 2023, Vol. 44
2023, Vol. 44

2. 湖南省灌溉水源水质污染净化工程技术研究中心, 长沙 410128;
3. 农业农村部南方产地污染防控重点实验室, 长沙 410128
 , LIU Yu-ling1,2,3 , WU De1,2,3 , HUANG Xin-ming1,2,3 , ZHANG Pu-xin1,2,3 , TIE Bo-qing1,2,3
, LIU Yu-ling1,2,3 , WU De1,2,3 , HUANG Xin-ming1,2,3 , ZHANG Pu-xin1,2,3 , TIE Bo-qing1,2,3
2. Hunan Engineering & Technology Research Center for Irrigation Water Purification, Changsha 410128, China;
3. Key Laboratory of Southern Farmland Pollution Prevention and Control, Ministry of Agriculture and Rural Affairs, Changsha 410128, China
农田土壤污染问题关乎国计民生, 一直以来受到社会各界广泛关注, 在“十三五”期间启动关于“农业面源和重金属污染农田综合防治与修复技术研发”的重点专项中, 农田镉(Cd)和砷(As)污染被列为重点研究对象[1].据统计我国所有粮食作物中稻米Cd-As超标率最高, 严重威胁我国的粮食安全[2, 3]. 据2014年中国环境保护部公布的第二次全国土地调查结果显示, Cd-As复合污染造成的耕地问题正在给我国造成越来越大的经济损失[4].Mu等[5]采集了中国4个主要水稻产区19个省份的113个土壤样本, 发现土壤中ω(Cd)和ω(As)的平均值分别为0.45 mg·kg-1和11.80 mg·kg-1, 其中超过国家允许Cd和As含量的土壤分别占比33.6%和6.19%.在“有色金属之乡”湖南, 株洲某县受镉和砷复合污染的耕地面积约有23.23 km2[6].Cd主要以Cd(Ⅱ)阳离子的形式存在, 通过电镀、塑料色素、绘画和电池制造等工业活动进入环境[7].As主要以As(Ⅲ)和As(Ⅴ)的形式存在, 且As(Ⅲ)的毒性强, 通过施用农药、燃烧化石燃料和采矿进入土壤.进入土壤的Cd和As易累积、难降解且造成的污染几乎不可逆转[8], 导致作物产量低和品质差, 经食物链进入人体后危害人体健康[9], Cd会影响肾脏和骨骼, 长期接触导致骨质疏松和脊柱畸形等[10], As在体内长期积累会引起皮肤癌、肝癌和肺癌等[11].重金属复合污染的土壤中各种重金属元素相互作用极其复杂, 且这种作用会因各元素存在浓度的不同而表现出不同的作用形式[12~14].据报道, 目前Cd-As复合污染在不同土壤环境下主要是拮抗和协同作用[15, 16].
近年来, 针对土壤的重金属污染问题, 微生物修复法正逐渐得以运用.现阶段已证实的修复机制包括细胞代谢、吸附、沉淀和氧化还原反应等, 不同类群的微生物对重金属的修复机制不同[17~21].如, 张雅荟等[22]筛选鉴定的菌株Burkholderia sp.Y4, 能够有效定殖在水稻根表和根际土壤中, 通过生长代谢和菌体吸附等途径钝化Cd, 降低水稻籽粒Cd积累量.迄今为止, 国内外学者已经从自然界中分离出许多细菌[23, 24], 包括节杆菌属、微球菌属和假单胞菌属, 这些细菌都对重金属具有良好的耐受性, 大部分细菌主要通过细胞内外吸附固定镉和砷等, 从而增加重金属的生物稳定性[25, 26].而一些具有硫酸盐还原功能的菌株, 则能够将硫酸盐还原形成H2S、HS-和S2-, 进一步与重金属离子发生反应生成难溶性金属硫化物从而达到固定重金属的目的[27, 28].例如, Xia等[29]发现一种硫酸盐还原菌Alishewanella sp.WH16-1, 可生成H2S, 其与Cd反应生成CdS沉淀, 将该细菌用于水稻盆栽试验发现土壤中的生物可利用Cd, 降低糙米中Cd的含量78%; 此外, 该类细菌的硫酸盐还原过程还间接参与了砷的生物地球化学循环[30, 31].通常可生成雌黄(As2S3)、雄黄(AsS)和砷黄铁矿(FeAsS)从而降低了砷的可利用性[32~34].在本研究中, 筛选了1株具有硫酸盐还原功能的肠杆菌, 通过去除率实验结合表征实验, 通过模拟体系从微观角度阐释了该细菌对镉、砷的固定机制, 以期为金属类金属复合污染微生物治理提供一定的修复思路.
