 2014, Vol. 35
2014, Vol. 35



蓝藻是水源水体中危害最大的藻种,其中的微囊藻属是最普遍的代表属[1, 2, 3, 4, 5]. 大多数蓝藻细胞内存在气囊,为蓝藻提供浮力,使其悬浮于适宜的水深,以获得最适宜光照条件而大量繁殖. 蓝藻细胞内气囊为柱状结构,横断面为六边形,气囊壁为蛋白质结构,能透过气体,不能透过水,能抵御400~700 kPa的外压,当外压超出这一压强时气囊就不可逆转地破裂. 蓝藻细胞内的气囊总是在不停地组装和破裂,表层水体中的蓝藻利用充足的光照合成大量淀粉,导致细胞体重增加,且细胞内渗透压增大,衰老气囊在较大的渗透压作用下破裂,浮力减小,当体重大于浮力时,蓝藻开始下沉; 随着蓝藻的下沉,光照度逐步减弱,合成物质减少,渗透压减小,气囊的破裂速度小于气囊的组装速度,同时由于呼吸作用消耗细胞内储藏物质,体重减小,当体重与浮力相等时,蓝藻悬浮于水中; 当蓝藻体重小于浮力时,蓝藻开始上浮[6, 7, 8, 9]. 自然条件下,蓝藻呈现上下少、 中间多的竖向分布,最大藻类数量出现在水体2~3倍透明度处[10,11]. 此外,衰老和死亡的蓝藻重量降低,会一直漂浮在水面形成水华. 湖泊和水库平缓的水流及水体分层特性为蓝藻的悬浮生长提供了良好的水力条件[12, 13, 14]. 可采用空气混合上下水层的方法干扰蓝藻在湖泊水库中的生长环境,目前国内外采用的空气混合技术包括空气羽混合[15,16]和扬水曝气混合[17, 18, 19, 20, 21]两种,利用压缩空气混合水库上下层水体,将上层水体的蓝藻输送到下层水体,使其在弱光[22]、 低温[23]、 高压等胁迫条件下生长受到抑制. 空气混合控制水体蓝藻过程中,必须有足够大的混合强度,使得向下流动的水流速度大于蓝藻上浮速度,才能将上层藻类带到下层. 本研究的主要目的是探索水体中蓝藻在不同光照度、 压力、 温度件下的运动特性,以期为空气混合技术抑制蓝藻提供理论依据和技术支撑.
本实验所使用的藻样取自江苏太湖梅梁湾,取样时间为2011~2013年的7、 8、 9月,优势藻种为铜绿微囊藻,图 1为利用显微镜放大400倍后看到的水样中的蓝藻. 水样取回后放入有机玻璃培养装置内,并提供光照和慢速搅拌条件进行培养待用.
蓝藻上浮/下沉速度测定装置如图 2所示,有机玻璃柱上下端封闭,高3.2 m,直径150 mm,沿高度每隔0.5 m设一个取样口,在柱顶开设进样口与原水箱相连,柱底设放空阀. 柱顶与加压泵出口连接,通过调节加压泵回流管上的阀门开启度调节加压压力,同时有机玻璃柱安装在人工气候室内,室内温度及光照度分别由空调和日光灯控制,水温用冰块调节.
 | 图 1 水样中蓝藻显微图片
Fig. 1 Micrograph of Cyanobacteria
|
 | 图 2 藻类上浮(下沉)速度测定装置
Fig. 2 Algae floating (subsiding) velocity measuring device
|
控制实验用水样温度为(25±1)℃,实验光照度分别为:0、 100、 800、 1500、 3000、 6000、 10000、 15000、 25000 lx,实验压强为常压. 将水样从有机玻璃培养装置中取出放入原水箱中,充分搅拌均匀,取样测定水样初始叶绿素a质量浓度c0. 将水样注入有机玻璃柱内,在初始状态,各种上浮速度的藻颗粒在有机玻璃柱内竖向是均匀分布的. 由于水样中蓝藻颗粒的上浮速度有大有小,经过t时段静置后,有机玻璃柱内上下层藻类分布出现了明显的差别. 设某一藻颗粒上浮速度为v,若它从有机玻璃柱最底端出发,经过t时间后,它上浮的距离为h=v ·t,则在距离最底端h位置处及其以下已测不到该颗粒,而上浮速度小于v的藻颗粒仍然能被测到,它们的质量浓度为cht,则上浮速度小于v(v=h/t)的蓝藻占总量的比例为cht/c0,而上浮速度大于v的藻类占总量的比例为E=1-cht/c0.
