 2014, Vol. 35
2014, Vol. 35



2. 中国科学院大学, 北京 100049;
3. 南京农业大学公共管理学院, 南京 210095
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. College of Public Administration, Nanjing Agricultural University, Nanjing 210095, China
进入水体的光一部分会被吸收,水体对光的吸收特性可以用吸收系数表示,决定内陆水体光吸收特性的物质主要有4种: 纯水、 浮游植物、 非藻类颗粒物及黄色物质. 水体的总吸收系数等于各组分在该波段的吸收系数之和,即a=aw+aph+ad+aCDOM,式中,aw、 aph、 ad和aCDOM分别为纯水、 浮游植物、 非藻类颗粒物和黄色物质的吸收系数; 纯水的吸收系数一般为常数,关于其值已进行了较多测定[1]; 其它物质的吸收系数可以通过实验或者计算而得到. 大量已有研究表明,浮游植物、 非藻类颗粒物的吸收系数是随着水体营养水平[2]、 浮游植物群落结构[3,4]、 颗粒物粒径大小及色素组成[5, 6, 7, 8, 9]、 季节和空间[10, 11, 12, 13]等因素的变化而变化的量. 悬浮颗粒物特别是浮游植物吸收及比吸收系数是建立水色遥感模型的关键参数[14, 15, 16],伴随着遥感技术的快速发展和广泛应用,对悬浮颗粒物吸收特性的研究将为水色遥感模型的建立以及反演精度的提高提供输入参数. 目前关于内陆水体悬浮颗粒光学特性的研究较多,但主要集中在浑浊的富营养化浅水湖泊[17, 18, 19, 20, 21, 22],对水质良好的水库型湖泊如千岛湖水体悬浮颗粒物吸收特性的测定和研究很少. 因此,开展千岛湖不同季节悬浮颗粒物吸收特性及其影响因素研究,有利于进一步丰富不同营养水平、 不同类型以及不同浮游植物主导水体的生物光学特性,可以更好地服务于湖泊光学分类及水质遥感监测.
千岛湖又称新安江水库,是1959年因建造新安江水电站蓄水筑坝而形成的人工深水湖泊,属钱塘江水系. 千岛湖呈树枝型,岸线总长度1406 km,分为东北、 东南、 西北、 西南、 中心五大湖区[23]. 千岛湖历来以四周树木葱郁,生物多样性丰富,湖水清澈澄明,环境优美而先后被评为国家级重点风景名胜区和国家级森林公园[24]. 可以说千岛湖的水环境质量对周边的经济和社会发展具有极其重要的影响,因而有关其营养盐分布、 变化、 来源和水体的营养水平的研究也备受关注[23, 24, 25, 26]. 尤其是自1998年和1999年发生区域蓝藻水华后,对千岛湖水体富营养化发生机制及防治对策方面的研究更引起了广大学者的重视[27,28]. 另外,千岛湖作为特大型人工深水水库,上层水体光学参数均一,是当前内陆湖泊(水体) 水质遥感的理想研究对象[29]. 本研究从千岛湖冬、 夏季的实测数据入手,分析水中悬浮颗粒物的吸收特性季节和空间差异以及与湖泊营养水平之间的相互关系,旨在加深对此大型水库固有光学特性的了解,填补千岛湖水体固有光学性质研究的空白,同时为初级生产力的估算、 光辐射传输及水色遥感提供基本参数.