1 材料与方法 1.1 菌株筛选及鉴定 1.1.1 菌种来源及培养基菌株筛选供试土壤为湖南省株洲市某Cd和As复合污染农田, 取样土层为10~15 cm耕作层土壤, 其pH为5.42, ω(Cd)和ω(As)平均值分别为23.07 mg·kg-1和44.5 mg·kg-1, 分别超过《土壤环境质量农用地土壤风险管控标准》(GB 15618-2018)中的农用地土壤风险筛选值的76.9倍和1.48倍, 为Cd和As复合污染土壤.菌株的筛选培养及富集培养均采用改进的Postgate培养基[35], 其主要成分如下:NH4Cl 0.5 g·L-1、CaCl2 0.1 g·L-1、Na2SO4 1.0 g·L-1、K2HPO4 0.5 g·L-1、MgSO4 2.0 g·L-1、酵母膏1.0 g·L-1、乳酸钠3.5 mL·L-1、L-半胱氨酸盐酸盐0.5 g·L-1、抗坏血酸0.1 g·L-1、FeSO4(指示剂)0.5 g·L-1, pH调至7.0±0.2.在以上培养基中加2%琼脂即为Postgate固体培养基.
1.1.2 菌株筛选菌株筛选参照万海清等[36]改进的一种筛选方法.首先, 按1g ∶ 10 mL将土样接入无菌水制成土壤菌悬液, 充分振荡后取10 mL接入选择培养基中, 充入N2在30 ℃厌氧培养.培养3~4 d后接入液体分离富集培养基进行厌氧培养, 待液体变成墨汁色(FeS), 瓶口有臭鸡蛋味(H2S), 即可采用稀释涂布-夹层培养法进行厌氧分离培养.培养5~6 d后平板中长出许多黑色球状小菌落, 将这些菌落连同琼脂块切割、适当捣烂分散移入液体培养基.重复进行稀释涂布、夹层培养和挑选等2次, 可达到对菌株的分离纯化, 获得可以鉴定、保存和转接的纯菌株.
1.1.3 菌株鉴定将筛选出的菌株培养至对数生长期, 送至湖南擎科生物技术有限公司进行扩增、测序.菌株的16S rRNA基因使用通用引物27F(5′-AGAGTTTG ATCCTGGCTCAG-3′)和1492R(5′-TACGGCTAC CTTGTTACGACTT-3′) 通过PCR扩增.PCR反应程序为:98℃ 30 s; 30次循环(98℃ 10 s, 55℃ 30 s, 72℃ 1 min); 72℃ 10 min.PCR产物纯化后测序.根据测序结果在NCBI数据库进行同源性比较分析, 确定亲缘关系, 申请基因登录号.使用MEGA6.5软件, 参考Tamura等[37]的方法采用Neighbor-Joining算法构建16S rRNA基因序列的系统发育树, 自展次数设定为1 000, 最终确定目标菌株的系统发育学地位.
1.1.4 菌株硫酸盐还原功能及镉、砷耐性(1) 菌株硫酸盐还原功能将目标菌株活化后, 以2%的接种量接种至含有0.5 g·L-1FeSO4(指示剂)的液体培养基中并在手套箱中操作将锥形瓶充入N2, 于27~30℃、120 r·min-1的振荡箱中培养, 观察培养基是否变为墨汁色, 瓶口是否有臭鸡蛋气味.设置3个平行.
(2) 不同镉、砷浓度下菌株生长曲线将目标菌株活化后, 以2%的接种量接种至含有不同镉、砷浓度(使用1 000 mg·L-1的CdCl2和Na3AsO4母液稀释)的液体培养基中并在手套箱中操作将锥形瓶充入N2, 于27~30℃、120r·min-1的振荡箱中培养.镉、砷浓度设置为0、0.2、0.5和1 mmol·L-1.在0~126 h内, 每2 h或6 h取一次样, 于600 nm的分光光度计下测定不同浓度下的生长曲线.设置3个平行.
