,二者的浓度之和与[Chla]HPLC的比值不同:硅藻赤潮站位(B1、 B2、 B5和B7)的范围为0.02~0.06,而甲藻赤潮站位(B8和B9)的范围为0.16~0.19,硅藻远小于甲藻,这与许多研究一致[18, 38, 39]. 由此推断,甲藻在465 nm处的吸收肩峰是多甲藻素、 硅甲藻黄素和叶绿素c2共同作用的结果.
4 结论(1)赤潮水体与非赤潮水体的浮游植物吸收光谱不同. 浮游植物吸收系数变化很大,440 nm处赤潮水体和非赤潮水体吸收系数范围分别为0.199~0.832 m-1和0.012~0.109 m-1; 而比吸收系数变化相对较小,440 nm处比吸收系数在赤潮和非赤潮水体的平均值分别为0.023 m2 ·mg-1和0.035 m2 ·mg-1.
(2)浮游植物粒径是影响比吸收系数的重要因素. 从赤潮水体向非赤潮水体过渡,大粒径浮游植物所占比例减小,小粒径浮游植物所占比例上升,打包效应减小,因而比吸收系数升高. 浮游植物粒径指数的变化对440 nm和675 nm处的比吸收系数变化的贡献可分别达到43%和25%.
(3)甲藻和硅藻色素组成差异对浮游植物比吸收系数有着重要影响. 甲藻赤潮中硅甲藻黄素和叶绿素c2的浓度之和与叶绿素a浓度的比值大于硅藻赤潮,是甲藻在465 nm附近出现吸收肩峰的重要原因.
致谢: 真诚感谢王桂芬博士、 周雯博士、 沈盎绿博士、 余小龙、 陈雨和姚林杰对于数据采集和处理的帮助,以及中国科学院海洋生态与环境科学重点实验室和宋书群博士提供相关同步测量数据.
| [1] | 梁松, 钱宏林, 齐雨藻. 中国沿海的赤潮问题 [J]. 生态科学, 2000, 19 (4): 44-50. |
| [2] | 韩秀荣, 王修林, 孙霞, 等. 东海近海海域营养盐分布特征及其与赤潮发生关系的初步研究 [J]. 应用生态学报, 2003, 14 (7): 1097-1101. |
| [3] | Chai C, Yu Z M, Song X X, et al. The status and characteristics of eutrophication in the Yangtze River (Changjiang) Estuary and the adjacent East China Sea, China [J]. Science of the Total Environment, 2009, 407 : 4687-4695. |
| [4] | 叶属峰, 纪焕红, 曹恋, 等. 长江口海域赤潮成因及其防治对策 [J]. 海洋科学, 2004, 28 (5): 26-32. |
| [5] | 周名江, 于仁成. 有害赤潮的形成机制、危害效应与防治对策 [J]. 自然杂志, 2007, 29 (2): 72-77. |
| [6] | 周名江, 朱明远, 张经. 中国赤潮的发生趋势和研究进展 [J]. 生命科学, 2001, 13 (2): 54-59. |
| [7] | 丘仲锋, 席红艳, 何宜军, 等. 东海赤潮高发区半分析算法色素浓度反演 [J]. 环境科学, 2006, 27 (8): 1516-1521. |
| [8] | 丘仲锋, 何宜军, 唐军武, 等. 东海赤潮高发区水体光谱吸收特性分析 [J]. 海洋科学集刊, 2007, 48 : 1-14. |
| [9] | 雷惠. 基于固有光学量的东海赤潮遥感提取算法研究[D]. 杭州: 浙江大学, 2011. 25-133. |
| [10] | 赵冬至. 赤潮灾害卫星遥感探测技术 [M]. 北京: 海洋出版社, 2009. 1-12. |
| [11] | Shen F, Zhou Y X, Hong G L. Absorption property of non-algal particles and contribution to total light absorption in optically complex waters, a case study in Yangtze Estuary and adjacent coast [A]. In: Advances in Computational Environment Science[C]. Berlin Heidelberg: Springer-Verlag, 2012. 61-66. |
| [12] | Cui T W, Zhang J, Tang J W, et al. Assessment of satellite ocean color products of MERIS, MODIS and SeaWiFS along the East China Coast (in the Yellow Sea and East China Sea) [J]. ISPRS Journal of Photogrammetry and Remote Sensing, 2014, 87 : 137-151. |
| [13] | NASA/TM-2000-209966. Ocean Optics Protocols For Satellite Ocean Color Sensor Validation, Revision 2 [S]. |
| [14] | 余小龙, 沈芳, 张晋芳. 影响悬浮颗粒物吸收系数测量的相关因素研究 [J]. 环境科学, 2013, 34 (5): 1745-1753. |
| [15] | Röttgers R, Gehnke S. Measurement of light absorption by aquatic particles: improvement of the quantitative filter technique by use of an integrating sphere approach [J]. Applied Optics, 2012, 51 (9): 1336-1351. |
| [16] | Bricaud A, Claustre H, Ras J, et al. Natural variability of phytoplanktonic absorption in oceanic waters: Influence of the size structure of algal populations [J]. Journal of Geophysical Research: Oceans (1978-2012), 2004, 109 (C11): 183-197. |
| [17] | Neveux J, Lantoine F. Spectrofluorometric assay of chlorophylls and phaeopigments using the least squares approximation technique [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 1993, 40 (9): 1747-1765. |
| [18] | 庄宏儒. 水质自动监测系统在厦门同安湾赤潮短期预报中的应用[J]. 海洋环境科学, 2006, 25 (2): 58-61. |
| [19] | 宋书群, 孙军, 沈志良, 等. 三峡库区蓄水后夏季长江口及其邻近水域粒级叶绿素a [J]. 中国海洋大学学报(自然科学版), 2006, 36 (S1): 114-120. |
| [20] | 韦钦胜, 臧家业, 战闰, 等. 夏季长江口东北部上升流海域的生态环境特征 [J]. 海洋与湖沼, 2011, 42 (6): 899-905. |
| [21] | 朱建荣. 长江口外海区叶绿素a浓度分布及其动力成因分析 [J]. 中国科学D辑:地球科学, 2004, 34 (8): 757-762. |
| [22] | Liu K K, Tang T Y, Gong G C, et al. Cross-shelf and along-shelf nutrient fluxes derived from flow fields and chemical hydrography observed in the southern East China Sea off northern Taiwan [J]. Continental Shelf Research, 2000, 20 (4-5): 493-523. |
| [23] | 俞志明, 沈志良. 长江口水域富营养化 [M]. 北京: 科学出版社, 2011. 50-96. |
