 2014, Vol. 35
2014, Vol. 35



2. 中国科学院上海高等研究院,上海 201210
2. Shanghai Advanced Research Institute, Chinese Academy of Sciences, Shanghai 201210, China
小球藻(Chlorella)属于绿藻门小球藻属,是一种单细胞淡水藻,其所含氨基酸、 维生素、 多糖、 糖蛋白,使其成为一种很有价值的医药制品[1,2,3].另外,小球藻可以耐受高盐、 高渗透压及高浓度抗生素,这些特性使小球藻在污水处理领域也有很好的应用[4,4].因此,小球藻是极具应用价值的重要生物资源.
微藻因其光合效率高、 生长速度快、 油脂含量高[6, 7],可以利用空气中的CO2降低温室效应等优势,被认为是制备生物柴油的理想原料.研究表明许多微藻在不同营养胁迫的条件下能积累更高含量的油脂.如氮源缺乏可以提高普通小球藻(Chlorella vulgaris)和微拟球藻(Nannochloropsis)的油脂含量[8, 9]; 磷限制会导致凯氏拟小球藻(Parachlorella kessleri)和微拟球藻的油脂含量增加[10, 11]; 缺硫条件下,小球藻和莱茵衣藻(Chlamydomonas reinhardtii)细胞内总脂的含量将会增加[12, 13].王秀锦等[14]通过对蛋白核小球藻进行光异养培养条件优化,使其油脂产量达到了121.3 mg ·(L ·d)-1.
镁是微藻生长必需的营养元素之一,对维持叶绿体结构和功能具有重要作用,同时适宜的镁离子浓度可以提高光系统活性和光能转化效率.Huang 等[15]研究证实在小球藻异养过程中,缺镁胁迫使油脂含量达到干重的46.90%.但小球藻在自养条件下,缺镁胁迫是否会提高油脂含量还鲜见报道.本研究主要是以普通小球藻(Chlorella vulgaris)为对象,在光合自养培养过程中,考察缺镁胁迫对普通小球藻光合生理和油脂含量的影响,以期为进一步研究微藻油脂合成相关机制提供依据.
本实验所用藻种为普通小球藻(Chlorella vulgaris FACHB-25),购自中国科学院水生生物研究所淡水藻种库.
1.2 培养条件普通小球藻光自养生长培养基分别为TAP[16]和不含Mg2+的TAP(以下简写为TAP-Mg).首先将小球藻接种至含TAP的培养液中正常培养至对数生长期D680为0.8~0.9,经离心弃上清后,使用TAP-Mg培养液对藻细胞清洗两遍后,分别接入含300 mL TAP的培养液中和含300 mL TAP-Mg的培养液中继续培养6 d.各设置3个平行样,培养温度为25℃,光强为50 μmol ·(m2 ·s)-1,光暗比14 h ∶10 h,每天定时摇动3次.
1.3 测定方法 1.3.1 生物量的测定通过重量法测定生物量来监测细胞的生长.先将Whatman滤纸(GF/F,0.7 μm,47 mm)在105℃烘箱内烘干12 h,取3 mL藻液过膜抽滤后,将带有藻体的滤纸放入105℃烘箱内烘干至恒重.利用公式计算出单位体积藻液的干重,W1为滤膜干重,W2为抽滤藻液后滤膜干重,W为单位体积藻细胞干重(g ·L-1).
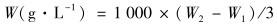
使用BCA法测定普通小球藻中蛋白含量.取5 mL藻液,离心收集藻细胞,依次加入 1 mL KH2PO4 (M/15)pH 4.5 和2 mL 20% NaOH,混匀后,100℃沸水浴5 min[17].离心后,取上清液,使用美国Pierce公司生产的BCA蛋白分析试剂盒对所抽提出的蛋白进行浓度测定,以牛血清白蛋白为标品,制作标准曲线.使用的检测仪器是美国BioTek公司生产的Synergy H1全功能微孔板检测仪,设定吸收光波长为560 nm.