1.2 Cd和As去除率实验及样品表征 1.2.1 菌株对Cd和As去除率实验用去离子水配制1 000 mg·L-1的CdCl2和Na3AsO4母液.将保存在平板培养基中的菌株切块捣碎移至液体培养基中, 活化48 h后, 以2%的接种量接种至含有Cd、As和Cd及As复合(母液提前调节pH至5.5~5.6)的液体培养基中, 充入N2, 在25~27℃振荡箱中培养48 h.培养后, 将反应体系于离心机中以5 000 r·min-1的速度离心5 min, 收集上层液体测定镉、砷浓度.镉、砷浓度的测定使用电感耦合等离子体发射光谱(ICP-OES, PerkinElmer Optima 8300).无论在单一还是复合体系中, Cd和As的浓度均设置为0、0.01、0.05、0.1和0.2 mmol·L-1.每个浓度均设置一组对照, 3组平行, 对照组不接种细菌.Cd和As去除率由以下公式计算:
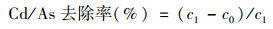
|
式中, c1为相应浓度中对照组金属浓度, c0为相应浓度中实验组金属浓度.
1.2.2 样品表征(SEM-EDS、XRD和XPS)首先, 将参与固定Cd和As后的样品先用0.2 mol·L-1的磷酸缓冲盐溶液(PBS缓冲液)冲洗3次, 再于25%戊二醛中固定过夜并离心收集菌体, 配制一系列浓度梯度的乙醇进行脱水(30%、50%、70%、80%、90%和100%), 最后于-53℃的冷冻干燥机中干燥并研磨成粉末.使用场发射扫描电镜(SEM)-形貌(FEI Inspect F50, 美国)观察菌株参与反应前后细胞形貌, 使用场发射扫描电镜(SEM)-点扫EDS(EDAX super octane, 美国)测定元素组成.
将反应体系于离心机中以5 000 r·min-1的速度离心5 min, 收集下层反应底物于-53℃的冷冻干燥机中干燥并研磨成粉末.使用Bruker D8 Advance-广角XRD(Bruker D8 Advance)测定反应底物中的物质组成, 使用Jade6软件进行物质比对.使用KRATOS Axis Ultra X射线光电子能谱仪(Thermo Fisher Scientific, 美国)对参与固定镉、砷前后的样品进行分析.使用C1s峰(284.8 eV)校准结合能.用软件包XPSPEAK 41进行数据处理, 背景信号用Shirley方法扣除, 采用Lorentzian-Gaussian方法进行分峰拟合.
2 结果与分析 2.1 菌株鉴定 2.1.1 菌株形态及菌种鉴定从镉、砷复合污染土壤中成功分离1株细菌命名为M5, 图 1(a)为菌株在Postgate固体培养基中生长72 h左右的菌落形态.单菌落呈白色、圆形且边缘较光滑.目前保藏于中国微生物菌种保藏管理委员会普通微生物中心, 保藏号为GCMCC NO.22636.基因测序表明, 该菌株的16S rRNA基因(登录号为MT641242)与菌株Enterobacter_strain_FC18569(登录号为MK577384.1)具有99.45% 的同源性.系统发育分析表明[图 1(b)], 该菌株与Enterobacter_strain_fC18569菌株可以形成一个簇, 并与其他菌株区分开来.因此, 菌株M5被鉴定为肠杆菌属(Enterobacter sp.).

|
图 1 菌株M5的菌落形态及其16S rRNA基因序列的系统进化树 Fig. 1 Colony morphology and phylogenetic tree of 16S rRNA of strain M5 |
在液体培养基中添加FeSO4(指示剂), 接种细菌培养后, 培养基变为墨汁色(图 2)且瓶口有臭鸡蛋气味, 且在3个平行中均产生以上现象, 这表明该菌的硫酸盐还原功能并非偶然.原因在于培养基中的硫酸根被还原为S2-进而分别与Fe2+和H+结合生成黑色沉淀FeS及臭鸡蛋气味的H2S.以上现象验证了目标菌株具有硫酸盐还原功能.

|
图 2 墨汁色的培养基 Fig. 2 Ink-colored medium |
图 3为不同Cd和As浓度下肠杆菌M5的生长曲线.在纯培养基中, 菌株M5大约在60 h达到对数生长期, 最大生长量时D600为1.4~1.5. Cd和As的添加对其对数生长期的发生时间没有明显影响, 但不同程度抑制菌株生长, 且Cd和As浓度越高对其生长抑制程度越强, 当Cd和As浓度达到1 mmol·L-1时, 对M5的生长有明显抑制, 生长量降低了约68%.但在Cd和As浓度均为0.2 mmol·L-1和0.5 mmol·L-1时, 菌株生长稳定后D600值在1.0~1.4之间, 生长状况良好, 表明肠杆菌M5对Cd和As具有一定耐受性.