1 材料与方法 1.1 采样时间和站位
于冬季(2012年12月、 2013年1~2月)和夏季(2013年6~8月)每个月月初在千岛湖采集水样,共布设8个采样点(图 1),采样区域分布全湖. 选择风平浪静和小风浪的天气,使用中国科学院水生生物研究所研制的有机玻璃采水器采集表层水样(0~0.5m),现场过滤留膜并且将一定水样装入干净的塑料瓶置于冷藏箱内黑暗低温保存,全部带回实验室分析营养盐参数、 颗粒物吸收以及浮游植物色素浓度等. 透明度(SD)现场用30 cm透明度盘测定. 其中7月采样时因降雨影响街口点水体浑浊,分析中予以剔除,而8月西园数据缺失. 1.2 理化参数测定
总氮(TN)、 总磷(TP)测定采用碱性过硫酸钾消解紫外、 钼锑抗显色测定. 叶绿素a的测定采用分光光度法,用GF/F滤膜过滤水样,然后将滤膜置于冰箱中冷冻48 h以上,取出用90%的热乙醇萃取,然后在岛津UV-2550分光光度计上测定665、 750 nm 处吸光度,并加入1滴1%稀盐酸酸化,换算得到叶绿素a和脱镁叶绿素浓度. 浮游植物色素浓度为叶绿素a和脱镁叶绿素浓度之和[30],用Chla表示.
 | 图 1 千岛湖采样点分布示意Fig. 1 Location of sampling stations in Lake Qiandaohu |
悬浮颗粒物的吸收采用定量滤膜技术(QFT)测定,用直径25 mm的GF/F(Whatman公司)滤膜过滤一定体积的水样(根据水里悬浮物浓度决定过滤的体积),在UV-2550PC型分光光度计下用ISR-240A型积分球采用光透射-光反射(T-R)方法测定滤膜上颗粒物的吸光度. 采用Tassan 等[31]提出的如下公式进行放大因子校正:
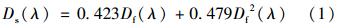
式中,Ds(λ)为校正后的滤膜上悬浮颗粒物吸光度; Df(λ)为直接在仪器上测定的滤膜上悬浮颗粒物吸光度.
滤膜上悬浮颗粒物的光谱吸收系数按下式进行计算得到[32]:
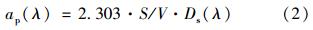
式中,V为被过滤水样的体积,S为沉积在滤膜上的颗粒物的有效面积.
用次氯酸钠浸泡滤膜将色素萃取掉,滤膜上只剩下非藻类颗粒物,使用同样浸泡过的空白滤膜作参比,按与总颗粒物吸收系数同样的方法测定得到非藻类吸收系数. 由于浮游颗粒物的吸收是非藻类颗粒物和浮游植物吸收的简单线性叠加,因而由总悬浮物颗粒物的光谱吸收系数ap(λ)减去非藻类的光谱吸收系数ad(λ)就得到浮游植物的光谱吸收系数aph(λ)[33].
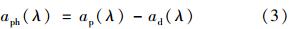
浮游植物比吸收系数是单位色素浓度吸收能力的大小,可以表示为浮游植物吸收系数aph(λ)与浮游植物色素浓度(Chla)的比值,即:
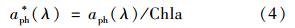
1.4 水体营养水平评价
国内常用的评价湖泊富营养化程度的基本方法有:营养状态指数法(卡尔森营养状态指数,TSI)、 修正的营养状态指数、 综合营养状态指数(TLI)、 营养度指数法、 评分法[34]. 以上5种方法在实际工作中都被采用,其中营养度指数法计算步骤繁琐、 耗时长,不如综合营养指数法简便易行; 评分法在实际应用过程中,受人为因素的干扰较多,影响结果的准确性. 因此,本研究选取综合营养状态指数法评价湖泊富营养化程度.
对千岛湖水体营养状态评价重点选取了与水体富营养化密切相关的监测项目:Chla、 TN、 TP、 SD这4项. 综合营养状态指数为:
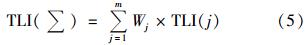
式中,TLI(
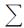 )为综合营养状态指数; Wj为第j种参数的营养状态指数的相关权重; TLI(j)为第j种参数的营养状态指数.
