 2014, Vol. 35
2014, Vol. 35



2. 上海水生环境工程有限公司, 上海 200090
2. Shanghai Aquatic Environmental Engineering Corporation Limited, Shanghai 200090, China
轮虫是水体中常见的一类后生浮游动物,具有数量大、 种类多、 发育时间短、 周转快、 生产量高等特点,它们对维持水生态系统结构稳定性和功能完整性方面具有重要意义[1,2]. 同时,轮虫对环境变化反应灵敏,其群落结构能够及时地反映水体的环境状况,是良好的环境指示生物[3,4,5,6,7,8,9]. 因此,从生态学角度而言,对不同水体中轮虫群落结构及其与水体环境因子的相互关系进行研究具有十分重要的意义. 目前关于轮虫的研究,以湖泊、 水库、 池塘等水体为对象的占大部分[10,11],而关于河流、 溪流的报道较少,尤其关于河口性新建成的水体中轮虫群落次生演替的研究鲜少报道.
上海临港新城B港河道(于2010年12月人工开挖)位于上海临港新城东南沿海滩涂之中,外连芦潮引河和秋涟河,毗邻杭州湾,属于典型的河口性河道. 本研究于2011年12月-2012年11月逐月对临港B港河道轮虫和其他环境因子进行调查研究,分析了该新建河道轮虫群落结构的动态变化和其与环境因子的相互关系,通过摸清新建河口性河道轮虫群落的次生演替规律,以期为新建河口区河道的水质保护和生态治理提供科学依据. 1 材料与方法 1.1 临港B港河道概况
上海临港B港河道于2011年2月初开挖完成,河道水体总面积约109070 m2,河口宽30-200 m,排涝最高水位3.5 m,正常控制水位2.5-2.7 m,河底标高0.00 m,河道承载着区域400万km2的蓄洪排涝、 雨水收集以及景观生态服务功能. 该河道在2011年7月下旬至9月期间实施了水生植被生态恢复工程,在此期间种植了睡莲(Nymphaea alba)、 黄菖蒲(Lris pseudacorus)、 苦草(Vallisneria natans)、 穗花狐尾藻(Myriophyllum spicatum)、 再力花(Thalia dealbata)、 梭鱼草(Pontederia cordata)等20种水生植物,植被恢复面积约为25000 m2,由于临港B港河道刚刚建成,水体未完全开放,水流速度为0 m ·s-1.
根据临港B港河道形态及水文特征,在河道内设置4个采样点(见表 1和图 1),S1为B港(靠近水闸内侧),S2为靠近HY-8路,S3为靠近HY-12路,S4为秋涟河附近; 另外,在B港的外河水系芦潮引河(水闸外侧)设置一个对照采样点S0,在杭州湾(水闸内侧)设置一个对照采样点S0 . 于2011年12月-2012年11月,逐月采样监测临港B港河道轮虫及其水质状况(2月、 10月由于天气原因未采样),对照点S0 主要在春季和夏季(3-6月)进行了采样.
 | 图 1 临港B港河道采样点设置示意Fig. 1 Sampling sites in the Lin gang B Gang River |
轮虫定性样品:用10 μm浮游生物网采集,装入两个50 mL的样品瓶中,一瓶用4%福尔马林溶液固定,另一瓶用于活体观察; 某些轮虫的鉴定,应用5%的NaOH溶液对轮虫进行消化,以观察其咀嚼器的形态特征[12,13,14,15,16].
轮虫定量样品:用5L有机玻璃采水器采集距表层0.5 m和距水底0.5 m的水样,经混匀后,取1 L混合水样装入聚乙烯瓶,经4%的福尔马林溶液固定后,浓缩至50 mL; 定量计数时,取1 mL于计数框中在10×10倍显微镜下全片计数轮虫密度(3次平行). 轮虫种类的鉴定参考文献[12,13,14,15,16].
浮游植物、 原生动物、 枝角类和桡足类的采集与鉴定参考文献[17,18,19,20,21,22].