|
图 3 不同Cd、As浓度下Enterobacter sp. M5的生长曲线 Fig. 3 Growth curve of Enterobacter sp. M5 under different Cd and As concentrations |
由图 4可知, 在单一体系中, 当c(Cd)为0.1 mmol·L-1时, 去除率最大达94.13%, 而当c(Cd)达到0.2 mmol·L-1时, 去除率最低为80.12%; 相比之下, 菌株对As的去除率远低于对Cd的, 当c(As)为0.01 mmol·L-1时, 去除率最大为27.26%, 在其他3个浓度中, 去除率差异不明显, 范围在19.96% ~22.84%.在复合体系中, 由于As的添加, 菌株对Cd的去除率有不同幅度的增加, 其中浓度为0.1 mmol·L-1时促进程度最大, 菌株对Cd的去除率相较于单一体系增加了15.50%; 然而, Cd的添加对As的去除率影响不显著.

|
图 4 菌株M5对Cd(Ⅱ)和As(Ⅴ)去除率 Fig. 4 Cd(Ⅱ)/As(Ⅴ) removal efficiency of the M5 strain |
(1) SEM-EDS和XRD分析电镜图片显示, 纯细菌M5形态为杆状[图 5(a)], 表面较为光滑, 长约1.5 μm, 直径约为0.6 μm.而参与固定Cd和As后, 细菌的细胞形态未发生明显变化, 而是在细胞表面出现颗粒物质, 推测为Cd和As被固定产生的硫化物沉淀[图 5(b)].结合EDS结果, 在纯细菌中, 细菌表面主要由C、N、O、Na、Mg、P、S和Cl等元素组成, 固定Cd和As后, 细菌表面主要组成元素增加Cd和As, 其中Cd和As所占质量分数分别为1.64%和0.80%, 所占原子分数分别为0.31%和0.18%, 这一结果表明部分镉、砷已经被细菌固定.通过菌株参与固定Cd和As的反应物的XRD分析可以确定(图 6), 通过标准光谱比对, 发现存在CdS(JCPDS卡片编号43-0985:CdS)和As2S3(JCPDS卡片编号71-2435:As2S3).综合以上表征结果, 可以确定菌株M5可以将Cd和As固定为CdS和As2S3.

|
图 5 菌株M5固定Cd和As前后的扫描电镜(SEM)和能量色散X射线光谱(EDS) Fig. 5 Scanning electron microscopy (SEM) and energy dispersive X-ray spectroscopy (EDS) of strain M5 before and after fixing Cd and As |

|
图 6 反应底物的X射线衍射 Fig. 6 X-ray powder diffraction (XRD) of the precipitates |
(2) XPS分析图 7是肠杆菌M5参与固定Cd和As前后的C 1s高分辨光谱.针对肠杆菌, 将其碳峰分解为3个部分:288 eV处的峰归属于碳与氧形成单键或双键(C=O和C—O—C), 主要包括羧酸盐; 286 eV处的峰归属于C—(O, H); 285 eV处归属于C—(O, N), 这两部分主要包括醚、醇、胺和酰胺[38, 39].分析C 1s光谱发现, 参与固定Cd和As前后的肠杆菌存在明显差异, 即固定金属后的肠杆菌在288 eV和286 eV处的峰变宽且峰面积增大.在纯肠杆菌中, C=O和C—O—C所占面积比例为6.04%, C—(O, N)所占面积比为28.03%, 在参与固定Cd和As后这两种组分分别增加了3.22%和3.74%.这一结果表明金属离子与微生物的羧基、羟基和酰胺基的氢原子之间的离子交换主要参与生物吸附.