1.3.3 中性脂的测定方法微藻中的中性脂与尼罗红荧光染色溶液结合能发出荧光检测信号,脂质的量与荧光强度呈正比.因此,可以快速、 准确地比较活体微藻细胞内中性脂的相对含量[18~20].取1 mL藻液,加入20 μL DMSO,20 μL 尼罗红溶液(0.5 mg ·mL-1丙酮),黑暗中放置10 min,然后使用美国BioTek公司生产的Synergy H1全功能微孔板检测仪检测激发波长488 nm和发射波长554 nm下的荧光值来比较不同样品中性脂的相对含量.
1.3.4 总脂的测定总脂的测定按照文献[21]的方法进行.取一定量冻干的藻粉,加入5 mL氯仿/甲醇(2 ∶1),置于35℃水浴振荡1 h后,离心,取出上清保存,继续向残存的藻体中加入5 mL氯仿/甲醇(2 ∶1),置于35℃水浴振荡1 h后,离心,取出上清.将两次的上清液混合后,加入3 mL蒸馏水,离心后,油脂留在下层液体中,取出下层液体,使用氮吹仪将有机溶剂吹干,称重并计算油脂含量.其中W0为空瓶重,W1为冻干的藻粉质量,W2为装有油脂的瓶重.
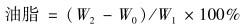
叶绿素含量的测定采用的是分光光度法.取5 mL藻液,离心弃上清后,加入95%丙酮至10 mL处,振荡后放在暗处过夜.离心取上清,使用分光光度计测定其在645 nm、 663 nm波长下的吸光值.根据Arnon公式[22]分别为:
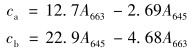
使用德国Walz公司生产的Water-PAM水样叶绿素荧光仪进行叶绿素荧光参数的测定.向比色杯中依次加入3 mL蒸馏水、 15 μL藻液,混匀后,将样品暗适应10 min.叶绿素荧光参数PSⅡ的最大光能转化效率Fv/Fm和非光化学淬灭qN可在荧光仪上直接读出.
1.4 数据处理 采用SPSS 19.0软件对实验数据进行单因素方差分析.正常培养(TAP)和缺镁胁迫条件下(TAP-Mg)普通小球藻的生长状况见图 1.随着培养时间的延长,正常培养条件下的小球藻的生物量呈不断上升趋势,前3 d生物量增加较快,生长快速,细胞密度达到一定程度后,依然进行缓慢的增长,培养结束时的生物量为接种时的2.37倍,达到2.13 g ·L-1.而在缺镁胁迫后的前3 d,普通小球藻生长速度也较快,其中前2 d与正常培养无显著差异(P>0.05),而从第3 d开始生长显著低于正常培养条件(P<0.05),缺镁胁迫后的第3 d开始,普通小球藻的生物量基本不再增加,生长近乎停滞,至第6 d时生物量为1.61 g ·L-1,相比于正常培养下降了20%.以上结果表明,镁离子缺乏直接影响了普通小球藻的培养周期和生物量.
 | 图 1 正常培养和缺镁胁迫条件下普通小球藻的生长状况 Fig. 1 Growth of Chlorella vulgaris under normal condition and magnesium deficiency stress |
正常培养(TAP)和缺镁胁迫条件下(TAP-Mg)普通小球藻中蛋白含量的变化见图 2.从中可知,正常培养条件下普通小球藻的蛋白含量在接种至培养结束这段时间内无显著变化.而缺镁胁迫条件下普通小球藻的蛋白含量从第2 d开始显著降低(P<0.05).开始缺镁胁迫培养时其含量为78.74 mg ·g-1,而缺镁培养2 d后,其蛋白质含量为53.04 mg ·g-1,仅为刚开始培养的67.36%.随着缺镁培养时间的延长,蛋白质含量也在不断的降低,培养结束时其蛋白含量为40.03 mg ·g-1,与正常培养相比其蛋白质含量降低了43.96%.