| [24] | Bricaud A, Morel A, Babin M. Variations of light absorption by suspended particles with chlorophyll a concentration in oceanic (case 1) waters: analysis and implications for bio-optical models [J]. Journal of Geophysical Research: Oceans(1978-2012), 1998, 103 (C13): 31033-31044. |
| [25] | Wang G F, Cao W X, Yang Y Z, et al. Variations in light absorption properties during a phytoplankton bloom in the Pearl River estuary [J]. Continental Shelf Research, 2010, 30 (9): 1085-1094. |
| [26] | Brewin R J W, Devred E, Sathyendranath S, et al. Model of phytoplankton absorption based on three size classes [J]. Applied Optics, 2011, 50 (22): 4535-4549. |
| [27] | Cleveland J S. Regional models for phytoplankton absorption as a function of chlorophyll a concentration [J]. Journal of Geophysical Research, 1995, 100 (C7): 13333-13344. |
| [28] | Bricaud A, Babin M, Morel A, et al. Variability in the chlorophyll-specific absorption coefficients of natural phytoplankton: analysis and parameterization [J]. Journal of Geophysical Research, 1995, 100 (C7): 13321-13332. |
| [29] | Morel A, Bricaud A. Theoretical results concerning light absorption in a discrete medium, and application to specific absorption of phytoplankton [J]. Deep Sea Research Part A. Oceanographic Research Papers, 1981, 28 (81): 1375-1393. |
| [30] | Bricaud A, Stramski D. Spectral absorption coefficients of living phytoplankton and nonalgal biogenous matter: a comparison between the Peru upwelling area and the Sargasso Sea [J]. Limnology and Oceanography, 1990, 35 (3): 562-582. |
| [31] | Stramski D, Morel A. Optical properties of photosynthetic picoplankton in different physiological states as affected by growth irradiance [J]. Deep-Sea Research Part A. Oceanorgraphic Research Papers, 1990, 37 (2): 245-266. |
| [32] | Partensky F, Hoepffner N, Li W K W, et al. Photoacclimation of Prochlorococcus sp. (Prochlorophyta) strains isolated from the north Atlantic and the Mediterranean Sea [J]. Plant Physiology, 1993, 101 (1): 285-296. |
| [33] | Ciotti á M, Lewis M R, Cullen J J. Assessment of the relationships between dominant cell size in natural phytoplankton communities and the spectral shape of the absorption coefficient [J]. Limnology and Oceanography, 2002, 47 (2): 404-417. |
| [34] | Lohrenz S E, Weidemann A D, Tuel M. Phytoplankton spectral absorption as influenced by community size structure and pigment composition [J]. Journal of Plankton Research, 2003, 25 (1): 33-61. |
| [35] | Brewin R J W, Sathyendranath S, Hirata T, et al. A three-component model of phytoplankton size class for the Atlantic Ocean [J]. Ecological Modelling, 2010, 221 (11): 1472-1483. |
| [36] | Babin M, Stramski D, Ferrari G M, et al. Variations in the light absorption coefficients of phytoplankton, nonalgal particles, and dissolved organic matter in coastal waters around Europe [J]. Journal of Geophysical Research Oceans(1978-2012), 2003, 108 (C7): 1-20. |
| [37] | Bricaud A, Babin M, Claustre H, et al. Light absorption properties and absorption budget of Southeast Pacific waters [J]. Journal of Geophysical Research: Oceans (1978-2012), 2010, 115 (C8): 488-507. |
| [38] | Yao P, Yu Z G, Deng C M, et al. Classification of marine diatoms using pigment ratio suites [J]. Chinese Journal of Oceanology and Limnology, 2011, 29 (5): 1075-1085. |
| [39] | Zapata M, Fraga S, Rodríguez F, et al. Pigment-based chloroplast types in dinoflagellates [J]. Marine Ecology Progress Series, 2012, 465 : 33-52. |
| [40] | Johnsen G, Nelson N B, Jovine R V M, et al. Chromoprotein- and pigment-dependent modeling of spectral light absorption in two dinoflagellates, Prorocentrum minimum and Heterocapsa pygmaea[J]. Marine Ecology Progress Series, 1994, 114 : 245-258. |
| [41] | Bidigare R R, Ondrusek M E, Morrow J H, et al. In vivo absorption properties of algal pigments [J]. SPIE Ocean Optics X, 1990, 1302 : 290-302. |