 2015, Vol. 36
2015, Vol. 36



2. 中国科学院重庆绿色智能技术研究院, 重庆 400714
2. Chongqing Institute of Green and Intelligent Technology, Chinese Academy of Sciences, Chongqing 400714, China
蓝藻水华是我国淡水水域生态环境面临的典型生态问题[1, 2]. 水华鱼腥藻(Anabaena flosaquae)是蓝藻水华中除微囊藻之外的常见优势种群,具有很强的营养竞争性. 同时,水华鱼腥藻还可分泌藻毒素,对人类和动物神经系统有毒害作用[3]. 有研究表明,在可利用磷浓度较低的河流中,水华鱼腥藻比微囊藻(Microcystis sp.)更容易形成水华[4, 5, 6].
生态因子对水生环境中藻类的生长起着重要作用. 贾晓燕等[7]研究发现氮限制下鱼腥藻生长良好,说明氮不是其限制因子. 王亚超等[8]研究太湖水华鱼腥藻的生长机制,发现当藻类密度较低时,与总磷呈正相关. Nalewajko等[9]对Biwa湖的水华鱼腥藻进行研究,得出其最佳生长温度范围为28~32℃. Venugopal等[10]研究了高光照度(6000~15000 lx)对水华鱼腥藻生长的影响. Zapomělov等[11]研究了8种不同的水华鱼腥藻种对环境因子(光照、 温度)的响应特征.
目前研究多侧重于单一环境因素对鱼腥藻生长的影响研究,但关于磷限制下光、 热变量交互下水华鱼腥藻的生长动力学鲜有报道,多重环境要素影响下的生理响应特征并不明晰. 本研究通过室内机制实验,建立了磷限制下水华鱼腥藻生长的光热模型,初步分析磷限制下水华鱼腥藻对光、 热环境的响应,以期为后续深入研究水华鱼腥藻生长及其水华形成机制提供基础支持[12, 13, 14]. 1 材料与方法 1.1 实验方法
本研究中所有水华鱼腥藻FACHB245(Anabaena flosaquae,以下简称“鱼腥藻”)购自中国科学院水生生物研究所. 实验以BG-11培养基为基础,采用新鲜去离子水配制培养基,初始磷(K2HPO4)的摩尔浓度为5.0 μmol ·L-1,pH值调节至7.5左右. 将以上培养基放入灭菌锅,用121℃ 高压蒸汽灭菌30 min.
参照野外监测分析结果,本实验设置15、 20、 25、 30℃ 这4个温度水平和1000、 2000、 3000、 5000 lx这4个光照水平,研究所设定的磷浓度下光热交互作用对鱼腥藻生长的影响. 鱼腥藻培养采用序批式培养,每次实验均采用单因素变量,实验设计如表 1所示.
| 表 1 环境变量实验设计表 Table 1 Experimental design of environment variables |
实验过程中的主要仪器与设备有:光照培养箱、 生物光学显微镜等. 实验需要的玻璃仪器用超声波处理器洗净后,在10%稀盐酸中浸泡24 h,然后用新鲜去离子水冲洗干净,用121℃ 高压蒸汽灭菌,烘干后备用.
藻种实验前扩大培养1周,再饥饿培养48 h. 取一定体积的藻种以4200 r ·min-1的速度离心15 min,弃掉上清液,用15 mg ·L-1的碳酸氢钠溶液洗涤后离心,重复3次,用无菌水稀释后用于接种.
本实验采用动力学中的“分批培养实验”,每批实验的光照、 温度按照表 1所设置,光暗比保持为12 h ∶12 h. 藻类样品培养装置采用250 mL三角瓶,培养期间每天人工摇动3~4次. 1.3 细胞生物量测定与增长率计算
从接种第一天起,每日上午08:00用微量吸液器吸取1.0 mL的藻类培养液,移至经高压灭菌的加有1.0 mL鲁哥试剂的定量瓶中. 采用浮游植物计数框计数法测定生物量,每个样计数3次,每次50个视野,取其平均数计算藻密度,并计算计数误差棒.
增长速率(μ)是指藻细胞在单位时间内所增加的藻细胞质量[15],其计算如公式(1):
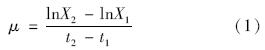
光照、 温度交互作用下的动力学模型[16,17]如式(2):

实验数据全部录入Origin或SPSS进行相关统计分析,并采用Matlab进行光热模型拟合分析与参数求取.