|
图 7 肠杆菌M5固定Cd和As前后的高分辨率C1s XPS光谱 Fig. 7 High-resolution C1s XPS spectra of M5 before and after the sorption of Cd(Ⅱ)/As(Ⅴ) |
在本研究中, 从镉、砷复合污染农田土壤中筛选出1株肠杆菌M5(Enterobacter sp.), 在含有指示剂的培养基中生长可使指示剂显色(FeS)且瓶口有臭鸡蛋(H2S)气味, 表明该菌具有硫酸盐还原功能.Hoffmann等[40]于2005年首次发现并报道了该种细菌, 近年来, 其硫酸盐还原功能也逐渐被学者们广泛报道.例如, Wang等[41]将肠杆菌A11(Enterobacter sp.)和毛单胞菌A23(Comamonas sp.)与Cd共培养发现可产生H2S和琥珀酸等, 并且Cd被高效固定形成CdS.同样地, 在本研究中发现当Cd浓度为0.1 mmol·L-1时, 去除率最大达94.13%, 当As(Ⅴ)浓度为0.01 mmol·L-1时, 去除率最大为27.26%, 且在反应底物的表征中发现了CdS和As2S3. 在厌氧还原条件下, 菌株M5对硫酸盐进行还原形成H2S、HS-和S2-的混合物, 这些物质进一步与镉、砷反应生成金属硫化物.此外, 由于菌株M5可以利用电子供体将As(Ⅴ)还原为As(Ⅲ), 且S2-等物质有较强的还原性也可以将As(Ⅴ)还原为As(Ⅲ), 故最终生成As2S3而非As2S5.有大量研究报道了细菌对重金属的吸附作用[42, 43], 如尹雪斐等[44]的研究表明1株蜡状芽孢杆菌(Bacillus cereus sp.), 其对Cd的吸附原因主要是通过胞外沉淀作用, Cd与—COOH和—OH等官能团发生络合作用. Du等[45]报道表明细菌表面的—COOH等活性官能团参与Cd的固定, 以上报道与本研究的结论类似.本研究的XPS结果显示该菌除了发挥硫酸盐还原功能, 其表面丰富的官能团也发挥了生物吸附作用.参与固定Cd和As后C=O/C—O—C和C—(O, N)这两类官能团所归属的峰面积分别增加了3.22%和3.74%, 表明金属离子与微生物的羧基、羟基和酰胺基的氢原子之间的离子交换主要参与生物吸附.
土壤重金属污染, 尤其是Cd和As复合污染, 不仅危及生物圈中的动植物生长, 更涉及人类的食品安全问题.由于镉、砷存在形式和化学性质等不同, 能够对其同时钝化有一定难度.目前微生物修复手段, 一方面是利用微生物的胞外聚合物以及官能团对金属离子进行吸附固定, 另一方面在于某些菌株功能性的发挥.在本研究中发现的肠杆菌既能发挥其硫酸盐还原功能又能利用表面官能团同时钝化Cd和As, 此外, 由于该菌筛选自南方弱酸性农田土壤并曾长期暴露于自然环境中, 所以能够适应普遍的农田环境, 具有良好的应用前景.综上, 本研究为微生物应用修复重金属及类金属污染土壤提供思路和理论依据.
4 结论(1) 从镉、砷复合污染的农田土壤中分离得到1株耐镉、砷菌株M5, 经鉴定该菌种为肠杆菌属(Enterobacter sp.), 该菌株具有硫酸盐还原功能, 对镉、砷有高耐受性.
(2) 在模拟体系中, 菌株M5对镉去除率最大达94.13%, 对砷去除率最大为27.26%.SEM-EDS及XRD的结果证实了Cd和As被固定成CdS和As2S3, XPS结果表明该菌表面的羧基、羟基和酰胺基主要参与生物吸附.
| [1] |
徐长春, 郑戈, 林友华. "十三五"国家重点研发计划农田镉砷污染防治领域资助情况概述[J]. 农业环境科学学报, 2018, 37(7): 1321-1325. Xu C C, Zheng G, Lin Y H. Brief introduction to research projects on prevention and control of cadmium and arsenic pollution in croplands supported by National Key R & D Program of China in 13th Five-Year Period[J]. Journal of Agro-Environment Science, 2018, 37(7): 1321-1325. |
| [2] | Zhao F J, Ma Y B, Zhu Y G, et al. Soil contamination in China: Current status and mitigation strategies[J]. Environmental Science & Technology, 2015, 49(2): 750-759. |
| [3] |
李园星露, 叶长城, 刘玉玲, 等. 硅肥耦合水分管理对复合污染稻田土壤As-Cd生物有效性及稻米累积阻控[J]. 环境科学, 2018, 39(2): 944-952. Li Y X L, Ye C C, Liu Y L, et al. Bioavailability of silicon fertilizer coupled water management on soil bioavailability and cumulative control of rice in compound contaminated paddy soils[J]. Environmental Science, 2018, 39(2): 944-952. DOI:10.13227/j.hjkx.201707011 |
| [4] | Gu J F, Zhou H, Yang W T, et al. Effects of an additive (hydroxyapatite-biochar-zeolite) on the chemical speciation of Cd and As in paddy soils and their accumulation and translocation in rice plants[J]. Environmental Science and Pollution Research, 2018, 25(9): 8608-8619. DOI:10.1007/s11356-017-0921-2 |
| [5] | Mu T T, Wu T Z, Zhou T, et al. Geographical variation in arsenic, cadmium, and lead of soils and rice in the major rice producing regions of China[J]. Science of the Total Environment, 2019, 677: 373-381. DOI:10.1016/j.scitotenv.2019.04.337 |
| [6] |
