 2014, Vol.35
2014, Vol.35 



2. 中国科学院大学, 北京 100049;
3. 宝鸡文理学院陕西省灾害监测与机理模拟重点实验室, 宝鸡 721013;
4. 鲁东大学地理与规划学院, 烟台 264025
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Key Laboratory of Disaster Monitoring and Mechanism Simulating of Shaanxi Province, Baoji University of Arts and Sciences, Baoji 721013, China;
4. Institute of Geography and Plan, Ludong University, Yantai 264025, China
N2 O是仅次于CO2和CH4的重要温室气体,增温效应显著,其在过去100 a中对全球温室效应的贡献达4%~7%[1,2]. 滨海湿地作为陆地和海洋过渡区的重要组成部分,是海岸带一个非常重要的生态类型. 硝化-反硝化作用是滨海湿地土壤氮循环的重要过程,其对土壤N2 O的产生具有重要影响[3]. 目前,大多数研究认为N2 O的生成主要来自生物过程,如Bouwman等[2]指出,土壤微生物主导的硝化-反硝化作用所产生的N2 O约占全球大气中N2 O总量的90%. 但也有研究表明,土壤中的N2 O可能来自非生物作用,如铁的还原作用[4,5]. 因此,揭示滨海湿地土壤N2 O产生的生物与非生物机制及贡献对于准确估算土壤N2 O的释放量具有重要意义. 当前,国外已对滨海湿地N2 O排放过程、 生成机制及影响因素进行了较多研究,且其研究对象已涉及海湾湿地或盐沼[6]、 河口湿地或海岸湿地[7]以及南极滨岸湿地[8]. 与之相比,国内关于滨海湿地N2 O排放过程的研究还不多见,且主要围绕长江口和珠江口潮滩湿地[9,10]、 南极海岸湿地[11]开展了一些相关研究. 黄河口滨海湿地是河口与渤海相互作用形成的重要湿地类型. 当前,关于黄河口滨海湿地N2 O排放特征及影响因素已开展了较多研究,并取得许多重要成果[12,13],但关于湿地土壤N2 O生成机制的研究还鲜有报道.
20世纪80年代以来,黄河流域水量的年际变化较大. 根据利津水文站1980~2007年实测资料统计,黄河流域利津水文站最大年径流量为491×108 m3 (1983年),此后年径流量一直下降到200×108 m3左右,由此导致河水漫滩几率减少,侧渗补给到黄河口两岸湿地的水量也呈明显减少趋势,使得保护区内部分湿地严重退化. 为此黄河水利委员会自1999年开始对黄河水量实行统一调度,以引黄补水为主要手段对退化湿地进行恢复. 已有研究表明,生态恢复工程使退化湿地土壤氮素和有机质含量、 pH值、 质地和氧化还原电位等发生很大变化[14,15]. 特别是水文情势的改变可显著影响土壤硝化和反硝化过程,进而对土壤N2 O释放量产生重要影响. 目前,关于黄河口生态恢复工程对湿地土壤N2 O生成机制的影响研究还未见报道. 鉴于此,本研究应用Wrage等[16]于2004年报道的方法,通过室内控制实验对不同恢复阶段湿地土壤的N2 O生成机制及贡献进行了研究,详细区分了土壤硝化作用、 反硝化作用、 硝化细菌反硝化作用以及非生物作用N2 O产生过程,分析了生态工程实施前后水文情势变化以及温度水分对N2 O排放过程的影响,结果将有助于评估黄河口生态恢复工程对湿地温室气体源/汇功能的影响,旨在为下一步生态工程调控湿地土壤N2 O排放和氮转化过程提供科学依据. 1 材料与方法 1.1 研究区域概况
研究区位于山东省黄河三角洲国家级自然保护区内(37°40′ N~38°10′ N,118°41′ E~119°16′ E). 保护区属暖温带季风气候区,具有明显的大陆性季风气候特点,雨热同期,四季分明,冷热干湿界限极为明显. 该区年平均气温12.1℃,无霜期196 d,≥10℃的年有效积温约4 300℃,年均蒸发1 962 mm,年均降水量为551.6 mm,70% 的降水集中于7、 8月. 区域内地势十分平坦(平均坡降为1/8 000~1/12 000),生态格局时空变化迥异,湿地类型多样,植被以草甸为主.
1.2 研究方法 1.2.1 样品采集与处理实验区位于山东省黄河三角洲自然保护区大汶流管理站和黄河口管理站所辖区域内(图 1). 按照“典型性、 代表性、 一致性”的原则,设置未恢复区(取样前一直处于退化状态,R0)、 2002年恢复区(从2002年开始生态恢复,采样时已恢复10 a,R2002)和2007年恢复区(从2007年开始生态恢复,采样时已恢复5 a,R2007)这3个典型芦苇湿地样地. 在各采样区内采集3个典型土壤剖面,采样深度为0~20 cm,采样间隔为10 cm. 然后对3个剖面样品进行等层次混合,共采集12个土壤样品. 将采集的土样带回实验室,自然风干后拣去石块、 植物残根等杂物,用球磨机磨碎,分成2份:一份用于土壤理化性质测定; 另一份用于培养实验. 土壤pH值采用酸度计(PHS-3D) 测定(水土比2.5 ∶1),电导率(水溶性盐总量EC) 采用电导法(DDS-307电导率仪) 测定(水土比5 ∶1),TC和TN采用元素分析仪测定,NH+4-N和NO-3-N用2 mol ·L-1的KCL浸提后通过连续流动分析仪(Bran-Lubee AAA3)测定,有机碳采用高温外热重铬酸钾容量法测定,土壤粒度采用激光粒度仪测定,按国际制分类.