控制实验用水样温度为(25±1)℃,实验光照度为6000 lx,当注满水时调节加压泵回流管阀门开启度,调节有机玻璃柱内压强分别为0、 0.1、 0.2、 0.3、 0.4、 0.5、 0.6 MPa,其余方法同1.3.1节. 测定在不同压强条件下经过t时段后有机玻璃柱内各个取样口叶绿素a质量浓度cht. 设取样口距有机玻璃柱顶的距离为h,可计算出下沉速度大于等于v(v=h/t)的藻颗粒的比例为E=1-cht/c0.
空调控制室温,冰块调节实验用水样温度分别为25、 20、 17、 8℃,实验光照度为6000 lx,在常压下进行藻类上浮/沉淀实验,其余方法同1.3.1节. 测定在不同温度条件下经过t时段后有机玻璃柱内各个取样口叶绿素a质量浓度cht.
为了模拟扬水曝气混合条件下蓝藻被混合到不同水深处,受到光照度、 温度、 压力等综合胁迫条件影响下的运动特性,参考黑河水库不同水深处的光照度、 温度和压强条件,作为本实验水深综合条件,见表 1.
| 表 1 水库不同水深综合条件 Table 1 Integrated conditions in different depths of the water |
用冰块调节水温,日光灯调节光照度,加压泵提供实验所需压强,其余方法同1.3.1节,测定在不同水深综合条件下原水样叶绿素a质量浓度c0、 经过t时段后有机玻璃柱内各个取样口叶绿素a质量浓度cht及蓝藻粒径分布.
藻类叶绿素a采用分光光度法测定[24],丙酮提取时间为24 h,测定仪器型号为UV759S紫外-可见分光光度计. 各取样口蓝藻粒径的测定采用丹东百特公司生产的BT-9300ST激光粒度分布仪. 藻细胞场发射超薄切片透射电镜扫描委托扬州大学测试中心测定,透射电镜为美国FEI公司生产的Tecnai G2 F30-TWIN型.
共进行了9种不同光照度下蓝藻运动特性的测定. 经过一定的静置时间,各取样口水样的叶绿素a质量浓度均小于初始值,且下层质量浓度小于上层,说明藻类发生了上浮. 绘制不同光照度下蓝藻上浮速度分布曲线,如图 3所示. 由于25000 lx光照度下蓝藻上浮趋势不明显,故在图中未绘出.
根据图 3绘制不同光照度下蓝藻上浮速度分别大于0.4、 0.8、 1.2cm ·min-1时所占比例,见图 4. 从中可知,蓝藻在光照度1500~6000 lx范围内均表现出较强的上浮性能,光照度为1500 lx时,上浮速度大于0.4、 0.8、 1.2 cm ·min-1的蓝藻分别占总量的72.5%、 58%、 46%; 当光照度小于1500 lx时,随着光照度的减小,蓝藻上浮性能逐渐减弱; 当光照度大于6000 lx时,随着光照度的增大,蓝藻上浮性能逐渐减弱. 原因是,在光线很弱时,蓝藻的生长趋于停滞,不再组装新的气囊,而老化的气囊在不断破裂,使得上浮速度降低; 在光线很强时,光合作用合成了大量的淀粉储存在细胞内,增加了藻细胞的重量,同时,合成物质增加了胞内渗透压,加速了气囊的破裂,浮力减小,从而使蓝藻上浮速度降低,当藻颗粒重力与浮力平衡时便不再上浮.