水质测定:现场用YSI-Professional Plus多功能水质分析仪测量水温、 盐度; 同时取一定量的混合水样带回实验室当天测定水化指标:总氮(TN)采用碱性过硫酸钾消解紫外分光光度法测定、 总磷(TP)采用钼锑抗分光光度法测定、 叶绿素a采用丙酮提取分光光度法测定[23]. 1.3 数据分析
运用优势度(Y)、 丰富度指数Margalef指数(D)、 多样性指数Shannon-Wiener指数(H′)、 物种均匀度指数Pielou指数(J)、 杰卡特(Jaccad)群落相似性系数(CJ)对轮虫群落结构进行分析,分别按下列公式计算[24,25].
优势度:
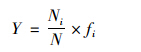
丰富度:
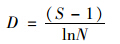
Shannon-Wiener多样度:
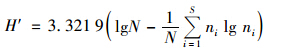
均匀度:
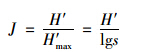
杰卡特(Jaccad)群落相似性系数:
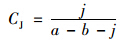
采用Origin 8.0软件进行作图. 应用SPSS 18.0软件对轮虫的密度与环境因子等水质数据进行相关性分析,P<0.05表示显著相关,P<0.01表示极显著性相关. 2 结果与分析 2.1 环境因子的变化 2.1.1 非生物环境因子
自2011年12月-2012年4月,总磷浓度变幅较小,在0.21-0.29 mg ·L-1之间变化; 总氮在试验初期变化剧烈,自5月开始,变化较为平缓,0.7-4.24 mg ·L-1. 2011年12月水温为10.4℃,自2012年1-8月,水温从6.1℃上升至33.2℃,而后下降至18.7℃. 盐度为0.87-2.73 (见图 2).
 | 图 2 临港B港河道非生物环境因子的变化Fig. 2 Monthly variation of abiotic environmental factors in the Lin gang B Gang River |
2011年12月临港B港河道的叶绿素a的含量最高,达40.96 μg ·L-1,1月较低. 原生动物纤毛虫在2012年3月达到高峰,密度为9300 ind ·L-1,纤毛虫开始以急游虫(Strombidium spp.)占优势,随着温度的升高,砂壳纤毛虫(Tintinnid)所占的比重越来越大(密度最大值为5875 ind ·L-1),主要是筒壳虫(Tintinnus spp.). 枝角类密度在2012年5月出现峰值,为5 ind ·L-1. 桡足类动物的密度在2012年1月达到最大值,其密度为24 ind ·L-1,桡足类以无节幼体和桡足幼体为主,成熟的个体多为剑水蚤(Cyclopoidea),哲水蚤只在部分月份出现且不会大量出现; 剑水蚤密度2011年12月出现最大值,其值为1 ind ·L-1(图 3).
 | 图 3 临港B港河道生物环境因子的变化Fig. 3 Monthly variation of biological environmental factors in the Lin gang B Gang River |
在调查期间,临港B港河道共发现6科15属23种轮虫,污染指示种19种. 寡污性种类5种,占总污染指示种的26.3%; 寡污-β中污性4种,占总污染指示种的21.1%; β-中污性9种占总污染指示种的47.4%,β-α中污性1种占总污染指示种的5.3%(表 1).
| 表 1 临港B港河道轮虫种类组成及其分布 1) Table 1 Species of rotifer occurring in different sampling stations of Lin gang B Gang River |
临港B港河道与芦潮引河轮虫的种类数与现存量变化见图 4:临港B港河道轮虫的种类数在2-13种之间变化,轮虫种类数总体处于上升状态,芦潮引河的轮虫种类数在0-9种之间变化,临港B港河道轮虫种类数除2012年2月外均高于芦潮引河. 临港B港河道和芦潮引河轮虫密度的变化均有一个主高峰和两个次高峰,临港B港河道分别是3月(3637.5 ind ·L-1)、 6月(2837.5 ind ·L-1)和9月(2516.7 ind ·L-1),最低值是12月仅有100 ind ·L-1; 芦潮引河轮虫密度主高峰出现在6月密度为2800 ind ·L-1,第一个次高峰是1月(2000 ind ·L-1),第二个次高峰在8月(2000 ind ·L-1). 临港B港河道的轮虫生物量在0.01-3.21 mg ·L-1之间变化,有一个明显的峰值在9月出现; 芦潮引河的轮虫生物量的范围为0-1.96 mg ·L-1,最高值也是在9月出现,11月盐度异常,未检出轮虫.