 | 图 1 黄河三角洲自然保护区位置及恢复区样点布设
Fig. 1 Location of the Yellow River natural reserve and experimental sites of wetland restoration
|
称取10 g过2 mm孔筛的3种供试土样(每种供试土样分0~10 cm和10~20 cm两个土层),分别放入300 mL血清瓶中. 加适量去离子水调整到80%田间持水量(WHC),置于25℃的培养箱中预培养1 d后,按照表 1所示添加抑制剂后继续在25℃恒温培养箱内培养24 h. 分别在添加抑制剂后培养的第0、 6和24 h抽取定量气体,立刻用Agilent7890气相色谱仪测定. 实验设3个重复,共72份. 相关研究表明,当土壤水分含量为最大持水量的50%~70%时最有利于硝化微生物的活动,即硝化作用最旺盛[17]; 当水分增加促使形成厌氧条件时,硝化速率呈明显下降趋势[18],而某些嫌气性和兼气性微生物开始活跃,反硝化作用增强. 另外当土壤水分含量低于田间持水量的60%时,反硝化作用可忽略不计[19]. 故设置两个水分梯度(60% WHC,80% WHC),方法同上,称取3种供试土样表层样品(0~10 cm),25℃恒温培养(设3个重复,共72份),以研究水分对N2 O生成过程的影响. N2 O的排放通量与温度呈正相关性,通常在25~35℃范围内达到N2 O的最大排放通量[20]. 张树兰等[21]的研究表明,30℃时硝化作用最大,20℃则对硝化作用产 生一定的抑制. 而反硝化作用可在一个较宽的温度范围内(2~65℃)进行,最适宜温度为25℃左右. 故设置3个温度梯度(20、 25、 30℃),方法同上,调整水分含量为80% WHC(设3个重复,共108份),以研究温度对土壤N2 O生成过程的影响. 方法同上,设置20℃和80% WHC、 25℃和80% WHC、 30℃和80% WHC、 20℃和60% WHC、 25℃和60% WHC、 30℃和60% WHC这6个水分与温度交互处理(设3个重复,共216份),以研究温度和水分交互作用对土壤N2 O生成过程的影响.
| 表 1 抑制剂及其对N2 O产生过程影响 1) Table 1 Inhibitors and effects on soil processes generating N2 O |
(1)不同过程N2 O产生量采用下式计算:

(2) 利用添加抑制剂后0、 6和24 h测定的N2 O排放速率(μL·L-1),应用Slope函数,求得N2 O浓度随时间变化的回归曲线斜率(dc/dt),选择|R|>0.9 数值,然后按下式计算N2 O产生率.
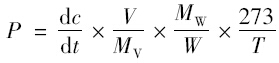
运用Origin 8.0和SPSS 16.0软件对数据进行作图、 计算和相关分析. 2 结果与分析 2.1 湿地土壤N2 O 生成过程及其贡献
图 2为不同恢复阶段(R0、 R2007和R2002)湿地土壤表层和亚表层不同过程的N2 O产生量及其贡献. 就表层土壤(0~10 cm)而言,R0 的N2 O总产生量最低,且主要来自非生物作用的贡献(1 194.56%). 硝化作用和硝化细菌反硝化作用的贡献率较低(仅有44.31%和26.67%),而反硝化作用对其产生削弱作用(-1 165.53%). R2002 的N2 O产生量主要来自硝化细菌反硝化作用和非生物作用的贡献(分别为53.89%和229.36%). 硝化作用的贡献较低(仅有16.68%),而反硝化作用对其产生削弱作用(-199.93%). R2007 的N2 O产生量最大,并以硝化作用和非生物作用的贡献较高(分别为47.69%和93.38%),而反硝化作用则对其产生削弱作用(-64.30%). R0、 R2007和R2002表层土壤仅非生物作用N2 O产生量的差异显著(P<0.05),而硝化作用、 反硝化作用和硝化细菌反硝化作用N2 O产生量的差异均不显著(P>0.05).