 | 图 3 不同光照度下蓝藻上浮速度分布
Fig. 3 Distribution of the floating velocity of the Cyanobacteria under different illuminance
|
 | 图 4 光照度与蓝藻上浮速度关系图
Fig. 4 Relationship of the illuminance and the floating velocity of the Cyanobacteria
|
共进行了7种不同压强下蓝藻运动特性的测定. 不同压强条件下,蓝藻表现出不同的运动特性. 在0、 0.1 MPa两种压强下,蓝藻明显呈上浮趋势; 0.2 MPa压强下,有少量蓝藻漂浮在顶端,中间各取样口蓝藻质量浓度与初始值差别不大; 0.3 MPa压强下,顶端与底端蓝藻质量浓度均增加,中间各取样口蓝藻质量浓度比初始值稍小,说明在此压强下部分蓝藻开始下沉,部分上浮,而大部分蓝藻既不上浮也不下沉,悬浮于水中; 当压强达到0.4 MPa以上时,蓝藻表现出很强的沉降性能,顶端蓝藻质量浓度减小,最底端蓝藻质量浓度明显增大. 绘制不同压强下蓝藻运动速度分布曲线,如图 5所示. 由于0.2~0.3 MPa压强下蓝藻上浮/下沉趋势不明显,故未绘制速度曲线. 图 6为蓝藻运动速度与压强的关系图,从中可以看出,0~0.1 MPa下,随着压强的增大,蓝藻上浮速度基本不变; 0.4~0.6 MPa下,随着压强的增大,蓝藻下沉速度增大,压强为0.6 MPa时,下沉速度大于0.5、 1.0、 1.5、 2.0 cm ·min-1的蓝藻分别占总量的64.5%、 52.5%、 41.5%、 31.4%.
 | 图 5 不同压强下蓝藻运动速度分布
Fig. 5 Distribution of the motion veolocity of the Cyanobacteria under different external pressure
|
 | 图 6 压强与蓝藻运动速度关系
Fig. 6 Relationship of the external pressure and the motion velocity of the Cyanobacteria
|
 | 图 7 藻细胞场发射超薄切片电镜扫描 Fig. 7 Algal cell field emission scanning electron microscopy of ultrathin sections (a)未加压藻细胞; (b)0.6 MPa加压藻细胞 |
共进行了4种不同温度下蓝藻运动特性的测定,在4种温度下,蓝藻都是上浮的,各个温度下蓝藻上浮速度分布曲线如图 8所示. 图 9为蓝藻上浮速度与温度的关系. 随着温度的降低,蓝藻上浮性能有所下降,可能是由于随着温度的降低,藻细胞内气囊体积收缩,浮力减小. 此外,随着水温降低水体黏度增大,从而增加了藻颗粒上浮的阻力.
 | 图 8 不同温度下蓝藻上浮速度分布
Fig. 8 Distribution of the floating velocity of the Cyanobacteria under different temperature
|
 | 图 9 温度与蓝藻上浮速度关系图
Fig. 9 Relationship of the temperature and the floating velocity of the Cyanobacteria
|
综合条件下蓝藻运动速度分布如图 10所示. 0~30 m水深条件下,在低压、 适宜光照度、 高温等有利于蓝藻上浮的因素作用下,蓝藻呈上浮运动状态,且随着水深的增加,其上浮速度变慢; 当水深达到40 m时,顶端与底端蓝藻质量浓度均增加,中间各取样口蓝藻质量浓度比初始值稍小,说明在此压力下部分蓝藻开始下沉,部分上浮,而大部分蓝藻既不上浮也不下沉,悬浮于水中; 当水深达到50~60 m时,在高压、 无光、 低温等不利于蓝藻上浮的因素作用下,蓝藻表现出下沉运动特性,且随着水深的增加,其下沉速度变大. 由于40 m水深条件下蓝藻上浮或下沉趋势不明显,故未绘制速度分布.
 | 图 10 不同水深下蓝藻运动速度分布
Fig. 10 Distribution of the motion velocity of the Cyanobacteria in different depth of the water
|
由于本实验周期较长,2.1.1~2.1.4节分别采用的是不同批次的实验原水和蓝藻,而同节实验采用是相同批次的蓝藻. 由于不同时期蓝藻生长环境和生理阶段的变化,导致蓝藻的生理特性的差异,因而不同批次实验测得的蓝藻上浮速度有较大差异. 但同一实验条件下的藻类是相同的,具有较好的可比性.
表 2中列出了综合水深条件下,0 MPa和0.6 MPa两种情况下实验结束时各个取样口蓝藻的峰值粒径和对应运动速度. 从中可以看出,粒径大的蓝藻上浮或下沉速度很快,主要是因为藻颗粒粒径越大,其比表面积越小,在水中作上浮或下沉运动时所受到的阻力越小.
蓝藻颗粒是由数百个藻细胞通过胶质鞘粘连在一起的团体,就一个藻细胞而言,由细胞壁、 原生质体组成. 蓝藻细胞壁由内外两层组成,内层主要成分为肽聚糖,薄而坚固,外层为胶质鞘,主要成分为果胶质; 蓝藻原生质体的核区有许多裸露的环状DNA分子,核区周围的周质中存在光合产物,主要包括蓝藻淀粉(Cyanophycean starch)、 脂质颗粒(Lipid granule)等. 蓝藻碳水化合物的密度约为1500 kg ·m-3,核酸的密度约为1700 kg ·m-3,脂类物质的密度约为860 kg ·m-3(只占细胞干重约10%),因而在无气囊时蓝藻颗粒的密度大于水,在有气囊时蓝藻颗粒的密度小于水[25].