)为综合营养状态指数; Wj为第j种参数的营养状态指数的相关权重; TLI(j)为第j种参数的营养状态指数. 以Chla作为基准参数,则第j 种参数的归一化的相关权重计算公式见表 1[35].
| 表 1 中国湖泊部分参数与叶绿素a的相关关系r及r2值Table 1 Correlation between selected parameters and chlorophyll a in lakes of China |
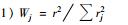
各项目营养状态指数计算式如下:
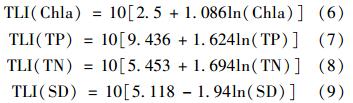
采用0~100的一系列连续数字对湖泊营养状态进行分级: TLI()<30为贫营养; 30≤TLI()≤50为中营养; TLI()>50为富营养,其中50<TLI()≤60为轻度富营养,60<TLI()≤70 为中度富营养,TLI()>70为重度富营养.
1.5 统计分析
运用SPSS 19.0进行数据统计分析,包括计算平均值、 标准差和方差等. 季节间参数差异性分析采用独立样品t检验,显著性水平P值小于0.05表示存在显著性差异. 2 结果与分析 2.1 营养状态评价
千岛湖各采样点冬夏季的Chla、 TN、 TP、 SD及TLI()数据见表 2,显然千岛湖的综合营养状态指数较低,全湖综合营养状态指数在30~40之间,属中营养水平. 千岛湖夏季综合营养状态指数显著高于冬季(P<0.005),12月及1月综合营养状态指数均较小,6、 7月随着降雨增多,地表径流量和入湖营养盐增加,伴随浮游植物生物量增加和透明度下降,综合营养状态指数增加,街口和小金山两个点达到50,属轻度富营养. 就全湖来看,西北(街口-小金山一线)及东北(航头岛)湖区的营养水平相对较高,因为街口和航头岛分别位于千岛湖流域的主要来水河流新安江和富强溪的下游入湖口; 西南、 东南及湖心区的营养水平较低,表明入湖携带的物质被逐渐稀释、 沉淀等.
| 表 2 千岛湖冬季和夏季的色素浓度(Chla)、 总氮(TN)、 总磷(TP)、 透明度(SD)及综合营养状态指数TLI()1)Table 2 Parameter values of four assessment indexes and the evaluation results of eutrophication level in Lake Qiandaohu |
 | 图 2 千岛湖冬季总悬浮颗粒物、 浮游植物、 非藻类颗粒物吸收光谱 Fig. 2 Absorption spectra of total suspended particles,phytoplankton,tripton and their mean values in winter |
 | 图 3 千岛湖夏季总悬浮颗粒物、 浮游植物、 非藻类颗粒物吸收光谱Fig. 3 Absorption spectra of total suspended particles,phytoplankton,tripton and their mean values in summer |
图 2(d)和图 3(d)是对千岛湖冬季和夏季400~700 nm总悬浮颗粒物、 浮游植物和非藻类颗粒物吸收系数均值做出的光谱曲线,从图 2(d)可明显地看出冬季除了660~695 nm的浮游植物色素强吸收波段外,非藻类颗粒物吸收贡献要大于浮游植物吸收; 而夏季[图 3(d)]浮游植物吸收大于非藻类颗粒物吸收. 2.3 浮游植物吸收系数
440 nm和675 nm附近是浮游植物的特征吸收波段,675 nm主要是叶绿素a贡献,而440 nm除了叶绿素还可以反映辅助色素的贡献. 千岛湖冬季浮游植物吸收的光谱曲线见图 2(b),其中小金山的吸收值最大明显高于其它点,三潭岛和茅头尖的最小; 440 nm和675 nm吸收系数aph(440)、 aph(675)变化范围分别为:0.06~0.21 m-1、 0.03~0.11 m-1; 均值分别为(0.10±0.03)m-1、 (0.05±0.02)m-1. 夏季浮游植物光谱曲线如图 3(b)所示,其中,街口和小金山吸收值最大,三潭岛、 大坝的最小. 夏季440 nm和675 nm附近的吸收峰明显,aph(440)、 aph(675) 变化范围分别为:0.05~0.64 m-1、 0.02~0.28 m-1; 均值(0.17±0.14)m-1和(0.08±0.07)m-1,空间差异并不大. 但其随季节变化明显,夏季吸收值远远大于冬季(P<0.005).