 | 图 2 不同恢复阶段湿地土壤不同过程N2 O产生量
Fig. 2 N2 O production from different processes in wetland soils of different restoration phases
Ni.硝化作用;De.反硝化作用;ND.硝化细菌反硝化作用;OS.非生物源;TP.总产生量;下同 |
就亚表层土壤(10~20 cm)而言,R0 的N2 O总产生量亦最低,但主要自硝化细菌反硝化和非生物作用的贡献(分别为606.18%和1532.08%),而硝化作用和反硝化作用对其产生削弱作用(分别为-614.15%和-1 424.10%). 对恢复湿地土壤而言,其N2 O总产生量大于未恢复湿地土壤,且差异达到极显著水平(P<0.01). 与表层土壤不同,R2002的 N2 O总产生量要远大于R2007,且硝化细菌反硝化作用和非生物作用对其贡献较高(分别为101.87%和96.92%),而硝化作用和反硝化作用对其产生削弱作用(分别为-12.25%和-86.54%). R2007的N2 O产生量主要来自硝化作用和非生物作用的贡献(分别为128.85%和494.95%),反硝化作用和硝化细菌反硝化作用则对其产生削弱作用(分别为86.54%和101.87%). R0、 R2007和R2002亚表层土壤硝化细菌反硝化作用和非生物作用N2 O的产生量均存在显著差异(P<0.01或P<0.05),而硝化作用和反硝化作用N2 O产生量的差异均不显著(P>0.05).
2.2 水分对湿地土壤N2 O生成过程的影响水分对不同恢复阶段(R0、 R2007和R2002)湿地表层土壤N2 O生成过程的影响不尽一致(图 3、 图 4). 随着水分的增加,R0的 N2 O总产生量反而降低,而R2007 和R2002 的N2 O总产生量分别增加12.32倍和7.24倍. 具体而言,水分对R0土壤 N2 O生成过程的影响较小. 当水分增加后,R0 的硝化细菌反硝化作用降低(降低0.88倍),非生物作用增强,但两个过程在不同水分处理间的差异均不显著(P>0.05). 与之相比,R2007和R2002土壤的N2 O生成过程受水分影响较大. 当水分增加后,二者反硝化作用过程的N2 O生成均受到明显抑制(分别降低1.21倍和0.37倍). R2007土壤非生物作用的N2 O产生量在80% WHC下较60% WHC 增加1.23倍,且其差异达到极显著水平(P<0.01). 相对于R2007,R2002土壤的硝化细菌反硝化作用和非生物作用的N2 O产生量增幅较小,仅增加1.61倍和0.54倍.
 | 图 3 温度和水分对湿地土壤N2 O总产生量的影响
Fig. 3 N2 O production in wetland soil due to the effects of temperature and water content
|
 | 图 4 温度和水分对湿地土壤N2 O生成过程的影响
Fig. 4 Effects of temperature and water content on wetland soil processes generating N2 O
|
温度对不同恢复阶段(R0、 R2007和R2002)湿地表层土壤N2 O排放过程的响程度差异较大(图 3、 4). R2002土壤的N2 O总产生量表现为25℃>30℃>20℃,且不同温度处理之间存在显著差异(P<0.05). R0土壤的N2 O总产生量整体表现为20℃>30℃>25℃,且不同温度处理之间亦存在显著差异(P<0.05). 与R2002 相似,R2007土壤的N2 O总产生量亦表现为25℃>30℃>20℃,但不同温度处理之间的差异未达到显著水平(P>0.05). 具体而言,R0土壤的诸N2 O生成过程受温度的影响程度不尽一致. 硝化作用的N2 O产生量在25℃条件下取得最大值,硝化细菌反硝化作用在20℃条件下取得最大值,而非生物作用在30℃条件下取得最大值. 与之相比,R2007土壤的诸N2 O生成过程受温度的影响程度较为一致. 25℃条件下,硝化作用、 硝化细菌反硝化作用、 非生物作用的N2 O产生量以及N2 O总产生量均取得最大值. 不同的是,R2002土壤硝化作用和非生物作用的N2 O产生量在30℃条件下取得最大值,而硝化细菌反硝化作用的N2 O产生量在25℃条件下取得最大值.
2.4 温度和水分交互作用对湿地土壤N2 O生成过程的影响温度和水分对不同恢复阶段(R0、 R2007和R2002)湿地表层土壤的N2 O总产生量存在不同程度的交互 影响(图 3). R0土壤的N2 O总产生量在20℃和80% WHC条件下取得最大值,而R2007和R2002土壤均在25℃ 和80% WHC条件下取得最大值. 方差分析表明,水分对R0、 R2007和R2002土壤的N2 O总产生量均存在显著影响(P<0.05),温度对R0 和R2002土壤的N2 O总产生量存在极显著影响(P<0.01),而温度和水分交互作用对R0 和R2002土壤的N2 O总产生量均存在极显著影响(P<0.01)(表 2). 温度和水分交互作用对不同恢复阶段(R0、 R2007和R2002)湿地表层土壤N2 O诸生成过程的影响程度差异较大(图 4). 就R0土壤而言,尽管20℃和60% WHC条件对反硝化作用和硝化细菌反硝化作用N2 O产生量的抑制作用最大,但却明显促进了非生物作用的N2 O产生量. 与之相比,R2007土壤的硝化作用、 硝化细菌反硝化作用和非生物作用的N2 O产生量均在25℃和80% WHC条件下取得最大值,而R2002土壤的硝化作用和非生物作用的N2 O产生量在30℃和80% WHC条件下取得最大值.