直接测定藻细胞和藻颗粒的密度难度较大,本研究从藻颗粒在水中的运动速度的角度,反推藻颗粒的表观密度,采用的方法包括Fluent水力模拟和Stokes公式计算法.
假设蓝藻颗粒为球形,利用Fluent模拟软件模拟粒径为d、 上浮速度为v藻颗粒的密度ρ的方法是,设一段垂直的水流通道,在初始状态,藻颗粒均匀分布于水流通道中. 此后,不含藻的水流自上向下恒定流动,流动速度为v. 在Fluent模拟过程中,只设定固相(藻颗粒)的密度ρ和粒径d,而不能设定颗粒的速度,采用两相流二维模型模拟稳态运行后流道中藻类的分布. 若设定的藻颗粒密度较大,藻类将随水流向下流动,稳定运行后流道中将不再有藻颗粒,反之亦然. 不断改变藻颗粒密度ρ值,直到运行后藻颗粒仍然均匀分布于流道中,说明密度为ρ和粒径为d的藻颗粒上浮速度刚好为v,与自上向下的水流速度v相抵消,藻颗粒没有发生竖向位移. 此时的密度ρ值即为藻颗粒的密度.
采用相似的方法,设定流道中水流自下向上,可以模拟粒径为d、 下沉速度为v藻颗粒的密度ρ. 模拟计算的结果如表 2所示.
按照Stokes公式,藻颗粒的运动速度与密度的关系为:
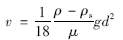
由此可推算上浮藻类的密度为:
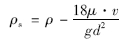
下沉藻类的密度为:
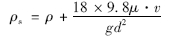
式中,ρ为水的密度,kg ·m-3; ρs为藻颗粒的密度,kg ·m-3; μ为水的动力黏度,Pa ·s.
利用Stokes公式计算的藻颗粒密度如表 2所示. 从中可见,采用Fluent模拟软件模拟的藻颗粒密度与Stokes公式计算值接近.
| 表 2 不同取样口藻类峰值粒径及藻密度 Table 2 Peak value of algae diameter and its density |
通过Stokes公式计算与Fluent模拟两种方法得出的蓝藻密度值显示,常压下,大颗粒蓝藻很快漂浮到顶部,蓝藻粒径越大,上浮速度越快,密度越轻; 高压下,大颗粒蓝藻很快沉淀到底部,蓝藻粒径越大,下沉速度越快,密度越大.
(1)蓝藻在光照度1500~6000 lx范围内均表现出很强的上浮性能,光照度为1500 lx时,上浮速度大于0.4、 0.8、 1.2 cm ·min-1的蓝藻分别占总量的72.5%、 58%、 46%; 当光照度小于1500 lx时,随着光照度的减弱,蓝藻上浮性能逐渐减弱; 当光照度大于6000 lx时,随着光照度的增大,蓝藻上浮性能逐渐减弱.
(2)在8~25℃环境下,蓝藻呈上浮运动,且随着温度的升高,上浮速度增加.
(3)随着压强的增大,蓝藻由上浮转变为下沉. 0~0.1 MPa压强条件下蓝藻上浮,0.2~0.3 MPa压强时大部分蓝藻悬浮于水中,只有少部分上浮或下沉; 压强0.4~0.6 MPa时,蓝藻明显下沉,且随着压强的增加下沉速度加快,压强为0.6 MPa时,下沉速度大于0.5、 1.0、 1.5、 2.0 cm ·min-1的蓝藻分别占总量的64.5%、 52.5%、 41.5%、 31.4%. 常压下,蓝藻粒径越大,上浮速度越快,密度越轻; 高压下,蓝藻细胞内气囊破裂,浮力减小,蓝藻粒径越大,下沉速度越快.