2 结果与分析 2.1 不同温度、 光照下鱼腥藻细胞数量变化过程
温度从15℃ 上升到30℃,鱼腥藻最大生物量分别为56.75×105、 66.14×105、 75.59×105、 95.87×105 cell ·mL-1. 鱼腥藻最大生物量随温度升高逐渐增大,15℃ 时生物量最小,30℃ 时生物量最大. 在适宜的温度范围内,温度升高有利于鱼腥藻的增殖.
培养期间,不同温度下鱼腥藻细胞数量增殖过程如图 1所示. 在15~30℃ 温度范围内,鱼腥藻生物量显著增加; 从藻类生长周期来看,在1~3 d时,鱼腥藻处于适应期,增殖缓慢; 在4~8 d时,鱼腥藻对营养物质的利用率提高,藻细胞快速增殖,进入对数增长期. 在9~11 d时,鱼腥藻的生物量增长速率有所减缓,甚至降低.
 | 图 1 不同温度下鱼腥藻生物量变化
Fig. 1 Variation of biomass of Anabaena flosaquae
at different temperatures
|
不同光照下鱼腥藻细胞数量增殖过程见图 2. 在1000~5000 lx光照范围内,鱼腥藻生物量均有明显增加,其生物量增长趋势与不同温度条件下基本相似,在1~3 d时,鱼腥藻需要对新环境适应调整,增殖缓慢; 在4~8 d时,鱼腥藻快速增殖,进入对数增长期,营养物质消耗增多. 当光照自1000 lx上升到5000 lx时,鱼腥藻最大生物量分别为53.79×105、 70.78×105、 66.14×105、 40.97×105 cell ·mL-1. 在5000 lx时鱼腥藻生物量最小,2000 lx时生物量最大. 鱼腥藻在低光照水平下(1000 lx)生物积累量较低,在高光照水平下(5000 lx),鱼腥藻生长明显受到抑制,生物量积累水平低于低光照的生物量. 可见,较低或较高光照都不利于鱼腥藻的生长.
 | 图 2 不同光照下鱼腥藻生物量变化
Fig. 2 Variation of biomass of Anabaena flosaquae at different light intensity
|
光照为3000 lx,温度从15~30℃ 时,鱼腥藻生物积累量不断增加. 温度对藻类生长的影响主要体现在细胞内酶促反应速率的变化,并影响藻细胞生长速率. 以整个培养周期(10 d)为边界,计算整个培养周期的平均生长速率,结果如图 3所示.
 | 图 3 不同温度下鱼腥藻平均生长速率比较
Fig. 3 Comparison of the average growth rate of
Anabaena flosaquae at different temperatures
|
温度在15~30℃ 范围内,鱼腥藻平均生长速率分别为0.3556、 0.4454、 0.4027、 0.3754 d-1; 在温度为20℃ 时,生长速率最大. 15℃时鱼腥藻平均生长速率最小,同其他 4个温度水平相比,上述差异具有显著性 (One-way ANOVA,P≤0.05). 平均生长速率在20℃达到最高,但随着温度进一步升高,呈现出比较明显的下降趋势. 适宜的温度能够促进鱼腥藻生理活性,如酶的活性、 呼吸强度及合成代谢等; 而温度较高时(高于25℃),鱼腥藻生长速率降低,对数期和稳定期会缩短.
 | 图 4 不同光照下鱼腥藻平均生长速率比较
Fig. 4 Comparison of the average growth rate of
Anabaena flosaquae at different light intensity
|
光照在1000~5000 lx范围内,鱼腥藻生长速率分别为0.3101、 0.3813、 0.4454、 0.2717 d-1,在光照为3000 lx时,生长速率最大. 如图 4所示,光照在2000~3000 lx范围内,鱼腥藻平均生长速率较高,而光照为1000 lx和5000 lx时,平均生长速率处于较低水平; 光照在3000 lx时,平均生长速率最大,5000 lx时生长速率最小,比3000 lx时相比降低了39%,可见光照度对藻的生长影响显著(One-way ANOVA,P≤0.05). 适宜的光照辐射为藻类光合作用提供代谢的能源、 提高光合碳固定速率; 而光照处于较高水平(>5000 lx),超过光合作用的饱和点时,藻的光能利用效率降低,生长受到抑制,生物积累量下降. 2.3 光热交互作用下鱼腥藻生长对光热条件的响应
光照、 温度作为藻类细胞能量输入的重要指标,单一分析各自变化对鱼腥藻生长影响具有一定局限性,且与实际环境下光照、 温度交互作用存在一定的偏差. 在自然水体中,对春季鱼腥藻而言,磷是其生长的限制因子之外,光照、 温度也是影响鱼腥藻生长的关键因子. 为探究光照、 温度两者交互作用下鱼腥藻生长状况,构建光照、 温度、 生长速率三者之间的动力学模型进行初步分析.