 | 图 4 临港B港河道和芦潮引河轮虫种类和现存量(密度、 生物量)月变化比较Fig. 4 Monthly variation of species,density and biomass of rotifer in the Lin gang B Gang River and Luchaoyin River |
主要优势种及其数量动态见图 5. 螺形龟甲轮虫是2012年3月和4月的单一优势种,但3月的密 度远远高于4月; 6月优势种为广布多肢轮虫、 裂痕龟纹轮虫和暗小异尾轮虫; 7月优势种比前月增加了角突臂尾轮虫; 8月优势种主要为广布多肢轮虫、 暗小异尾轮虫、 角突臂尾轮虫、 萼花臂尾轮虫、 叶状帆叶轮虫和前节晶囊轮虫; 9月优势种种类数量有所下降,主要是广布多肢轮虫、 暗小异尾轮虫、 前节晶囊轮虫和裂痕龟纹轮虫; 11月梳状疣毛轮虫和曲腿龟甲轮虫成为了新的优势种群.
 | 图 5 主要轮虫种类的密度变化Fig. 5 Monthly variation of density of main rotifer types |
在临港B港河道调查研究中,轮虫香农威尔指数(H′值)的范围为0.236-2.151; 轮虫均匀度指示(J值)在0.170-0.893之间变化; 轮虫丰富度指数(D值)的变化范围为1.443-4.678. (图 6). 临港B港的E/O值的变幅为0.75-4(表 2).
 | 图 6 临港B港河道轮虫多样性指数Fig. 6 Shannon-Wiener,species richness and evenness indexes of rotifer in the Lin gang B Gang river |
| 表 2 每个月的E/O值Table 2 E/O value of each month |
计算临港B港河道每月轮虫的群落相似性,除4月与5月、 7月与8月外其他各月间的群落相似性皆<0.5(表 3),表明临港B港河道各月间轮虫的群落结构有很大的差异. 轮虫群落相似性,从另外一方面也反映了临港B港河道各月间的生境不同.
| 表 3 各月之间的相似性系数Table 3 Similarity coefficient between months |
轮虫的密度及主要种类的轮虫密度与环境因子作Pearson相关分析,结果表明:轮虫总密度与温度、 叶绿素a呈显著正相关; 暗小异尾轮虫与温度、 叶绿素a呈极显著正相关,与盐度、 总氮、 总磷呈极显著负相关; 广布多肢轮虫与温度、 叶绿素a呈极显著正相关,与盐度、 总氮呈极显著负相关; 角突臂尾轮虫与温度呈极显著正相关,与盐度、 总氮呈极显著负相关; 螺形龟甲轮虫与盐度、 与总氮呈极显著正相关(表 4).
| 表 4 轮虫密度与环境因子的相关分析1) Table 4 Regression analysis of rotifer in the Lin gang B Gang River with environmental factors |
温度是影响浮游动物生长发育的一个重要环境因子[26],随着温度的上升,轮虫卵的发育时间缩短,在丰富的食物供给条件下,使种群周转加快,导致种群密度迅速增加,因此轮虫总数的高峰一般出现在20℃以上[27]. 临港B港河道的温度在3月时开始回升,广温性种类螺形龟甲轮虫大量繁殖,轮虫密度增大; 萼花臂尾轮虫在温度高于20℃时才大量繁殖,7月叶绿素a的浓度开始升高,为萼花臂尾轮虫提供了适宜的生长繁殖条件,使之在8月时成为优势种; 7、 8月水温高于33℃,较高的温度反而抑制了轮虫的生长[28]; 9月温度下降,轮虫密度随着温度降低而回升.