浮游植物吸收系数主要受水体中浮游植物的色素浓度及群落组成的影响,随着Chla浓度的变化而变化,冬季相应的Chla变化范围:1.27~5.00 μg ·L-1,均值为(2.24±0.83)μg ·L-1; 夏季变化范围:1.21~17.291 μg ·L-1,均值为(4.93±4.40)μg ·L-1; 冬季显著低于夏季(P<0.05). 图 4(a)和 4 (b) 分别为冬季440 nm和675 nm浮游植物吸收系数与Chla浓度的拟合关系,可以看出浮游植物的吸收系数随着Chla浓度的增加呈线性增加,675 nm相关性更高些. 因为冬季浮游植物浓度低,440 nm处受其它色素含量的影响相关性降低. 图 4 (c) 和 4 (d)为夏季440 nm和675 nm浮游植物吸收系数与Chla浓度的拟合关系,可以看出二者相关性和冬季一致即浮游植物吸收系数随着Chla浓度增加呈线性变化,但夏季的相关性要明显高于冬季,且相关系数都在0.8以上.
 | 图 4 浮游植物吸收系数aph(440)、 aph(675)与Chla浓度的关系Fig. 4 Correlations between phytoplankton absorption coefficients aph(440),aph(675) and chlorophyll a concentration in Lake Qiandaohu |
浮游植物比吸收系数表征的是单位色素浓度对光的吸收能力,它随水体营养水平、 光照条件和浮游植物群落结构的变化而变化. 浮游植物比吸收系数a*ph(λ)是建立浮游植物生物量与色素光谱吸收关系的重要参数[3],可以将现场的生物光学测量与卫星遥感数据相联系,在利用卫星遥感估测水体初级生产力中发挥着重要作用[36]. 千岛湖冬季各点之间的浮游植物比吸收系数变化较小,其中三潭岛和茅头尖的相对较大,街口相对较小. 冬季440 nm和675 nm浮游植物比吸收系数a*ph(440)、 a*ph(675)的变化范围:0.030~0.068 m2 ·mg-1、 0.016~0.033 m2 ·mg-1; 均值分别为:(0.045±0.010)m2 ·mg-1、 (0.022±0.004)m2 ·mg-1. 夏季东南和西南湖区比吸收系数相对较大,西北湖区最小. 夏季440 nm和675 nm浮游植物比吸收系数a*ph(440)、 a*ph(675)的变化范围:0.020~0.071 m2 ·mg-1、 0.010~0.029 m2 ·mg-1sup>; 均值:(0.039±0.013)m2 ·mg-1、 (0.019±0.005)m2 ·mg-1,低于冬季. 对千岛湖浮游植物比吸收系数和Chla浓度进行相关性分析发现,440 nm和675 nm均存在幂函数关系(图 5); 浮游植物比吸收系数随着Chla浓度增加逐渐减小,但冬季和夏季Chla浓度含量大都集中在较低的浓度范围.
如图 2(c)和图 3(c)所示,非藻类颗粒物吸收系数随着波长增加呈指数递减. 冬季和夏季非藻类颗粒物的吸收系数ad(440)的变化范围分别为:0.03~0.25 m-1、 0.03~0.22 m-1,均值分别为(0.1±0.07)m-1、 (0.07±0.04)m-1,冬季和夏季的差异不大(P≥0.05). 各湖区之间,西北湖区非藻类颗粒物吸收最大,东南湖区最小. 对千岛湖水体的ad(440)、 ad(675)与总悬浮物浓度(TSS)、 无机颗粒物(ISS)和有机颗粒物(OSS)进行线性回归分析(表 3),冬季ad(440)、 ad(675)与有机颗粒物没有线性相关性,但与总悬浮颗粒物和无机颗粒物的相关性显著,与无机颗粒物的相关性最好,说明千岛湖冬季非藻类颗粒物主要以无机颗粒物为主; 夏季ad(440)与总悬浮颗粒物和有机颗粒物呈线性关系,随着总悬浮颗粒物和有机颗粒物含量增加440 nm附近非藻类颗粒物吸收增加,而与无机颗粒物没有显著相关,说明夏季非藻类颗粒物吸收主要是有机碎屑引起.