在前述实验结果基础上,并根据式(2)对实验数据进行非线性回归和光热模型拟合. 分析的结果见表 2.
| 表 2 光热模型拟合参数 Table 2 Fitting parameters of the photo-thermal model |
光热模型的拟合结果如图 5所示:在最适光照、 温度下,鱼腥藻的生长速率明显呈现出单峰响应. 当温度恒定时,在低光照下鱼腥藻生长速率不断升高,达到最大值后随光照升高反而逐渐下降. 当光照恒定时,鱼腥藻受温度影响的生长速率响应模式与Lehman温度模型相吻合[18].
 | 图 5 鱼腥藻光热模型拟合
Fig. 5 Nonlinear fitting of the photo-thermal model of Anabaena flosaquae
|
藻类吸收营养物质的效率和利用程度和呼吸代谢等生理特性,以及细胞内酶促反应都受到温度的影响. 在适宜的温度环境中,鱼腥藻种群增长速率高,有利于种群在短期内快速增殖形成优势藻种. 磷限制下,温度为30℃ 时的生长速率值略小于25℃ 时的数值(t-检验,P≥0.05),分析可能是因为温度升高引起藻细胞内部某些酶类活性降低或膜脂和蛋白及类囊体膜结构和功能发生变化,导致鱼腥藻代谢能力略有下降[19].
常锋毅等[20]研究发现螺旋鱼腥藻生长最适的生长温度为 20~25℃,与本实验的结果基本一致. Liu等[21]研究发现25~30℃ 是微囊藻生长的最适温度范围,略高于鱼腥藻,分析应该是不同藻种的生理特性差异造成的. Converti等[22]培养微绿球藻时发现,在20~25℃ 内微绿球藻的生长速率处于高水平,且其脂质含量从7.90%增加到14.92%,说明微绿球藻的生理特性与鱼腥藻有一定相似. 在20~25℃ 内鱼腥藻生长速率始终维持在较高水平,有利于种群在短期内大量增殖迅速形成优势藻种. 这一温度范围接近春夏期间水体实际温度,为春末夏初鱼腥藻水华出现创造了适宜的温度条件. 3.2 磷限制下不同光照对鱼腥藻生长影响的分析
适宜的光照环境有助于鱼腥藻通过提高光合色素含量和碱性磷酸酶活性来适应低磷环境. 大量研究表明:鱼腥藻在低光照环境下,能够通过降低细胞含糖量并合成新的伪空胞增加藻细胞浮力,使藻细胞上浮至最适光照水层[23]. 当光照足够充足(高于3000 lx)时,藻细胞光合速率提高,光合产物不断累积(如糖、 蛋白质),细胞密度逐渐增大,此时伪空胞提供的浮力不足以支撑整个细胞而下沉至合适水层,可见鱼腥藻对光照有很强的适应性[24]. 光照高于5000 lx时,鱼腥藻的生长速率只有0.2717 d-1,分析原因可能是光照度超过了鱼腥藻的光饱和点,产生强光抑制作用,或是藻细胞下沉到过低水层导致光能利用效率降低,最终鱼腥藻生长速率降低.
Imaizumi等[25]通过对小球藻进行高细胞密度连续培养,结果表明:光照为28~45 μmol ·(m2 ·s)-1时生长速率能够达0.5 d-1,大于本实验中生长速率最大值0.4454 d-1. Litchman等[26]发现在20℃ 下鱼腥藻最佳生长光照约为40 μmol ·(m2 ·s)-1(≈3200 lx),其光照抑制常数Kinh高达61μmol ·(m2 ·s)-1(≈4900 lx),该结果与本实验结果比较接近. 本实验在磷限制环境下进行,磷对鱼腥藻的生长胁迫可能会影响光合作用中光反应阶段ATP的合成. 春末夏初时,太阳辐射一般处于较高水平,水层表面光照较高,远高于鱼腥藻的最适生长光照(大于3000 lx),鱼腥藻依旧能够繁殖,甚至形成水华[27]. 造成这种情况的原因可能在于太阳辐射光照不能完全被水体吸收,水体中实际光照度小于所测太阳辐射值. 3.3 光热模型分析
从光热模型拟合效果来看,在温度15~25℃和光照2000~3000 lx范围内,鱼腥藻的生长速率处于相对较高水平. 光热模型表明鱼腥藻最适生长的光照、 温度环境条件较为恒定,即21.03℃±1.55℃、 2675.12 lx±262.93 lx,该拟合参数值接近春末夏初水体实际的光热条件. 在淡水河流藻类模型中,Monod模型[28]能够较好地揭示营养盐浓度与藻类生长之间的动力学关系,但缺乏环境因子连续变化对藻类生长影响的分析; Lehman模型用来描述温度、 营养盐浓度和藻类生长速率三者间的连续响应关系; Steele模型[29]描述光照度由低向高连续变化下藻类生长速率响应特征,但其均只是描述单一光热条件下藻类的生长特征. 而本研究的光热模型不仅可以得到磷限制下相应的鱼腥藻生长动力学参数,而且还可以得到鱼腥藻生长速率随温度、 光照交互作用下连续变化的变化趋势. Litchman[26]对营养盐充足下鱼腥藻生长特征展开室内实验,结果表明在20℃、 光照度为35μmol ·(m2 ·s)-1(≈3000 lx)的光热条件下,其生长速率约为0.4000 d-1,与本实验结果相比稍小,分析可能是鱼腥藻更适应低磷环境. 柴小颖[30]通过对三峡库区典型流域两江的浮游藻类的调查研究,并结合实验室模拟的方法,构建了光照、 温度与微囊藻最终生物量的三维模型,发现温度在25℃ 左右时最终生物量始终保持较高水平,并随着光照度增加而呈上升趋势,但当光照超过5200 lx时最终生物量有所下降. 与本研究中鱼腥藻在光热交互下的生长趋势基本一致. Duarte[31]结合光机制模型和温度经验函数,构建了光照、 温度对藻类初级生产力的影响模型,同时也探讨了温度对光照-藻初级生产力的影响,但缺乏对营养盐的讨论.