营养盐浓度的变化也能够通过对浮游植物的“上行作用”间接影响轮虫密度. 但在本研究中,轮虫的总密度与营养盐浓度没有显著性关系,这可能是因为临港B港河道属于新开挖的河道,其生态系统还未稳定,营养盐的变化较大,研究后期因为种植水草等修复工程营养盐的浓度降低,而温度、 叶绿素对轮虫密度升高的作用抵消了营养盐对轮虫的影响.
轮虫多以藻类和有机碎屑为食,但是不同种类的轮虫其自身的牧食特性不同[29],食物利用性的竞争会引起优势种群的变化. 3月和4月占优势的螺形龟甲轮虫属于小型滤食性轮虫,能够滤食一些体积较小的食物(个体大小在1-12 μm之间)[30]; 本研究中,银灰平裂藻等小型藻类在3月和4月时数量较高,为螺形龟甲轮虫提供了较丰富的食物来源,而随着浮游藻类的优势种向个体较大的种类转变,同样以藻类为食物的砂壳纤毛虫数量增多,螺形龟甲轮虫的食物受到限制,因此其密度从5月开始降低; 广布多肢轮虫个体较螺形龟甲轮虫大,能够牧食藻类和碎屑的范围较后者宽,且其适应性较强,在变化较为剧烈的6月大量出现成为优势种. 轮虫内部的一些杂食性(肉食性)轮虫可以通过直接捕食小型、 柔软的轮虫[31],而占据大部分资源. 个体较大的前节晶囊轮虫的繁殖力高峰在春夏之交,在8月大量出现,能够摄食一些小型的轮虫[32,33,34,35],例如螺形龟甲轮虫、 裂痕龟纹轮虫等,因而在其大量出现时裂痕龟纹轮虫等小型轮虫的密度急剧下降.
轮虫总密度的改变与“下行效应”也有一定的关系[36],在冬季桡足类的密度和其它一些捕食者能够限制轮虫的密度,同时轮虫本身在低温时生长繁殖速率较小也限制了其大量出现. 本研究中,5月时,枝角类的密度较高,增大了与轮虫间食物竞争压力,轮虫的密度不是很高. 自6月起,桡足类、 纤毛虫和枝角类密度较小,变化也比较平稳,剑水蚤桡足类的摄食压力、 与纤毛虫、 枝角类等的竞争压力均减小,轮虫的密度上升,形成了峰值. 3.2 盐度对轮虫群落的影响
河口区是不同水文性质的水流交汇处,其环境既不同于海水,又不同于淡水,水中营养物质丰富,为原生动物、 轮虫等小型浮游动物提供了重要的食物来源,曾有研究指出[37],河口浮游生物种类数随着盐度的降低,逐渐向江河方向减少,反之,愈近海洋,盐度愈高,种类也愈多. 亦有研究[38]表明在淡水和盐度较低的水体中轮虫的物种丰富度没有太大的不同,但在低盐度水体中轮虫的丰富度与盐度有显著性正相关关系. 与2012年3-6月在杭州湾调查结果相比,轮虫种类数:芦潮引河<临港B港河道<杭州湾,随着盐度的增加,轮虫的种类数增多; 4月和6月:杭州湾的盐度超过10,轮虫种类数:临港B港河道>芦潮引河>杭州湾,较高的盐度限制了轮虫的种类数及其密度; 5月:临港B港河道>杭州湾>芦潮引河(表 5). 与南汇东滩湿地围垦水域内的浮游动物群落结构变化与外围水体的对比结果相似[39]. 申屠青春等[40]在实验中发现当盐度在6.76时,轮虫的生物量和种类数会有明显下降的趋势. 在本研究中,临港B港河道底质为河口海滩滩涂,其水体盐度为0-3,河道盐度较低,轮虫的密度没有明显变化; 但一些主要的淡水轮虫类群如暗小异尾轮虫、 广布多肢轮虫、 角突臂尾轮虫等的数量与盐度表现出显著负相关关系,而螺形龟甲轮虫与盐度呈显著正相关,随着时间的推移盐度降低,并维持在一个相对稳定的较低水平. 在临港B港水体的盐度条件下,轮虫的群落结构受盐度的影响较小.