| 表 2 温度和水分交互作用下湿地土壤N2 O产生量方差分析 Table 2 Results of ANOVA for N2 O production in wetland soil under the interactive effects of temperature and water content conditions |
目前大多数研究认为,土壤N2 O的生成主要来自生物过程,如硝化作用、 反硝化作用和硝化细菌反硝化作用等. 本研究表明,R0表层土壤的硝化作用对N2 O产生虽有贡献,但贡献率较低; 而亚表层土壤硝化作用则对其存在较大的削弱作用. 原因在于,R0表层和亚表层土壤的水分条件较差,含盐量较高(表 3),而较高的盐分条件可对硝化作用产生一定的抑制作用[22,23]. 相关分析也表明,土壤硝化作用与土壤EC呈负相关,这与牟晓杰等[24]对闽江河口湿地的研究结果一致. 由于土壤硝态氮含量高低是表征土壤硝化作用强弱的重要参数,所以硝态氮含量越高其硝化潜力越强[24]. 相对于恢复区,R0表层土壤的硝态氮含量较高(表 3),由此使得其硝化作用可能对N2 O产生具有一定贡献. 已有研究表明,硝化细菌的最适pH为7.7~8.1,过高的pH会降低细菌活性,进而影响土壤硝化作用[25]. 本研究表明,不同恢复阶段土壤不同土层中,R2007 表层土壤硝化作用产生的N2 O量最大. 这与Strauss等[26]的研究结果一致,即当河流沉积物的硝化作用在pH为7.5~7.8时最强,但随pH的增加而降低. 生态恢复工程导致R2007表层土壤的pH值有所降低,仅为7.64(表 3),由此使得其硝化作用产生的N2 O增加. 此外,湿地水分状况可通过影响土壤通气状况和氧分压而对好气性硝化细菌活性产生重要影响[27]. Flowers等[28]指出,适宜水分条件可促进硝化作用的进行. 由于恢复区的水分条件适宜,为土壤硝化作用提供了良好反应条件. Rückauf等[29]的研究发现,淹水湿地土壤条件下,反硝化作用是主要过程,但N2的释放量超过N2 O而成为反硝化作用的主要产物. 但本研究得到不同结论,即不同恢复阶段湿地土壤的反硝化作用一直对N2 O产生削弱作用. 本研究中,反应物NO-3-N 可被该区丰富的活性铁还原为N2 O而利用[5,30],而这也是导致非生物作用对N2 O的产生具有很大贡献大的重要原因. 反硝化微生物利用有机碳作为能量的电子供体和细胞成分合成物质,因此反硝化作用在很大程度上依赖于土壤有机碳含量. 有研究指出,田间条件下土壤中易溶性有机碳有利于反硝化作用的进行[31]. 土壤中反硝化作用与土壤中有机碳的含量密切相关,有机碳含量不仅决定了土壤反硝化作用的大小而且影响着的土壤反硝化活性[32]. 本研究的黄河口区域属新生湿地,有机碳含量很低(表 3),这使得反硝化细菌能量可能供应不足,导致其活性降低,不利于反硝化作用进行. Webster等[33]的研究发现,硝化细菌反硝化作用能够产生可观的N2 O. Wrage等[34]则认为,硝化细菌反硝化作用是硝化作用的一个特殊过程,通过乙炔抑制法测得的硝化细菌反硝化作用对土壤N2 O生成的贡献为40%[16],与本研究中不同恢复阶段湿地表层土壤硝化细菌反硝化作用对N2 O生成的贡献相近. 恢复区土壤硝化细菌反硝化作用的N2 O产生量明显高于未恢复区(图 2),这可能与其NH+4-N含量比未恢复区高,更有利于硝化细菌反硝化作用的进行有关[4]. 相关分析表明,土壤硝化细菌反硝化作用与土壤NO-3-N含量呈显著负相关(P<0.05),与NH+4-N含量呈较大正相关(表 4),说明硝化细菌反硝化作用与NO-3-N含量、 NH+4-N含量密切相关. 可见,硝化作用和反硝化作用受土壤NH+4/NO-3含量、 有机碳、 pH、 含水量和盐分等多种因素制约. 由于生态恢复工程显著改变了湿地土壤的理化性质,进而对硝化-反硝化作用机制以及土壤的N2 O生成产生重要影响. 与生物作用相比,非生物作用下土壤N2 O的产生研究相对较少,但其对N2 O产生的贡献却不容忽视. 已有研究表明,土壤N2 O的产生可能来自非生物作用,如化学反硝化过程(NO-2自身的化学分解或NO-2与其他物质的化学反应),其产物主要为NOx、 N2 O和N[4,5]2. Wrage等[16]认为,无微生物参与的化学反硝化过程在特定情况下可产生少量的N2 O. 与生物过程相比,非生物过程具有反应速率快的特点. 在反应产物方面,某些化学还原剂可在一定条件下通过反硝化作用获得比生物方法更多的N2 O. 本研究表明,黄河口不同恢复阶段湿地土壤的非生物过程对N2 O产生量的贡献很高,这可能与研究区为高活性铁区有关[30]. 另外相关研究表明[30,35],黄河口地区铁含量较高,悬浮物中的铁高达4.92%,沉积物中铁含量均值为22.54 g ·kg-1. NO-3-N 在碱性条件下可被Fe(Ⅱ)还原,而NO-2在酸性和碱性条件下均可被Fe(Ⅱ)还原,主要还原产物为NH+4、 N2 O和N[5]2. 相关分析也表明,土壤非生物作用与pH呈极显著负相关(P<0.01),说明非生物作用适合在酸性环境中进行. 此外,也可能与乙炔抑制剂对生物作用抑制不完全,由此使得部分归于非生物作用贡献的N2 O实际上是由于生物作用而产生[16]. 本研究表明,R0土壤的非生物作用过程对N2 O产生的贡献要远远大于R2007和R2002,且其非生物作用的N2 O产生量也要大于R2007和R2002,这可能是由于非生物作用过程受粒度影响较大,R0 土壤以砂粒为主(表 3),且表层和亚表层土壤砂粒含量明显高于恢复区(R2002和R2007),适宜非生物过程进行有关. Wrage等[16]对草原土壤的相关研究亦得到类似结论,即砂质土壤的N2 O产生主要来自非生物作用过程的贡献. 综上,尽管黄河口不同恢复阶段湿地土壤N2 O的产生是生物作用与非生物作用共同作用的结果,但因非生物作用对N2 O产生的影响较大,应受到特别关注.