光热模型拟合的磷限制下鱼腥藻最适生长温度、 光照条件与实际环境的光热条件基本一致,说明模型有一定的实际参考意义. 通过建立磷限制下鱼腥藻生长的光热模型,为深入探究鱼腥藻的生长机制和进一步预测鱼腥藻水华提供重要基础. 但由于在模型的建立过程中,只考虑了光照和温度两个因素的影响,并且自然水体中藻种类繁多和相互竞争影响,该模型的应用存在一定的局限性. 自然水体中各种环境因素不断变化并且不同环境因子相互交错,要更准确地反映实际水体中藻类生长情况,需要进一步的进行研究和论证.
4 结论
(1)光照条件恒定,磷限制下水华鱼腥藻的最适生长温度为20℃; 温度条件恒定,磷限制下水华鱼腥藻的最适生长光照为3000 lx; 但光照度过高,超过水华鱼腥藻光饱和点会抑制鱼腥藻的生长,生物积累量显著减少.
(2)光热模型拟合的最佳生长温度、 最佳生长光照分别为21.03℃±1.55℃、 2675.12 lx±262.93 lx,接近实际水环境的光热条件,要更准确反映实际水体中光热对藻类生长影响,还需要深入探究.
| [1] | Kormas K A, Gkelis S, Vardaka E, et al. Morphological and molecular analysis of bloom-forming Cyanobacteria in two eutrophic, shallow Mediterranean lakes[J]. Limnologica-Ecology and Management of Inland Waters, 2011, 41 (3): 167-173. |
| [2] | O'Neil J M, Davis T W, Burford M A, et al. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change[J]. Harmful Algae, 2012, 14: 313-334. |
| [3] | 胡宗达, 周元清. 水华蓝藻毒素研究概述[J]. 云南环境科学, 2004, 23 (3): 8-11. |
| [4] | 谭香, 沈宏, 宋立荣. 三种水华蓝藻对不同磷浓度生理响应的比较研究[J]. 水生生物学报, 2007, 31 (5): 693-699. |
| [5] | Shi X L, Yang L Y, Wang F P, et al. Growth and phosphate uptake kinetics of Microcystis aeruginosa under various environmental conditions[J]. Journal of Environmental Sciences, 2004, 16 (2): 288-292. |
| [6] | Sotero-Santos R B, Carvalho E G, Dellamano-Oliveira M J, et al. Occurrence and toxicity of an Anabaena bloom in a tropical reservoir (Southeast Brazil)[J]. Harmful Algae, 2008, 7 (5): 590-598. |
| [7] | 贾晓燕, 刘聪, 储昭升, 等. 氮磷限制条件下螺旋鱼腥藻产生土嗅素特征研究[J]. 中国环境科学, 2014, 34 (4): 1026-1030. |
| [8] | 王亚超, 徐恒省, 王国祥, 等. 氮、磷等环境因子对太湖微囊藻与水华鱼腥藻生长的影响[J]. 环境监控与预警, 2013, 5 (1): 7-10. |
| [9] | Nalewajko C, Murphy T P. Effects of temperature, and availability of nitrogen and phosphorus on the abundance of Anabaena and Microcystis in Lake Biwa, Japan: an experimental approach[J]. Limnology, 2001, 2 (1): 45-48. |
| [10] | Venugopal V, Prasanna R, Sood A, et al. Stimulation of pigment accumulation in Anabaena azollae strains: Effect of light intensity and sugars[J]. Folia Microbiologica, 2006, 51 (1): 50-56. |
| [11] | Zapomělová E,Řeháková K, Jezberová J, et al. Polyphasic characterization of eight planktonic Anabaena strains (Cyanobacteria) with reference to the variability of 61 Anabaena populations observed in the field[J]. Hydrobiologia, 2010, 639 (1): 99-113. |
| [12] | Wang Y J, Li J, Lü Y, et al. Effects of cerium on growth and physiological characteristics of Anabaena flosaquae[J]. Journal of Rare Earths, 2012, 30 (12): 1287-1292. |
| [13] | 刘海陆, 徐颖, 刘建辉, 等. 磷和铁对洋河水库螺旋鱼腥藻生长的影响[J]. 生态科学, 2013, 32 (1): 115-119. |