周俊驰, 铁柏清, 刘孝利, 等. 湖南矿区县域耕地重金属污染空间特征及潜在风险评价[J]. 湖南农业科学, 2017(4): 75-79, 82. Zhou J C, Tie B Q, Liu X L, et al. Spatial characteristics and potential risk assessment of heavy metal pollution in farmland in Hunan Mining Area[J]. Hunan Agricultural Sciences, 2017(4): 75-79, 82. |
| [7] | Mohan D, Singh K P. Single-and multi-component adsorption of cadmium and zinc using activated carbon derived from bagasse-an agricultural waste[J]. Water Research, 2002, 36(9): 2304-2318. DOI:10.1016/S0043-1354(01)00447-X |
| [8] |
王英杰, 邹佳玲, 杨文弢, 等. 组配改良剂对稻田系统Pb、Cd和As生物有效性的协同调控[J]. 环境科学, 2016, 37(10): 4004-4010. Wang Y J, Zou J L, Yang W T, et al. Synergetic control of bioavailability of Pb, Cd and As in the rice paddy system by combined amendments[J]. Environmental Science, 2016, 37(10): 4004-4010. |
| [9] |
朱维, 周航, 吴玉俊, 等. 组配改良剂对稻田土壤中镉铅形态及糙米中镉铅累积的影响[J]. 环境科学学报, 2015, 35(11): 3688-3694. Zhu W, Zhou H, Wu Y J, et al. Effects of combined amendment on forms of cadmium/lead in paddy soil and accumulation of cadmium/lead in brown rice[J]. Acta Scientiae Circumstantiae, 2015, 35(11): 3688-3694. |
| [10] | Luo L, Ma C Y, Ma Y B, et al. New insights into the sorption mechanism of cadmium on red mud[J]. Environmental Pollution, 2011, 159(5): 1108-1113. DOI:10.1016/j.envpol.2011.02.019 |
| [11] | Tseng C H. Cardiovascular disease in arsenic-exposed subjects living in the arseniasis-hyperendemic areas in Taiwan[J]. Atherosclerosis, 2008, 199(1): 12-18. DOI:10.1016/j.atherosclerosis.2008.02.013 |
| [12] |
连斌, 吴骥子, 赵科理, 等. 铁锰氧化物-微生物负载生物质炭材料对镉和砷的吸附机制[J]. 环境科学, 2022, 43(3): 1584-1595. Lian B, Wu J Z, Zhao K L, et al. Novel insight into the adsorption mechanism of Fe-Mn oxide-microbe combined biochar for Cd(Ⅱ) and As(Ⅲ)[J]. Environmental Science, 2022, 43(3): 1584-1595. |
| [13] |
丁萍, 贺玉龙, 何欢, 等. 复合改良剂FZB对砷镉污染土壤的修复效果[J]. 环境科学, 2021, 42(2): 917-924. Ding P, He Y L, He H, et al. Remediation effect of compound modifier FZB on arsenic and cadmium contaminated soil[J]. Environmental Science, 2021, 42(2): 917-924. |
| [14] | Wang X Q, Yu H Y, Li F B, et al. Enhanced immobilization of arsenic and cadmium in a paddy soil by combined applications of woody peat and Fe(NO3)3: possible mechanisms and environmental implications[J]. Science of the Total Environment, 2019, 649: 535-543. DOI:10.1016/j.scitotenv.2018.08.387 |
| [15] |
黄德乾, 汪鹏, 王玉军, 等. 污染土壤上水稻生长及对Pb、Cd和As的吸收[J]. 土壤, 2008, 40(4): 626-629. Huang D Q, Wang P, Wang Y J, et al. Growth and uptake of Pb, Cd and As of rice in polluted soils[J]. Soils, 2008, 40(4): 626-629. DOI:10.3321/j.issn:0253-9829.2008.04.023 |
| [16] | Wu J Z, Huang D, Liu X M, et al. Remediation of As(Ⅲ) and Cd(Ⅱ) co-contamination and its mechanism in aqueous systems by a novel calcium-based magnetic biochar[J]. Journal of Hazardous Materials, 2018, 348: 10-19. DOI:10.1016/j.jhazmat.2018.01.011 |
| [17] |
牛之欣, 孙丽娜, 孙铁珩. 重金属污染土壤的植物-微生物联合修复研究进展[J]. 生态学杂志, 2009, 28(11): 2366-2373. Niu Z X, Sun L N, Sun T H. Plant-microoaganism combined remediation of heavy metals-contaminated soils[J]. Chinese Journal of Ecology, 2009, 28(11): 2366-2373. |
| [18] | Kuffner M, Puschenreiter M, Wieshammer G, et al. Rhizosphere bacteria affect growth and metal uptake of heavy metal accumulating willows[J]. Plant and Soil, 2008, 304(1): 35-44. |