| 表 3 不同恢复阶段湿地土壤理化性质 Table 3 Physical and chemical properties of wetland soils in different restoration phases |
| 表 4 湿地土壤N2 O产生过程与土壤理化性质的相关系数 1) Table 4 Correlation coefficients between wetland soil processes generating N2 O and soil physical and chemical properties |
温度是影响土壤中微生物活动的重要因素之一[36],其主要是通过影响硝化细菌和反硝化细菌活性而影响 N2 O的总产生量. 本研究表明,R2002和R2007 土壤的N2 O总产生量均在25℃取得最大值,R0则在20℃取得极大值,说明温度对不同恢复阶段土壤N2 O生成过程的响程度差异较大. 这主要与不同温度条件下土壤微生物的活性差异有关,生态工程实施以后恢复区土壤一直处于滞水状态,使得恢复区土壤温度高于未恢复区,恢复区土壤微生物最适温度略高于未恢复区. 已有研究表明,不同温度条件下土壤有机质分解和氮矿化过程差异明显,由此导致其N2 O产生量差异较大[20]. Breuer等[37]的研究发现,土壤硝化作用强度与温度存在显著正相关. 土壤温度每升高1℃,硝化速率增高1.17 mg ·(m2 ·h)-1. Stottmeister等[38]发现,当温度介于15~20℃时,净硝化速率和总硝化速率随温度的上升而下降. Malhi等[39]对加拿大西部阿尔伯达一带3种土壤的研究显示,硝化作用的最适温度在20℃,当温度达到30℃时,硝化作用完全停止. 本研究表明,R0和R2007的硝化作用N2 O产生量在25℃取得最大值,而R2002在30℃取得极大值,这可能与不同恢复阶段湿地土壤中硝化细菌对温度的响应差异有关. 比较而言,反硝化作用可在一个较宽的温度范围内(2~65℃)进行,最适宜温度为25℃左右. 在该温度范围内,反硝化速率随温度的升高而增强,但温度过高或过低均不利于反硝化作用的进行[17]. 这与本研究关于恢复区土壤的相关结果一致,即R2002和R2007 土壤的硝化细菌反硝化作用均在25℃取得最大值,而在20℃和30℃时,硝化细菌反硝化作用处于明显抑制状态.
土壤水分是影响气体扩散率和微生物活动的关键因素[22]. 本研究表明,随着水分的增加,R0表层土壤的N2 O总产生量反而降低,而R2007和R2002表层土壤的N2 O总产生量则分别增加12.32倍和7.24倍,说明水分对不同恢复阶段湿地土壤N2 O产生过程的影响不尽一致,这主要与不同水分条件下土壤微生物的活性差异有关,生态工程实施以后恢复区土壤一直水源充足,使得其土壤微生物适宜在水分充足的环境中生存,而未恢复水分条件一直较差,其
微生物活性在水分含量较低时反而活跃. 本研究中,恢复区与未恢复区土壤的N2 O总产生量均在80% WHC时最高,说明土壤的微生物活性在此水分条件较强,有助于N2 O的产生. Malhi等[39]的研究表明,土壤硝化率在水势为-1 500 kPa时最低,为-33 kPa时最低. 在一定含水量范围内,土壤硝化速率随含水量的增加而增大,硝化势在田间持水量的60%时达到最大值[28]. 本研究得到不同研究结果,即恢复区与未恢复区土壤的硝化作用均在80% WHC时对N2 O总产生量有贡献,而在60% WHC时具有削弱作用. 这可能与黄河口地区硝化细菌对水分的响应程度密切相关. 由于反硝化作用是由某些嫌气性和兼气性微生物完成的,故反硝化速率与土壤中的氧气浓度呈负相关. 在较高含水量时,N2 O主要来自反硝化作用的贡献,而在较低含水量时,N2 O主要来自硝化作用贡献[40]. 这与本文关于恢复区土壤的研究结果类似,即硝化细菌反硝化作用在80% WHC时的N2 O产生量明显大于60% WHC,且R2002在80% WHC时硝化细菌反硝化作用的N2 O产生量大于硝化作用的产生量.