| [14] | 王正方, 张庆, 吕海燕. 温度、盐度、光照强度和pH对海洋原甲藻增长的效应[J]. 海洋与湖沼, 2001, 32 (1): 15-18. |
| [15] | 郑朔方, 杨苏文, 金相灿. 铜绿微囊藻生长的营养动力学[J]. 环境科学, 2005, 26 (2): 152-156. |
| [16] | Nicklisch A, Shatwell T, Köhler J. Analysis and modelling of the interactive effects of temperature and light on phytoplankton growth and relevance for the spring bloom[J]. Journal of Plankton Research, 2008, 30 (1): 75-91. |
| [17] | Lehman J T, Botkin D B, Likens G E. The assumptions and rationales of a computer model of phytoplankton population dynamics[J]. Limnology and Oceanography, 1975, 20 (3): 343-364. |
| [18] | 周婕, 曾诚. 湖泊富营养化数学模型研究进展[J]. 人民长江, 2007, 38 (11): 115-118. |
| [19] | 郑维发, 曾昭琪. 淡水蓝藻的高温适应[J]. 湖泊科学, 1994, 6 (4): 356-363. |
| [20] | 常锋毅, 潘晓洁, 康丽娟, 等. 洱海螺旋鱼腥藻生长生理特性的初步研究[J]. 水生生物学报, 2009, 33 (3): 385-390. |
| [21] | Liu X, Lu X H, Chen Y W. The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: An 11-year investigation[J]. Harmful Algae, 2011, 10 (3): 337-343. |
| [22] | Converti A, Casazza A A, Ortiz E Y, et al. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production[J]. Chemical Engineering and Processing: Process Intensification, 2009, 48 (6): 1146-1151. |
| [23] | 成慧敏, 邱保胜. 蓝藻的伪空泡及其对蓝藻在水体中垂直分布的调节[J]. 植物生理学通讯, 2006, 42 (5): 974-980. |
| [24] | Karlsson-Elfgren I, Brunberg A K. The importance of shallow sediments in the recruitment of Aanabaena and Aphanizomenon (cyanophyceae)[J]. Journal of Phycology, 2004, 40 (5): 831-836. |
| [25] | Imaizumi Y, Nagao N, Yusoff F M, et al. Estimation of optimum specific light intensity per cell on a high-cell-density continuous culture of Chlorella zofingiensis not limited by nutrients or CO2[J]. Bioresource Technology, 2014, 162 (12): 53-59. |
| [26] | Litchman E. Growth rates of phytoplankton under fluctuating light[J]. Freshwater Biology, 2000, 44 (2): 223-235. |
| [27] | 巫娟, 陈雪初, 孔海南, 等. 光照度对水华鱼腥藻细胞比重与藻丝长度的影响研究[J]. 中国环境科学, 2012, 32 (5): 875-879. |
| [28] | Monod J. La technique de la culture continue, théorie et applications[J]. Annales de I'Institute Pasteur Paris, 1950, 79 (4): 390-410. |
| [29] | Steele J H. Environmental control of photosynthesis in the sea[J]. Limnology and Oceanography, 1962, 7 (2): 137-150. |
| [30] | 柴小颖. 光照和温度对三峡库区典型水华藻类生长的影响研究[D]. 重庆: 重庆大学, 2009. |
| [31] | Duarte P. A mechanistic model of the effects of light and temperature on algal primary productivity[J]. Ecological Modelling, 1995, 82 (2): 151-160. |