 | 图 2 正常培养和缺镁胁迫条件下普通小球藻的蛋白含量变化Fig. 2 Protein content of Chlorella vulgaris under normal condition and magnesium deficiency stress |
本实验通过将处于对数生长后期的普通小球藻经6000 r ·min-1离心7 min后,藻细胞重悬于TAP和缺镁的TAP-Mg培养基中,分别培养至6 d,再分别用重量法和尼罗红染色法测定普通小球藻内总脂及中性脂含量的变化(见图 3).结果表明,正常培养条件下,普通小球藻的总脂含量为26.89%,而缺镁胁迫条件下,普通小球藻的总脂含量为32.16%,相比于正常培养提高了19.60%.缺镁胁迫条件下,普通小球藻尼罗红染色后检测到的中性脂荧光信号要显著高于正常培养条件下的普通小球藻.
 | 图 3 正常培养和缺镁胁迫条件下普通小球藻的总脂和中性脂含量Fig. 3 Total lipid and neutral lipid of Chlorella vulgaris under normal condition and magnesium deficiency stress |
镁是叶绿素的中心分子,对维持叶绿体的结构、 叶绿体的发育有很重要的作用[14].正常培养(TAP)和缺镁胁迫条件下(TAP-Mg)普通小球藻的叶绿素含量如图 4和图 5.从中可知,正常培养条件下普通小球藻的叶绿素a和叶绿素b含量呈螺旋上升趋势,在培养第5 d,叶绿素a和叶绿素b含量达到最高分别为18.21 mg ·g-1和5.81 mg ·g-1.而在缺镁胁迫条件下,叶绿素a和叶绿素b含量在前2 d下降很快,随后几乎没有明显变化,至培养结束时,缺镁胁迫条件下,叶绿素a和叶绿素b含量分别为12.56 mg ·g-1和3.69 mg ·g-1,比正常培养条件下分别下降了约27.52%和28.07%.以上实验结果表明,缺镁胁迫条件下严重影响了普通小球藻色素的合成.
 | 图 4 正常培养和缺镁胁迫条件下普通小球藻叶绿素a的含量变化Fig. 4 Chlorophyll a content of Chlorella vulgaris under normal condition and magnesium deficiency stress |
 | 图 5 正常培养和缺镁胁迫条件下普通小球藻的叶绿素b的含量变化Fig. 5 Chlorophyll b content of Chlorella vulgaris under normal condition and magnesium deficiency stress |
缺镁胁迫条件下和正常培养条件下普通小球藻的叶绿素荧光参数见图 6. Fv/Fm代表了PSⅡ的最大量子产量,它反映了绿色植物的潜在最大光合能力.与正常培养条件相比,缺镁胁迫下普通小球藻的Fv/Fm明显下降,并随着胁迫时间的延长,下降幅度逐步增大,至培养结束时比接种时下降了21.90%.分析结果表明,缺镁胁迫使普通小球藻的PSⅡ受到了损伤,光能转化率明显降低,光合作用受到了抑制.
 | 图 6 正常培养和缺镁胁迫条件下普通小球藻Fv/Fm的变化Fig. 6 The Fv/Fm of Chlorella vulgaris under normal condition and magnesium deficiency stress |
qN指非光化学淬灭,反映了绿色植物吸收的光能不能用于电子传递而以热等形式散耗掉的光能部分,是植物保护PSⅡ的重要机制.接种后,即出现了缺镁胁迫条件下的普通小球藻的qN值显著高于正常培养(见图 7),可见普通小球藻对于缺镁胁迫非常敏感,即刻启动了非光化学保护.至培养的第2 d时,正常培养和缺镁胁迫条件下的普通小球藻的qN 都已经降为0,但随着培养时间的延长,呈现上升趋势,至第4 d达到最大值后就不再上升.并且从第4 d开始缺镁胁迫条件下的普通小球藻的qN值高出对照组57.9%(见图 7).由以上结果可知,普通小球藻在培养期间受到的缺镁胁迫越来越严重,PSⅡ的热耗散能力增强,可以使其有效地避免或减轻因吸收过多的光能而造成的损伤.