| [19] | Li L J, Wang S T, Li X Z, et al. Effects of Pseudomonas chenduensis and biochar on cadmium availability and microbial community in the paddy soil[J]. Science of the Total Environment, 2018, 640-641: 1034-1043. DOI:10.1016/j.scitotenv.2018.05.287 |
| [20] | Saeed Z, Naveed M, Imran M, et al. Combined use of Enterobacter sp. MN17 and zeolite reverts the adverse effects of cadmium on growth, physiology and antioxidant activity of Brassica napus[J]. PLoS One, 2019, 14(3). DOI:10.1371/journal.pone.0213016 |
| [21] | Naveed M, Mustafa A, Azhar S Q T A, et al. Burkholderia phytofirmans PsJN and tree twigs derived biochar together retrieved Pb-induced growth, physiological and biochemical disturbances by minimizing its uptake and translocation in mung bean (Vigna radiata L.)[J]. Journal of Environmental Management, 2020, 257. DOI:10.1016/j.jenvman.2019.109974 |
| [22] |
张雅荟, 刘月敏, 王常荣, 等. 伯克氏菌Y4对水稻幼苗镉损伤和镉吸收的影响[J]. 环境科学, 2022, 43(4): 2142-2150. Zhang Y H, Liu Y M, Wang C R, et al. Effects of Burkholderia sp. Y4 on cadmium damage and uptake in rice seedlings[J]. Environmental Science, 2022, 43(4): 2142-2150. |
| [23] | Chiboub M, Saadani O, Fatnassi I C, et al. Characterization of efficient plant-growth-promoting bacteria isolated from Sulla coronaria resistant to cadmium and to other heavy metals[J]. Comptes Rendus Biologies, 2016, 339(9-10): 391-398. |
| [24] | Prapagdee B, Wankumpha J. Phytoremediation of cadmium-polluted soil by Chlorophytum laxum combined with chitosan-immobilized cadmium-resistant bacteria[J]. Environmental Science and Pollution Research, 2017, 24(23): 19249-19258. |
| [25] | Xu C R, He S B, Liu Y M, et al. Bioadsorption and biostabilization of cadmium by Enterobacter cloacae TU[J]. Chemosphere, 2017, 173: 622-629. |
| [26] | Peng W H, Li X M, Liu T, et al. Biostabilization of cadmium contaminated sediments using indigenous sulfate reducing bacteria: efficiency and process[J]. Chemosphere, 2018, 201: 697-707. |
| [27] | Zhao Q, Li X M, Xiao S T, et al. Integrated remediation of sulfate reducing bacteria and nano zero valent iron on cadmium contaminated sediments[J]. Journal of Hazardous Materials, 2021, 406. DOI:10.1016/j.jhazmat.2020.124680 |
| [28] | Shi W J, Song W J, Zheng J L, et al. Factors and pathways regulating the release and transformation of arsenic mediated by reduction processes of dissimilated iron and sulfate[J]. Science of the Total Environment, 2021, 768. DOI:10.1016/j.scitotenv.2020.144697 |
| [29] | Xia X, Li J H, Liao S J, et al. Draft genomic sequence of a chromate-and sulfate-reducing Alishewanella strain with the ability to bioremediate Cr and Cd contamination[J]. Standards in Genomic Sciences, 2016, 11(1): 48. |
| [30] | Alam R, McPhedran K N. Applications of biological sulfate reduction for remediation of arsenic-A review[J]. Chemosphere, 2019, 222: 932-944. |
| [31] | Rodriguez-Freire L, Sierra-Alvarez R, Root R, et al. Biomineralization of arsenate to arsenic sulfides is greatly enhanced at mildly acidic conditions[J]. Water Research, 2014, 66: 242-253. |