通常总悬浮颗粒物的吸收光谱曲线有3种类型:①与浮游植物色素的吸收相似; ②与非藻类颗粒物的吸收相似; ③与非藻类颗粒物和浮游植物色素的吸收光谱都不相似,但不管是哪种,总颗粒物在675 nm附近的吸收曲线跟浮游植物在此波段的吸收曲线均是非常相似的,这主要是由于浮游植物在此波段的特征吸收所致[37]. 对于第①种类型,浮游植物吸收系数一般都大于非藻类颗粒物吸收系数,总颗粒物吸收系数在675 nm、 440 nm有2个峰值,675 nm峰值非常明显,而440 nm处由于受非藻类颗粒物吸收系数影响有时表现的不是特别明显,这种光谱类型的站点主要分布在Chla浓度较高,同时无机颗粒物浓度较低,有机颗粒物占总颗粒物比例超过40%的站点. 对于第②种类型,总颗粒物吸收系数只在675 nm有个峰值,浮游植物吸收可能大于非藻类颗粒物吸收,其他波段浮游植物吸收均淹没在非藻类的吸收光谱中,这类站点主要分布在Chla浓度低以及无机颗粒物浓度高的地方. 对于第③种类型,非藻类颗粒物和浮游植物的吸收大致各占50%左右,在450 nm以下波段非藻类颗粒物吸收系数大于浮游植物,在600 nm以上波段非藻类颗粒物吸收系数大于浮游植物,其他波段交替出现大小,对应的为有机颗粒物占总颗粒物比例在30%~40%之间的站点.
对千岛湖8个采样点逐月的分析显示大概可以分为两类,其一,东南湖区的三潭岛、 大坝以及富文这3个点(称为第一类采样点)无论冬季和夏季,浮游植物吸收系数所占比例均在50%以上; 其二,剩余的5个采样点(称为第二类采样点),冬季以非藻类颗粒物吸收为主,而夏季则以藻类颗粒物吸收为主. 图 6所示为千岛湖第一类采样点冬季[图 6(a)]和夏季[图 6(b)]浮游植物及非藻类颗粒物吸收占总悬浮颗粒物吸收比例,浮游植物的吸收无论在夏季还是在冬季都大于非藻类颗粒物的吸收,总悬浮颗粒物的吸收光谱与浮游植物吸收类似; 图 6(c)和6(d)为第二类采样点冬季和夏季浮游植物及非藻类颗粒物吸收占总悬浮颗粒物吸收比例,冬季400~600 nm非藻类颗粒物所占的比例高出了浮游植物吸收占总悬浮颗粒物吸收比例,600~700 nm浮游植物吸收占得比例要高于非藻类颗粒物,675 nm达到最大值71.9%,因此可以确定这一类采样点冬季总悬浮颗粒物的吸收光谱曲线属于第二种类型与非藻类颗粒物吸收类似. 夏季,400~700 nm浮游植物吸收均占到50%以上,远大于非藻类颗粒物吸收,675 nm达到最大值86.9%,因此夏季千岛湖总悬浮颗粒物的吸收光谱曲线与浮游植物光谱曲线类似,属第一种类型.
千岛湖冬季和夏季营养状态指数差异显著,夏季高于冬季,类似的浮游植物吸收特性也随季节变化有显著性差异,如图 7所示,冬季和夏季440 nm和675 nm浮游植物吸收系数与营养状态指数有很好的线性相关性,随着营养状态指数增加,浮游植物吸收系数也不断增加; 夏季的相关性比冬季好且随着营养状态指数的增加变化快. 440 nm和675 nm浮游植物比吸收系数随季节的变化与营养状态指数相反,表现为冬高夏低,但浮游植物比吸收系数与营养状态指数之间的关系并不显著(r2极低,文中未列出).