 | 图 7 正常培养和缺镁胁迫条件下普通小球藻qN的变化Fig. 7 The qN of Chlorella vulgaris under normal condition and magnesium deficiency stress |
本研究发现,当小球藻受到缺镁胁迫时,生长缓慢并受到抑制,总油脂含量提高,表明镁元素是小球藻生长的限制因子,小球藻在遭受缺镁胁迫时启动了抗逆生理反应,即通过积累油脂来度过不利环境.小球藻细胞开始积累油脂的同时,细胞内的总蛋白、 叶绿素含量下降,这些现象的产生可能是蛋白质合成中氨基酸的活化过程需要镁的参加,镁又是叶绿素a结构的中心原子,缺镁会使小球藻细胞内蛋白质和叶绿素的合成受到影响,进而影响参与光合作用的重要蛋白和酶类,使藻细胞的光合作用受到抑制,因而碳代谢会更多地进入油脂合成的方向.
微藻油脂的生物合成是一个复杂的代谢网络,需要几十种相关酶的共同调控过程.而一些金属离子作为一些重要酶的辅因子,对于高等植物和藻类中调控基因表达起到了非常关键的作用[23].Huang等[15]通过无光照异养条件下金属离子Mg2+、 Ca2+对小球藻的总脂肪酸含量的影响,发现在Mg2+缺失、 100 mg ·L-1的CaCl2 ·2H2 O时都可得到很高的总脂肪酸含量. Wang 等[24]研究表明,20 mmol ·L-1 Mg2+对小球藻胁迫,使其脂肪酸组成发生了变化,饱和脂肪酸减少了14.5%,说明镁离子的存在对脂肪酸的合成有抑制作用.刘志媛等[25]研究发现在高浓度的Fe3+条件下培养海水小球藻,能够明显促进海水小球藻中油脂的积累,出现了两次油脂积累的高峰,并认为是由于己糖库的代谢途径由合成淀粉为主转变为合成油脂.本实验发现小球藻在缺镁胁迫下,油脂总量提高了19.6%,TAG的含量也显著提高了,说明了缺镁可以促进油脂的积累,与以上研究结果相一致.由以上结果,可以推测金属离子改变了藻体细胞油脂代谢网络,并可能影响到了一些关键酶基因的表达.
(1)缺镁胁迫不仅直接影响了小球藻的生长周期,还抑制了蛋白质合成.从胁迫第3 d开始,小球藻的生长近乎停滞; 蛋白含量随胁迫时间延长不断降低,最终仅为初始时的43.96%.
(2)缺镁胁迫与正常培养条件相比,小球藻叶绿素a和b的含量下降27.52%、 28.21%,Fv/Fm下降22.54%,qN上升47.81%,表明缺镁使叶绿素的合成受到限制,光合作用受到了严重抑制,PSⅡ的热耗散能力增强.
(3)缺镁胁迫下,小球藻的油脂总量提高了19.60%,中性脂也显著高于正常条件.这表明缺镁胁迫小球藻,伴随着生长抑制出现,蛋白质合成受阻,碳同化进入脂类合成代谢,促进了小球藻油脂含量的增加.
| [1] | 杨鹭生, 李国平, 陈林水. 蛋白核小球藻粉的蛋白质、氨基酸含量及营养价值评价[J]. 亚热带植物科学, 2003, 32 (1): 36-38. |
| [2] | 王业勤, 李勤生. 小球藻应用研究动态[J]. 微生物学通报, 1985, 12 (6): 275-277. |
| [3] | 陈晓清, 苏育才. 小球藻的应用研究进展[J]. 生物学教学, 2012, 37 (1): 8-9. |
| [4] | Bhatnagar A, Bhatnagar M, Chinnasamy S, et al. Chlorella minutissima—A promising fuel alga for cultivation in municipal wastewaters[J]. Applied Biochemistry and Biotechnology, 2010, 161 (1-8): 523-536. |
| [5] | Aslan S, Kapdan I K. Batch kinetics of nitrogen and phosphorus removal from synthetic wastewater by algae [J]. Ecological Engineering, 2006, 28 (1): 64-70. |
| [6] | Li Q, Du W, Liu D H. Perspectives of microbial oils for biodiesel production[J]. Applied Microbiology and Biotechnology, 2008, 80 (5): 749-756. |
| [7] | Li P L, Miao X L, Li R X, et al. In situ biodiesel production from fast-growing and high oil content Chlorella pyrenoidosa in rice straw hydrolysate[J]. Journal of Biomedicine and Biotechnology, 2011, 2011 : 1-8. |
| [8] | Mujtaba G, Choi W, Lee C G, et al. Lipid production by Chlorella vulgaris after a shift from nutrient-rich to nitrogen starvation conditions[J]. Bioresource Technology, 2012, 123 : 279-283. |
| [9] | 王辉, 陈来喜, 肖爱凤, 等. 氮和铁对小球藻和微拟球藻油脂积累的影响[J]. 海南师范大学学报, 2012, 25 (1): 86-89. |
| [10] | Li X L, Pribyl P, Bisova K, et al. The microalga Parachlorella kessleri-a novel highly efficient lipid producer[J]. Biotechnology and Bioengineering, 2013, 110 (1): 97-107. |