| 表 5 临港B港河道、 芦潮引河和杭州湾轮虫数量及现存量之间的对比Table 5 Different of Species,density and biomass of rotifer among Lin gang B Gang river,Luchaoyin river and Hangzhou bay |
一般认为,湖泊处于中等营养水平时生物群落的结构复杂,物种多样性高; 而在极端的贫营养或者超富营养条件下群落结构简单,物种多样性低[41]. 也就是说,在一定范围内,水体富营养化使物种多样性下降[42]. 在本研究中临港B港河道的轮虫物种多样性指数H′和丰富度指数D虽有波动,但总体上是上升的,这表明临港B港河道的水质状况有所好转. 而对于临港B港河道水质状况有所好转的原因,可能与2011-2012年河道陆续进行了人工修复等措施有关. 与物种多样性相关的轮虫种类数在临港B港河道逐月上升,这表明临港B港河道的富营养化程度有所降低,钱方平等[42]和熊金林等[43]研究发现轮虫种类数与综合状态指数呈显著的负相关关系也证明了这一点. 在调查初期,临港B港河道未发现寡营养型种类的轮虫,随着时间的推移,寡营养型种类的轮虫开始出现并逐渐增多,应用浮游动物中富营养型种(E)与寡营养型种(O)的种数比(E/O值)来评价水体营养类型,该比值在0.5以下为贫营养,0.5~1.0之间为中营养,0.5以上为富营养[44],9月以前的E/O值均大于1,显示临港B港河道处于富营养状态,9月开始E/O值小于1,表明临港B港河道的营养状态好转. 综合生物指数的分析认为临港B港河道的水环境有所好转.
但是,根据临港B港河道水质因子的变化不能够证明水体状况的改善,生物指数的判断与理化因子的判断出现了矛盾,即生态位理论不能够全面地解释临港B港河道轮虫群落结构变化的规律. 生态位理论能够很好地解释轮虫密度的变化,关于物种多样性等解释得不是很好,生态位忽视的某些方面,正是中性理论所包含的东西. 中性理论能够通过物种扩散解释物种多样性和物种相对多度[45,46],临港B港河道轮虫种类数随着时间的延长而增加,多样性指数逐月上升,这些都可以证明物种扩散在水体轮虫群落结构的演替中发挥着重要的作用. 因而,应用生态位理论与中性理论相结合能够更好地解释临港B港河道轮虫群落的次生演替规律. 4 结论
(1)临港B港河道作为一个新生的河口区河道,在建成1 a中,轮虫种类数整体呈增长趋势,由单一的螺形龟甲轮虫占优势向多种轮虫共同占优势的状态转变,轮虫的群落结构主要受温度和食物(藻类等)的影响.
(2)各月间的群落相似性指数低,这表明临港B港河道的轮虫群落结构还不稳定,处于变化的过程中. 关于临港B港河道何时能够稳定,还有待更长时期的进一步研究.
(3)轮虫的多样性指数及污染指示种显示临港B港河道的水体正在明显好转,可达到轻污染、 中营养状态.
(4)在河口区盐度为0-3的情况下,轮虫总密度受盐度影响不大,但是优势种随盐度变化有一定的影响.