将千岛湖夏季浮游植物吸收特性和重富及中富营养的太湖湖区、 轻度富营养的天目湖和中营养的博斯腾湖[38]比较,和3.2节的分析一致,440 nm和675 nm浮游植物吸收系数随着水体营养水平增加逐渐变大,中营养的千岛湖和博斯腾湖相近小于其它3个; 比吸收系数变化规律刚好和吸收系数相反,随着营养水平的增加而变小,也即千岛湖和博斯腾湖的比吸收最大(表 4). 这种变化规律可能的原因一方面水体的营养水平不同,浮游植物群落结构也不同,在营养水平低的千岛湖主要是以粒径较大的硅藻和隐藻为主(文中没列出此部分数据),而太湖主要是粒径小的蓝藻占绝对优势[7]; 另一方面不同的浮游植物体内的色素含量和组成也不尽相同,营养水平越高的水体色素包裹效应越强[39]使得浮游植物比吸收减小.
(1)千岛湖悬浮颗粒物吸收有一定的空间差异,营养水平最低的东南湖区,总悬浮颗粒物吸收及浮游植物吸收系数较小,浮游植物比吸收系数较大; 西北湖区则刚好相反.
(2)千岛湖夏季总悬浮颗粒物吸收光谱与浮游植物吸收相似,非藻类颗粒物吸收系数与总悬浮物、 有机颗粒物呈线性关系; 冬季东南湖区总悬浮颗粒物吸收光谱与浮游植物吸收类似,剩余站点与非藻类颗粒物相似,非藻类颗粒物吸收系数则与无机颗粒物相关性最好.
(3)440 nm和675 nm浮游植物吸收系数与Chla浓度存在显著线性相关; 比吸收系数和Chla浓度均存在显著幂函数负相关关系,表明随Chla浓度增加由于色素包裹效应造成比吸收降低. 湖泊的营养程度明显影响到浮游植物吸收、 比吸收等生物光学特性,随着营养状态增加由于浮游植物生物量增加以及群落结构发生变化导致其吸收系数增大,而比吸收则减小. 致谢: 特别感谢杭州市环境保护科学研究院的虞左明和韩轶才教授、 刘明亮、 何剑波、 殷燕工程师以及淳安县环境保护监测站的吴志旭站长和其他工作人员在采样过程中给予的帮助和支持.

图 5 浮游植物比吸收系数与Chla浓度的关系Fig. 5 Correlations between chlorophyll-specific absorption and chlorophyll a concentration ![]()
表 3 千岛湖冬季和夏季非藻类颗粒物吸收ad(440)、 ad(675)与总悬浮物(TSS)、 无机悬浮物(ISS)、 有机悬浮物(OSS)的线性回归Table 3 Linear models between tripton absorption at 440 ad(440) and 675 nm ad(675) and total suspended
solids (TSS),inorganic suspended solids (ISS),organic suspended solids (OSS) in Lake Qiandaohu 
图 6 浮游植物和非藻类颗粒物吸收占总悬浮颗粒物吸收比例Fig. 6 Relative contribution of phytoplankton and tripton to absorption of total suspended particles 
图 7 冬季和夏季浮游植物吸收系数与营养状态指数的关系Fig. 7 Correlations between phytoplankton absorption coefficients aph(440),aph(675) and trophic level index (TLI) ![]()
表 4 不同营养水平湖泊夏季440 nm和675 nm浮游植物吸收系数aph(λ)和比吸收系数a*ph(λ)对比1)[38]
Table 4 Comparison of phytoplankton absorption coefficients and chlorophyll-specific absorption coefficients at 440 nm and 675 nm in summer
| [1] | 李宁, 李学刚, 宋金明. 海洋碳循环的关键生物地球化学过程[J]. 海洋环境科学, 2005, 24 (2): 75-80. |
| [2] | 张述伟, 王江涛, 李宁, 等. 2010年春季长江口邻近海区水体中溶解有机碳、氮的分布特征及其影响因素[J]. 海洋环境科学, 2013, 32 (1): 33-38. |
| [3] | 张乃星, 宋金明, 贺志鹏. 海水颗粒有机碳(POC)变化的生物地球化学机制[J]. 生态学报, 2006, 26 (7): 2328-2339. |
| [4] | 刘瑞玉, 罗秉征. 三峡工程对长江口及邻近海域生态与环境的影响[J]. 海洋科学集刊, 1992, 33: 1-13. |
| [5] | 林以安, 唐仁友, 李炎. 长江口区C、N、P的生物地球化学变化对悬浮体凝絮沉降的影响[A]. 见: 张经(主编). 中国主要河口的生物地球化学研究——化学物质的迁移与环境[M]. 北京: 海洋出版社, 1996. 133-145. |