(4)下层水体中压力高、 温度低、 无光照,蓝藻上浮受到抑制,甚至下沉,利用混合技术将蓝藻混合到到下层水体中,有利于控制蓝藻水华.
| [1] | 黄廷林, 丛海兵, 柴蓓蓓. 饮用水水源水质污染控制[M]. 北京: 中国建筑工业出版社, 2009. 208-309. |
| [2] | 吴宇. 富营养化: 中国湖泊面临的治理难题[J]. 生态经济, 2008, (9): 14-19. |
| [3] | 彭芳, 黄廷林, 张延风, 等. 水源水库中藻类生长及分布特征[J]. 供水技术, 2009, 3 (1): 1-4. |
| [4] | 朱广伟. 太湖富营养化现状及原因分析[J]. 湖泊科学, 2008, 20 (1): 21-26. |
| [5] | 刘昂俊, 余华章, 赵孟绪, 等. 汤溪水库富营养化特征与水质管理对策[J]. 生态科学, 2006, 25 (3): 193-197. |
| [6] | 成慧敏, 邱保胜. 蓝藻的伪空泡及其对蓝藻在水体中垂直分布的调节[J]. 植物生理学通讯, 2006, 42 (5): 974-980. |
| [7] | 张海春, 陈初雪, 李春杰. 光照度对蓝藻垂直迁移特性影响研究[J]. 环境污染与防治, 2010, 32 (5): 64-67. |
| [8] | 唐忠波. 蓝藻垂直迁移及沉积物氮磷释放对其浮力调节影响研究[D]. 长沙: 湖南农业大学, 2008. 5-18. |
| [9] | 黄钰铃, 朱纯, 周召红, 等. 不同光照下微囊藻垂直迁移模拟研究[J]. 环境科学与技术, 2012, 35 (11): 21-25. |
| [10] | 周真明, 黄廷林, 丛海兵. 于桥水库中铜绿微囊藻生长特性与上浮性能试验研究[J]. 供水技术, 2007, 1 (2): 14-17. |
| [11] | 段婷婷. 水源水中藻类监测及水质变化原因分析[D]. 西安: 西安建筑科技大学, 2009. 35-36. |
| [12] | 田泽斌, 刘德富, 杨正健, 等. 三峡水库香溪河库湾夏季蓝藻水华成因研究[J]. 中国环境科学, 2012, 32 (11): 2083-2089. |
| [13] | 李凤彬. 洋河水库热成层对水质的影响[J]. 给水排水, 1999, 25 (9): 19-21. |
| [14] | 王晓东, 陈雪初, 黄莹莹, 等. 泽雅水库热成层形成过程及其对水体溶解氧的影响[J]. 给水排水, 2008, 34 (3): 37-38. |
| [15] | Visser P M, Ketelaars H A M, Mur L R. Reduced growth of the Cyanobacterium microcystis in an artificially mixed lake and reservoir [J]. Water Science and Technology, 1995, 32 (4): 53-54. |
| [16] | Simmons J. Algal control and destratification at Hanningfield Reservoir [J]. Water Science and Technology, 1998, 37 (2): 309-316. |
| [17] | 丛海兵, 黄廷林, 赵建伟, 等. 扬水曝气技术在水源水质改善中的应用[J]. 环境污染与防治, 2006, 28 (3): 215-218. |
| [18] | 丛海兵, 黄廷林, 周真明. 于桥水库铜绿微囊藻上浮运动规律及其控制[J]. 给水排水, 2009, 35 (11): 140-145. |
| [19] | 马越, 黄廷林, 丛海兵, 等. 扬水曝气技术在河道型深水水库水质原位修复中的应用[J]. 给水排水, 2012, 38 (4): 7-12. |
| [20] | 丛海兵, 黄廷林, 缪晶广, 等. 水体修复装置——扬水曝气器的开发[J]. 中国给水排水, 2005, 21 (3): 41-45. |
| [21] | 丛海兵. 扬水曝气水源水质改善技术研究[D]. 西安: 西安建筑科技大学, 2007. 37-78. |
| [22] | 陈雪初, 孙扬才, 张海春, 等. 遮光法控藻的中试研究[J]. 环境科学学报, 2007, 27 (11): 1830-1834. |
| [23] | 金相灿, 储昭升, 杨波, 等. 温度对水华微囊藻及孟氏浮游蓝丝藻生长、光合作用及浮力变化的影响[J]. 环境科学学报, 2008, 28 (1): 50-55. |
| [24] | 国家环境保护总局. 水和废水监测分析方法 [M]. (第四版). 北京: 中国环境科学出版社, 2002. 670-671. |
| [25] | 刘健康. 高级水生生物学[M]. 北京: 科学出版社, 2002. 1-197. |