温度和水分交互实验表明,不同恢复阶段土壤的N2 O总产生量均在80%WHC时取得最大值. 比较而言,温度变化的影响相对较弱,而水分含量对土壤N2 O产生过程的影响较为明显(图 4). 这与付素静[17]对干旱荒漠区典型土壤硝化-反硝化作用的研究结果一致,其原因主要有两方面:①反硝化作用可在一个较宽的温度范围内(2~65℃)进行,温度对反硝化作用的影响相对较小; ②水分含量对硝
化-反硝化作用的影响较大. 硝化细菌是好氧微生物,因此受土壤中氧分压的剧烈影响. 水分含量的变化通常会对土壤通气状况以及氧分压造成影响,进而影响硝化作用的进行. 而反硝化作用是由某些嫌气性和兼气性的微生物完成的,水分含量是影响氧气和其他气体有效性、 扩散率以及微生物活动的关键因素,进而可影响反硝化作用的进行[18].
3.3 湿地土壤N2 O产生量与相关研究结果对比本研究中,不同恢复阶段湿地土壤各土层的N2 O总产生量介于(0.37±0.08)~(9.75±7.64)nmol ·(kg ·h)-1,总体均表现释放N2 O. 恢复区土壤的N2 O产生量要大于未恢复区土壤. 目前国外对退化滨海湿地恢复已涉及景观恢复及其引起的温室气体源/汇功能评估等方面[41,42]. 与之相比,国内对该领域研究大多集中于景观恢复方面[14,15],而对于温室气体源/汇功能评估研究甚少. 尽管黄河口生态恢复工程极大的促进了湿地景观恢复,但本研究发现它的长期实施明显促进了N2 O的产生. 因而下一步生态恢复工程应统筹考虑景观恢复与温室气体削弱这两方面因素. 国内外学者亦采用类似研究方法对土壤N2 O的产生量进行了研究,但研究结果差别较大(表 5). 王玲玲[43]对黄河口潮滩土壤的研究发现,含水量和TN含量较高的光滩N2 O总产生量大于高潮滩,分别为0.653 nmol ·(kg ·h)-1和0.607 nmol ·(kg ·h)-1. 牟晓杰[44]对闽江口短叶茳芏和互花米草湿地土壤的研究发现,N2 O的总产生量分别为-0.51 nmol ·(kg ·h)-1 和38.38 nmol ·(kg ·h)-1,这主要是由于互花米草生长旺盛,生物量大,对土壤N吸收量大,导致其土壤N含量相对较低有关. Robertson等[45]对美国两种富氧森林土壤的研究发现,有机碳含量和pH较高的Rose Lake森林土壤N2 O总产生量相对较高,介于125~379 nmol ·(kg ·h)-1,而Warren Woods森林土壤N2 O总产生量相对较低,介于为4.46~41.96 nmol ·(kg ·h)-1. Webster等[33]对草原砂质壤土质地的含铁灰化土进行了研究,发现土壤含水量高,N2 O总产生量高,其介于384~564 nmol ·(kg ·h)-1. 另外,其N2 O总产生量明显高于其它地区,而这与其培养时间较长(50 h)有很大关系. Wrage等[16]对草原土壤的研究也发现,含水量高,N2 O总产生量高,其值介于34~184 nmol ·(kg ·h)-1. 整体而言,导致这些不同研究结果差异较大的原因可能与不同研究的土壤理化性质(如N含量、 pH、 质地、 盐分和水分条件等)以及实验培养条件(常温、 恒温和时间长短等)的差异有关. 据表 5可知,不同恢复阶段的湿地土壤,特别是恢复区土壤的有机碳含量要高于滨岸潮滩,但却明显低于闽江口湿地土壤、 芬兰草原土壤、 美国森林土壤以及英国草原含铁灰化土. 相关研究表明,大多数微生物从有机碳的氧化过程中获得所需的能源和基质,有机碳含量高可加速微生物活动,从而有助于N2 O的产生[32]. 此外,滨岸潮滩和芬兰草原土壤的培养实验是在室温条件下培养24 h,闽江口湿地土壤是在30℃下恒温培养,英国草原含铁灰化土是恒温培养50 h,而本研究为25℃下恒温培养24 h. 可见,培养条件的差异可能也是重要原因.
| 表 5 土壤N2 O产生量对比 Table 5 Comparison of N2 O production in soils |
(1)不同恢复阶段湿地土壤不同土层的N2 O总产生量明显不同,但总体均表现为N2 O释放. 恢复区土壤的N2 O产生量大于未恢复区.