| [6] | Zhang J, Wu Y, Jennerjahn T C, et al. Distribution of organic matter in the Changjiang (Yangtze River) Estuary and their stable carbon and nitrogen isotopic ratios: Implications for source discrimination and sedimentary dynamics [J]. Marine Chemistry, 2007, 106 (1-2): 111-126. |
| [7] | 林晶, 吴莹, 张经, 等. 最大浑浊带对长江口有机碳分布的影响初探[J]. 海峡科学, 2009, (6): 150-152. |
| [8] | Cauwet G, Mackenzie F. Carbon inputs and distribution in estuaries of turbid rivers: the Yangtze and Yellow rivers (China) [J]. Marine Chemistry, 1993, 43 (1-4): 235-246. |
| [9] | Milliman J D, Xie Q C, Yang Z S. Transfer of particulate organic carbon and nitrogen from the Yangtze River to the ocean [J]. American Journal of Science, 1984, 284 (7): 824-834. |
| [10] | Cai D L, Tan F C, Edmond J M. Sources and transport of particulate organic carbon in the Amazon River and estuary [J]. Estuarine, Coastal and Shelf Science, 1988, 26 (1): 1-14. |
| [11] | Wang X C, Chen R F, Gardner G B. Sources and transport of dissolved and particulate organic carbon in the Mississippi River estuary and adjacent coastal waters of the northern Gulf of Mexico [J]. Marine Chemistry, 2004, 89 (1-4): 241-256. |
| [12] | Abril G, Nogueira M, Etcheber H, et al. Behaviour of organic carbon in nine contrasting European estuaries [J]. Estuarine, Coastal and Shelf Science, 2002, 54 (2): 241-262. |
| [13] | 张向上. 黄河口有机碳的时空分布及影响因素研究[D]. 青岛: 中国海洋大学, 2004. 19-20. |
| [14] | Zhang S, Gan W B, Ittekkot V. Organic matter in large turbid rivers: the Huanghe and its estuary [J]. Marine Chemistry, 1992, 38 (1-2): 53-68. |
| [15] | 蔡艳雅, 韩舞鹰. 珠江口有机碳的研究[J]. 海洋环境科学, 1990, 9 (2): 8-13. |
| [16] | 刘庆霞, 黄小平, 张霞, 等. 2010年夏季珠江口海域颗粒有机碳的分布特征及其来源[J]. 生态学报, 2012, 32 (14): 4403-4412. |
| [17] | 谢肖勃, 戴敏英, 苗辉, 等. 长江口区海水中溶解有机碳和颗粒有机碳的分布及变化的研究[J]. 海洋科学集刊, 1992, 33: 131-137. |
| [18] | 林晶. 长江口及其毗邻海区溶解有机碳和颗粒有机碳的分布[D]. 上海: 华东师范大学, 2007. 30-47. |
| [19] | 李宁. 长江口与胶州湾海水有机碳的分布、来源及与氮、磷的耦合关系[D]. 青岛: 中国科学院海洋研究所, 2006. 1-85. |
| [20] | 张岩松, 章飞军, 郭学武, 等. 东海秋季典型站位沉降颗粒物通量[J]. 海洋与湖沼, 2006, 37 (1): 28-34. |
| [21] | 宋晓红, 石学法, 蔡德陵, 等. 三峡截流后长江口秋季TSM、POC和PN的分布特征[J]. 海洋科学进展, 2007, 25 (2): 168-177. |
| [22] | Tipping E, Marker A F H, Butterwick C, et al. Organic carbon in the Humber rivers [J]. Science of the Total Environment, 1997, 194-195: 345-355. |
| [23] | 王江涛, 于志刚, 张经. 鸭绿江口溶解有机碳的研究[J]. 青岛海洋大学学报, 1998, 28 (3): 471-475. |