| [11] | Rodolfi L, Zittelli G C, Bassi N, et al. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor[J]. Biotechnology and Bioengineering, 2009, 102 (1): 100-112. |
| [12] | Mizuno Y, Sato A, Watanabe K, et al. Sequential accumulation of starch and lipid induced by sulfur deficiency in Chlorella and Parachlorella species[J]. Bioresource Technology, 2013, 129 : 150-155. |
| [13] | Cakmak T, Angun P, Demiray Y E, et al. Differential effects of nitrogen and sulfur deprivation on growth and biodiesel feedstock production of Chlamydomonas reinhardtii[J]. Biotechnology and Bioengineering, 2012, 109 (8): 1947-1957. |
| [14] | 王秀锦, 李兆胜, 刑冠岚, 等. 蛋白核小球藻Chlorella pyrenoidosa-15的异养培养条件优化及污水养殖[J]. 环境科学, 2012, 33 (8): 2735-2740. |
| [15] | Huang G, Chen F. Effect of metal ions on biomass and total fatty acids content of Chlorella vulgaris[J]. Modern Food Science and Technology, 2009, 25 (5): 474-477. |
| [16] | Gorman D S, Levine R P. Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardtii[J]. Proceedings of the National Academy of Sciences of the United States of America, 1965, 54 (6): 1665-1669. |
| [17] | Bellou S, Aggelis G. Biochemical activities in Chlorella sp. and Nannochloropsis salina during lipid and sugar synthesis in a lab-scale open pond simulating reactor[J]. Journal of Biotechnology, 2012, 164 (2): 318-329. |
| [18] | Chen W, Sommerfeld M, Hu Q. Microwave-assisted nile red method for in vivo quantification of neutral lipids in microalgae[J]. Bioresource Technology, 2011, 102 (1): 135-141. |
| [19] | Lee S J, Yoon B D, Oh H M. Rapid method for the determination of lipid from the green alga Botryococcus braunii[J]. Biotechnology Techniques, 1998, 12 (7): 553-556. |
| [20] | Liu Z Y, Wang G C, Zhou B C. Effect of iron on growth and lipid accumulation in Chlorella vulgaris[J]. Bioresource Technology, 2008, 99 (11): 4717-4722. |
| [21] | Folch J, Lees M, Sloane Stanley G H. A simple method for the isolation and purification of total lipides from animal tissues[J]. Journal of Biological Chemistry, 1957, 226 (1): 497-509. |
| [22] | 叶济宇. 关于叶绿素含量测定中的Arnon计算公式[J]. 植物生理学通讯, 1985, (6): 69. |
| [23] | Raman V, Ravi S. Effect of salicylic acid and methyl jasmonate on antioxidant systems of Haematococcus pluvialis[J]. Acta Physiologiae Plantarum, 2011, 33 (3): 1043-1049. |
| [24] | Wang Y, Liu Z Y, Qin S. Effects of iron on fatty acid and astaxanthin accumulation in mixotrophic Chromochloris zofingiensis[J]. Biotechnology Letters, 2013, 35 (3): 351-357. |
| [25] | 刘志媛, 王广策. 铁促进海水小球藻油脂积累的动态过程[J]. 海洋科学, 2008, 32 (11): 56-59. |