(2)N2 O的产生主要以硝化作用、 硝化细菌反硝化作用和非生物作用为主,而反硝化作用对N2 O的产生存在较大的削弱作用. 非生物过程对N2 O产生量的较大贡献主要与黄河口为高活性铁区,Fe的还原作用关系密切.
(3)温度和水分对不同恢复阶段湿地土壤N2 O产生过程的影响不尽一致,与不同恢复阶段湿地土壤微生物活性对温度和水分的响应程度差异有关.
| [1] | Wang W C, Yung Y L, Lacis A A, et al. Greenhouse effects due to man-made perturbations of trace gases[J]. Science, 1976, 194 (4266): 685-690. |
| [2] | Bouwman A F. Exchange of greenhouse gases between terrestrial ecosystems and the atmosphere[A]. In: Bouwman A F. Soiland thegreenhouse effect, proceedings of the international conference on soils and the greenhouse effect[M]. Chichester: John Wiley and Sons, 1990. 61-127. |
| [3] | 孙志高, 刘景双, 杨继松, 等. 三江平原典型小叶章湿地土壤硝化-反硝化作用与氧化亚氮排放[J]. 应用生态学报, 2007, 18 (1): 185-192. |
| [4] | 蔡延江, 丁维新, 项剑. 土壤N2O 和NO 产生机制研究进展[J]. 土壤, 2012, 44 (5): 712-718. |
| [5] | 吴德礼, 傅旻瑜, 马鲁铭. 生物及化学反硝化过程中N2O的产生与控制[J]. 化学进展, 2012, 24 (10): 2054-2061. |
| [6] | Shingo U, Chun-Sim U G, Takahito Y, et al. Dynamics of dissolved O2, CO2, CH4 and N2O in a tropical coastal swamp in southern Thailand[J]. Biogeochemistry, 2000, 49 (3): 191-215. |
| [7] | Muñoz-Hincapié M, Morell J M, Corredor J E. Increase of nitrous oxide flux to the atmosphere upon nitrogen addition to red mangroves sediments[J]. Marine Pollution Bulletin, 2002, 44 (10): 992-996. |
| [8] | Gregorich E G, Hopkins D W, Elberling B, et al. Emission of CO2, CH4 and N2O from lakeshore soils in an Antarctic dry valley[J]. Soil Biology and Biochemistry, 2006, 38 (10): 3120-3129. |
| [9] | Wang D Q, Chen Z L, Wang J, et al. Summer-time denitrification and nitrous oxide exchange in the intertidal zone of the Yangtze Estuary[J]. Estuarine, Coastal and Shelf Science, 2007, 73 (1-2): 43-53. |
| [10] | 徐继荣, 王友绍, 殷建平, 等. 珠江口入海河段DIN形态转化与硝化和反硝化作用[J]. 环境科学学报, 2005, 25 (5): 686-692. |
| [11] | Zhu R B, Liu Y S, Ma J, et al. Nitrous oxide flux to the atmosphere from two coastal tundra wetlands in eastern Antarctica[J]. Atmospheric Environment, 2008, 42 (10): 2437-2447. |
| [12] | Sun Z G, Wang L L, Tian H Q, et al. Fluxes of nitrous oxide and methane in different coastal Suaeda salsa marshes of the Yellow River estuary, China[J]. Chemosphere, 2012, 90 (2): 856-865. |
| [13] | Sun Z G, Wang L L, Mou X J, et al. Spatial and temporal variations of nitrous oxide flux between coastal marsh and the atmosphere in the Yellow River estuary of China[J]. Environmental Science and Pollution Research, 2014, 21 (1): 419-433. |
| [14] | Cui B S, Yang Q C, Yang Z F, et al. Evaluating the ecological performance of wetland restoration in the Yellow River Delta, China[J]. Ecological Engineering, 2009, 35 (7): 1090-1103. |
| [15] | 董凯凯, 王慧, 杨丽原, 等. 人工恢复黄河三角洲湿地土壤碳氮含量变化特征[J]. 生态学报, 2011, 31 (16): 4778-4782. |
| [16] | Wrage N, Velthof G L, Laanbroek H J, et al. Nitrous oxide production in grassland soils: assessing the contribution of nitrifier denitrification[J]. Soil Biology and Biochemistry, 2004, 36 (2): 229-236. |
| [17] | 付素静. 干旱荒漠区典型土壤硝酸盐分布特征及硝化反硝化作用研究[D]. 兰州: 兰州大学, 2012. |
| [18] | Linn D M, Doran J W. Effect of water-filled pore space on carbon dioxide and nitrous oxide production in Tilled and Non-tilled soils[J]. Soil Science Society of America Journal, 1984, 48 (6): 1267-1272. |
| [19] | Magalhes C, Costa J, Teixeira C, et al. Impact of trace metals on denitrification in estuarine sediments of the Douro River estuary, Portugal[J]. Marine Chemistry, 2007, 107 (3): 332-341. |
| [20] | 朱永官, 王晓辉, 杨小茹, 等. 农田土壤N2O产生的关键微生物过程及减排措施[J]. 环境科学, 2014, 35 (2): 792-800. |