| [1] | 郭凯, 赵文, 殷守仁, 等. 北京官厅水库轮虫群落结构与水体富营养化状况[J]. 湖泊科学, 2010, 22 (2): 256-264. |
| [2] | Chen L J, Liu Q, Peng Z R, et al. Rotifer community structure and assessment of water quality in Yangcheng Lake[J]. Chinese Journal of Oceanology and Limnology, 2012, 30 (1): 47-58. |
| [3] | 周淑婵, 黄祥飞, 唐涛, 等. 香溪河库湾轮虫现状及水质评价初探[J]. 水生生物学报, 2006, 30 (1): 52-57. |
| [4] | Sládeček V. Rotifers as indicators of water quality[J]. Hydrobiologia, 1983, 100 (1): 169-201. |
| [5] | Angeler D G, Alvarez-Cobelas M, Sánchez-Carrillo S. Evaluating environmental conditions of a temporary pond complex using rotifer emergence from dry soils[J]. Ecological Indicators, 2010, 10 (2): 545-549. |
| [6] | Devetter M. Influence of environmental factors on the rotifer assemblages in an artificial lake[J]. Hydrobiologia, 1998, 387-388: 171-178. |
| [7] | Devetter M. Seasonal development of planktonic rotifers in Slapy Reservoir (Czech Republic)[J]. Biologia, 2011, 66 (4): 662-668. |
| [8] | Basińska A, Natalia Kuczyńska-Kippen N, Swidnicki K. The body size distribution of Filinia longiseta (Ehrenberg) in different types of small water bodies in the Wielkoposka region[J]. Limnetica, 2010, 29 (1): 171-182. |
| [9] | Barbieroa R P, Warren G J. Rotifer communities in the Laurentian Great Lakes, 1983-2006 and factors affecting their composition[J]. Journal of Great Lakes Research, 2011, 37 (3): 528-540. |
| [10] | 陈立婧, 顾静, 彭自然, 等. 上海崇明岛明珠湖轮虫群落结构[J]. 应用生态学报, 2009, 20 (12): 3057-3062. |
| [11] | 杨桂军, 秦伯强 高光, 等. 太湖不同湖区轮虫群落结构季节变化的比较研究[J]. 环境科学, 2008, 29 (10): 2963-2969. |
| [12] | 韩茂森, 束蕴芳. 中国淡水生物图谱[M]. 北京: 海洋出版社, 1995. |
| [13] | 王家楫. 中国淡水轮虫志[M]. 北京: 科学出版社, 1961. |
| [14] | 诸葛燕. 中国典型地带轮虫的研究[D]. 武汉: 中国科学院水生生物研究所, 1997. |
| [15] | Walter Koste. ROTATORIA: Die Rädertiere Mitteleuropas. 2 vols[M]. Gebrüder Borntraeger. Berlin. Stuttgart. West Germany, 1978. |
| [16] | Wallace R L, Snell T W, Ricci C. Rotifera. 1. Biology, ecology and systematics[M]. Ghent: Kenobi Productions, 2006. |
| [17] | 胡鸿钧, 李尧英, 魏印心, 等. 中国淡水藻类[M]. 上海: 上海科学技术出版社, 1979. |
| [18] | 中国原生动物学会. 原生动物[M]. 北京: 科学出版社, 1999. |
| [19] | 张武昌, 丰美萍, 于莹, 等. 砂壳纤毛虫图谱[M]. 北京: 科学出版社, 2012. |
| [20] | 中国科学院动物研究所甲壳研究组. 中国动物志 ·节肢动物门 ·甲壳纲 ·淡水桡足类[M]. 北京: 科学出版社, 1979. |
| [21] | 张武昌, 赵楠, 陶振铖, 等. 中国海浮游桡足类图谱[M]. 北京: 科学出版社, 2010. |
| [22] | 蒋燮治, 堵南山. 中国动物志 ·节肢动物门 ·甲壳纲 ·淡水枝角类[M]. 北京: 科学出版社, 1979. |
| [23] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [24] | 徐兆礼. 长江口北支水域浮游动物的研究[J]. 应用生态学报, 2005, 16 (7): 1341-1345. |
| [25] | 郑金秀, 胡菊香, 彭建华, 等. 长江口南北支浮游动物群落生态学研究[J]. 生态环境学报, 2011, 20 (6-7): 1102-1106. |
| [26] | Marques S C, Azeiteiro U M, Marques J C, et al. Zooplankton and ichthyoplankton communities in a temperate estuary: spatial and temporal patterns[J]. Journal of Plankton Research, 2006, 28 (3): 297-312. |