| [24] | lvarez-Salgado X A, Miller A E J. Dissolved organic carbon in a large macrotidal estuary (the Humber, UK): behaviour during estuarine mixing [J]. Marine Pollution Bulletin, 1999, 37 (3-7): 216-224. |
| [25] | 张龙军, 宫萍, 张向上. 河口有机碳研究综述[J]. 中国海洋大学学报, 2005, 35 (5): 737-744. |
| [26] | 金海燕, 林以安, 陈建芳, 等. 黄海、东海颗粒有机碳的分布特征及其影响因子分析[J]. 海洋学报, 2005, 27 (5): 46-53. |
| [27] | Hung J J, Lin P L, Liu K K. Dissolved and particulate organic carbon in the southern East China Sea [J]. Continental Shelf Research, 2000, 20 (4-5): 545-569. |
| [28] | 陈吉余, 陈沈良. 长江口生态环境变化及对河口治理的意见[J]. 水利水电技术, 2003, 34 (1): 19-25. |
| [29] | 胡利民, 邓声贵, 郭志刚, 等. 夏季渤海湾及邻近海域颗粒有机碳的分布与物源分析[J]. 环境科学, 2009, 30 (1): 39-46. |
| [30] | 吴家林, 石晓勇, 李克强, 等. 夏季黄东海颗粒有机碳的分布特征[J]. 中国海洋大学学报, 2012, 42 (增刊): 150-156. |
| [31] | Loh A N, Bauer J E. Distribution, partitioning and fluxes of dissolved and particulate organic C, N and P in the eastern North Pacific and Southern Oceans [J]. Deep Sea Research Part I: Oceanographic Research Papers, 2000, 47 (12): 2287-2316. |
| [32] | Becquevort S, Bouvier T, Lancelot C, et al. The seasonal modulation of organic matter utilization by bacteria in the Danube-Black Sea mixing zone [J]. Estuarine, Coastal and Shelf Science, 2002, 54 (3): 337-354. |
| [33] | 李炎. 杭州湾锋面颗粒态化学物质的输移: 潮周期相应和过滤作用[A]. 见: 张经(主编). 中国主要河口的生物地球化学研究——化学物质的迁移与环境[M]. 北京: 海洋出版社, 1996. 78-91. |
| [34] | Kawabe M, Kawabe M. Factors determining chemical oxygen demand in Tokyo Bay [J]. Journal of Oceanography, 1997, 53 (5): 443-453. |
| [35] | Wei X G, Shen C D, Sun Y M, et al. Characteristic of the organic carbon-isotope composition and contribution of suspended matter in the Pearl River [J]. Scientia Geographica Sinica, 2003, 23 (4): 471-476. |
| [36] | 周伟华, 袁翔城, 霍文毅, 等. 长江口邻域叶绿素a和初级生产力的分布[J]. 海洋学报, 2004, 26 (3): 143-150. |
| [37] | Cifuentes L A, Sharp J H, Fogel M L. Stable carbon and nitrogen isotope biogeochemistry in the Delaware estuary [J]. Limnology and Oceanography, 1988, 33 (5): 1102-1115. |
| [38] | Chang J, Shiah F K, Gong G C, et al. Cross-shelf variation in carbon-to-chlorophyll a ratios in the East China Sea, summer 1998 [J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2003, 50 (6): 1237-1247. |