| [21] | 张树兰, 杨学云, 吕殿青, 等. 温度、 水分及不同氮源对土壤硝化作用的影响[J]. 生态学报, 2002, 22 (12): 2147-2153. |
| [22] | 李建兵, 黄冠华. NaCl对粉壤土氨挥发及硝化、 反硝化的影响[J]. 农业环境科学学报, 2006, 25 (4): 945-948. |
| [23] | 王维奇, 王纯, 刘白贵. 盐度对湿地枯落物分解过程中碳氮磷化学计量比的影响[J]. 中国环境科学, 2012, 32 (9): 1683-1687. |
| [24] | 牟晓杰, 刘兴土, 仝川, 等. 人为干扰对闽江河口湿地土壤硝化-反硝化潜力的影响[J]. 中国环境科学, 2013, 33 (8): 1413-1419. |
| [25] | 孙英杰, 吴昊, 王亚楠. 硝化反硝化过程中N2O 释放影响因素[J]. 生态环境学报, 2011, 20 (2): 384-388. |
| [26] | Strauss E A. The effects of organic carbon and nitrogen availability on nitrification rates in stream sediments[D]. Notre Dame, Indiana: University of Notre Dame, 2000. |
| [27] | 孙志高, 刘景双. 湿地土壤的硝化- 反硝化作用及影响因素[J]. 土壤通报, 2008, 39 (6): 1462-1467. |
| [28] | Flowers T H, O'Callaghan J R. Nitrification in soils incubated with pig slurry or ammonium sulphate[J]. Soil Biology and Biochemistry, 1983, 15 (3): 337-342. |
| [29] | Rückauf U, Augustin J, Russow R, et al. Nitrate removal from drained and reflooded fen soils affected by soil N transformation processes and plant uptake[J]. Soil Biology and Biochemistry, 2004, 36 (1): 77-90. |
| [30] | 宋金明, 李鹏程. 渤海南部沉积物中的活性铁及氧化还原环境[J]. 海洋科学, 1997, (4): 32-36. |
| [31] | 王纯杰. 辽河口湿地沉积物硝化反硝化速率时空变化与脱氮效应的影响因素[D]. 青岛: 中国海洋大学, 2012. 11-33. |
| [32] | Aulakh M S, Rennie D A, Paul E A. Gaseous nitrogen losses from cropped and summer fallow soils[J]. Canadian Journal of Soil Science, 1982, 62(1): 187-195. |
| [33] | Webster F A, Hopkins D W. Contributions from different microbial processes to N2O emission from soil under different moisture regimes[J]. Biology and Fertility of Soils, 1996, 22 (4): 331-335. |
| [34] | Wrage N, Velthof G L, Van Beusichem M L, et al. Role of nitrifierdenitrification in the production of nitrous oxide[J]. Soil Biology and Biochemistry, 2001, 33 (12-13): 1723-1732. |
| [35] | 孙文广, 甘卓婷, 孙志高, 等. 黄河口新生湿地土壤Fe和Mn元素的空间分布特征[J]. 环境科学, 2013, 34 (11): 275-282. |
| [36] | Joergensen R G, Brookes R C, Jenkinson D S. Survival of the soil microbial biomass at elevated temperatures[J]. Soil Biology and Biochemistry, 1990, 22 (8): 1129-1136. |
| [37] | Breuer L, Kiese R, Butterbach-Bahl K. Temperature and moisture effects on nitrification rates in tropical rain forest soils[J]. Soil Science Society of America Journal, 2002, 66 (3): 834-844. |
| [38] | Stottmeister U, Wiener A, Kuschk P, et al. Effects of plants and microorganisms in constructed wetlands for wastewater treatment[J]. Biotechnology Advances, 2003, 22 (1-2): 93-117. |
| [39] | Malhi S S, Mcgill W B. Nitrification in there Alberta soils: Effect of temperature, moisture and substrate concentration[J]. Soil Biology & Biochemistry, 1982, 14 (4): 393-399. |
| [40] | 黄国宏, 陈冠雄, 韩冰, 等. 土壤含水量与N2O产生途径研究[J]. 应用生态学报, 1999, 10 (1): 53-56. |
| [41] | Williams J R. Addressing global warming and biodiversity through forest restoration and coastal wetlands creation[J]. Science of the Total Environment, 1999, 240 (1-3): 1-9. |
| [42] | Jenkins W A, Murray B C, Kramer R A, et al. Valuing ecosystem services from wetlands restoration in the Mississippi Alluvial Valley[J]. Ecological Economics, 2010, 69 (5): 1051-1061. |
| [43] | 王玲玲. 黄河口滨岸潮滩湿地系统N2O排放规律与影响机制研究[D]. 烟台: 中国科学院烟台海岸带研究所, 2011. 69-76. |
| [44] | 牟晓杰. 闽江河口湿地碳氮循环关键过程对氮输入的响应[D]. 长春: 中国科学院东北地理与生态研究所, 2013. 84-94. |
| [45] | Robertson G P, Tiedjea J M. Nitrous oxide sources in aerobic soils: nitrification, denitrification and other biological processes[J]. Soil Biology and Biochemistry, 1987, 19 (2): 187-193. |