| [32] | Kocar B D, Borch T, Fendorf S. Arsenic repartitioning during biogenic sulfidization and transformation of ferrihydrite[J]. Geochimica et Cosmochimica Acta, 2010, 74(3): 980-994. |
| [33] | Rodriguez-Freire L, Moore S E, Sierra-Alvarez R, et al. Arsenic remediation by formation of arsenic sulfide minerals in a continuous anaerobic bioreactor[J]. Biotechnology and Bioengineering, 2016, 113(3): 522-530. |
| [34] | Le Pape P, Battaglia-Brunet F, Parmentier M, et al. Complete removal of arsenic and zinc from a heavily contaminated acid mine drainage via an indigenous SRB consortium[J]. Journal of Hazardous Materials, 2017, 321: 764-772. |
| [35] | Postgate J R. The Sulphate-reducing bacteria[M]. (2nd ed). Cambridge: Cambridge University Press, 1984. |
| [36] |
万海清, 苏仕军, 葛长海, 等. 一种分离培养硫酸盐还原菌的改进方法[J]. 应用与环境生物学报, 2003, 9(5): 561-562. Wan H Q, Su S J, Ge C H, et al. A improved method for islation and cultivation of sulfate-reducing bacteria[J]. Chinese Journal of Applied and Environmental Biology, 2003, 9(5): 561-562. |
| [37] | Tamura K, Stecher G, Peterson D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution, 2013, 30(12): 2725-2729. |
| [38] | Lei M, Tao J, Yang R J, et al. Binding of Sb (Ⅲ) by Sb-tolerant Bacillus cereus cell and cell-goethite composite: implications for Sb mobility and fate in soils and sediments[J]. Journal of Soils and Sediments, 2019, 19(6): 2850-2858. |
| [39] |
李杨, 周丽, 杜辉辉, 等. Cd(Ⅱ)与As(Ⅴ)在一株土壤细菌Delftia sp.上的共吸附研究[J]. 土壤, 2020, 52(5): 935-940. Li Y, Zhou L, Du H H, et al. Co-adsorption of Cd(Ⅱ) and As(Ⅴ) to soil bacterium Delftia sp.[J]. Soils, 2020, 52(5): 935-940. |
| [40] | Hoffmann H, Stindl S, Stumpf A, et al. Description of Enterobacter ludwigii sp. nov., a novel Enterobacter species of clinical relevance[J]. Systematic and Applied Microbiology, 2005, 28(3): 206-212. |
| [41] | Wang X, Hu K, Xu Q, et al. Immobilization of Cd using mixed Enterobacter and Comamonas bacterial reagents in pot experiments with Brassica rapa L.[J]. Environmental Science & Technology, 2020, 54(24): 15731-15741. |
| [42] |
刘玉玲, 朱虎成, 彭鸥, 等. 玉米秸秆生物炭固化细菌对镉砷吸附[J]. 环境科学, 2020, 41(9): 4322-4332. Liu Y L, Zhu H C, Peng O, et al. Adsorption of cadmium and arsenic by corn stalk biochar solidified microorganism[J]. Environmental Science, 2020, 41(9): 4322-4332. |
| [43] |
王晓宇, 韩辉. 重金属固定细菌调控土壤铁氧化物阻控小麦Cd吸收效应[J]. 环境科学, 2020, 41(11): 5160-5167. Wang X Y, Han H. Effects of heavy metal-immobilizing bacteria on reducing wheat uptake Cd by regulation soil iron oxides[J]. Environmental Science, 2020, 41(11): 5160-5167. |
| [44] |
尹雪斐, 杨蕊嘉, 刘玉玲, 等. Cd(Ⅱ)与As(Ⅴ)在土壤铁氧化物和细菌表面上的共吸附研究[J]. 生态环境学报, 2021, 30(3): 614-620. Yin X F, Yang R J, Liu Y L, et al. Co-adsorption of Cd(Ⅱ) and As(Ⅴ) on soil iron oxide and bacterial surface[J]. Ecology and Environmental Sciences, 2021, 30(3): 614-620. |
| [45] | Du H H, Chen W L, Cai P, et al. Cd(Ⅱ) Sorption on montmorillonite-humic acid-Bacteria composites[J]. Scientific Reports, 2016, 6: 2505-2511. |