| [27] | 章宗涉, 黄祥飞. 淡水浮游生物的研究方法[M]. 北京: 科学出版社, 1991. |
| [28] | 王丽卿, 程婧蕾, 郑小燕, 等. 淀山湖轮虫多样性及生物量时空变化[J]. 水产学报, 2008, 32 (6): 906-914. |
| [29] | Holst H, Zimmermann H, Kausch H, et al. Temporal and spatial dynamics of planktonic rotifers in the Elbe Estury during Spring[J]. Estuarine Costal and Shelf Science, 1998, 47 (3): 26l-273. |
| [30] | Bogdan K G, Gilbert J J. Quantitative comparison of food niches in some freshwater zooplankton[J]. Oceologia, 1987, 72 (3): 331-340. |
| [31] | Bertani I, Leonardi S, Rossetti G. Antipredator-induced trait changes in Brachionus and prey selectivity by Asplanchna in a large river under low-discharge conditions: evidence from a field study[J]. Hydrobiologia, 2013, 702 (1): 227-239. |
| [32] | Gilbert J J. The cost of predator-induced morphological defense in rotifers: experimental studies and synthesis[J]. Journal of Plankton Research, 2013, 35 (3): 461-472. |
| [33] | García C E, Nandini S, Sarma S S S. Demographic characteristics of the copepod Acanthocyclops americanus (Sars, 1863) (Copepoda: Cyclopoida) fed mixed algal (Scenedesmus acutus)-rotifer (Brachionus havanaensis) diet[J]. Hydrobiologia, 2011, 666 (1): 59-69. |
| [34] | Guo R X, Snell T W, Yang J X. Ecological strategy of rotifer (Brachionus calyciflorus) exposed to predator-and competitor-conditioned media[J]. Hydrobiologia, 2011, 658 (1): 163-171. |
| [35] | Oganjan K, Virro T, Lauringson V. Food spectrum of the omnivorous rotifer Asplanchna priodonta in two large northeastern European lakes of different trophy[J]. Oceanological and Hydrobiological Studies, 2013, 42 (3): 314-323. |
| [36] | Yoshida T, Urabe J, Elser J J. Assessment of ‘top-down' and ‘bottom-up' forces as determinants of rotifer distribution among lakes in Ontario, Canada[J]. Ecological Research, 2003, 18 (6): 639-650. |
| [37] | 郑重. 海洋浮游生物生态学文集[M]. 厦门: 厦门大学出版社, 1986. |
| [38] | Kaya M, Fontaneto D, Segers H, et al. Temperature and salinity as interacting drivers of species richness of plantktonic rotifers in Turkish continental waters[J]. Journal of Limnology, 2010, 69 (2): 297-304. |
| [39] | 李强, 马长安, 吕巍巍, 等. 南汇东滩湿地围垦水域内浮游动物群落结构的变化[J]. 应用生态学报, 2012, 23 (8): 2287-2294. |
| [40] | 申屠青春, 董双林, 赵文, 等. 盐度、碱度对浮游生物和水化因子的影响[J]. 应用生态学报, 2000, 11 (3): 449-454. |
| [41] | 生态学丛书编委会. 环境和指示生物 ·水域分册[M]. 北京: 中国环境科学出版社, 1987. |
| [42] | 钱方平, 席贻龙, 温新利, 等. 湖泊富营养化对轮虫群落结构及物种多样性的影响[J]. 生物多样性, 2007, 15 (4): 344-355. |
| [43] | 熊金林, 梅兴国, 胡传林, 等. 湖北四湖泊营养类型与轮虫群落的关系[J]. 动物学杂志, 2003, 38 (6): 8-14. |
| [44] | 张觉民, 何志辉. 内陆水域渔业自然资源调查手册[M]. 北京: 农业出版社, 1991. 12-109. |
| [45] | 牛克昌, 刘怿宁, 沈泽昊, 等. 群落构建的中性理论和生态位理论[J]. 生物多样性, 2009, 17 (6): 579-593. |
| [46] | 张立敏, 陈斌, 李正跃. 应用中性理论分析局域群落中的物种多样性及稳定性[J]. 生态学报, 2010, 30 (6): 1556-1563. |