 2023, Vol. 44
2023, Vol. 44



2. 青岛海洋科学与技术试点国家实验室海洋生态与环境科学功能实验室, 青岛 266237
2. Laboratory for Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266237, China
生物气溶胶是指附着有微生物或生物大分子等具有生命活性物质的气溶胶粒子[1, 2], 其组成复杂, 来源广泛, 可以在全球尺度上进行传输[3].细菌作为生物气溶胶的重要组成部分, 在PM2.5和PM10的生物组分中占比高达80%以上[4], 会导致各种病原体和过敏原的扩散[5, 6], 对人类健康产生重要影响.另外, 空气中细菌可能会对云化学产生重要影响[7], 直接或间接影响全球气候变化[8, 9].作为空气中最丰富和分布最广的微生物[10], 细菌对人类健康、区域气候[11]和生物地球化学循环[12]都具有重要的意义.
目前, 生物气溶胶中活细菌浓度测定多采用活菌培养计数法[13]、ATP生物发光法[14]、定量PCR法[13]和荧光显微镜法.其中, 荧光显微镜法操作简单, 结果可靠, 有效地克服了部分微生物无法检出的短板, 在饮用水[15]、淤泥[16]和大气中死/活细菌浓度[17, 18]测定上获得了广泛应用.大气中细菌的浓度普遍在104~108 cells·m-3[19], 活细菌浓度在102~104 cells·m-3[20, 21], 并且存在明显的季节差异, 比如美国北科罗拉多州高海拔地区空气中细菌最高浓度出现在春秋季[22]; 北京[23]、敦煌[24]和希腊塞萨洛尼基[25]地区夏季生物气溶胶中细菌浓度最高; 青岛地区空气中总细菌浓度发现在秋季浓度最高[26].同时, 不同特殊天气条件下(雾、霾和沙尘), 空气中细菌浓度分布特征存在一定差异.重污染天中总细菌浓度为(2.75±3.25)×105 cells·m-3, 要高于非污染天[20]; 雾天活细菌浓度为7.13×103 cells·m-3, 明显低于晴天的1.15×104 cells·m-3[21]; 沙尘天空气中细菌总浓度是非沙尘天气时的40倍[27], 活细菌占细菌总数的比例在16% ~40%之间, 比非沙尘天细菌存活能力小得多[28].可见, 空气中细菌浓度受到气象、源等许多生物和非生物因素的综合影响, 因此, 研究雾和沙尘等典型污染过程下大气气溶胶中细菌浓度特征, 可为进一步评价大气生物气溶胶的环境和健康风险奠定科学基础.
生物气溶胶粒径的大小会影响微生物在人体呼吸道的沉积及引发的呼吸道疾病.大部分研究表明, 空气中细菌主要分布在粗粒径(>2.1 μm)上[18, 28], 如青岛沿海[26]和美国华盛顿[29]地区大气中细菌浓度峰值分布在>7 μm粒径上, 北京室外空气中细菌呈偏态分布, 主要分布在>2.1 μm的粒径范围内[30].在不同特殊天气条件下, 空气中细菌粒径分布特征同样存在一定差异.青岛地区雾、霾和沙尘天空气中活细菌均呈现出双峰分布, 峰值分布在1.1~2.1 μm和4.7~7.0 μm[21]; 北京市霾天大气中粒径<2.1 μm的细菌占比随着霾强度的增加而逐渐降低, 粒径分布向粗模态迁移[31], 而西安市霾天气下空气细菌粒径的偏态分布规律和晴天相似, 没有太大变化[32].显然, 目前研究对大气中细菌的分布规律仍不明确, 对雾天、沙尘天下存活微生物粒径分布的研究有助于探索生物气溶胶的健康效应.
目前对于空气中细菌的研究已经取得一定的进展, 但是大部分研究集中于空气中总细菌及可培养菌浓度, 而对于空气中存活细菌的系统研究有限, 不同污染天气对细菌存活率的影响研究更为匮乏.因此, 本文于2020年9月至2021年8月在青岛近海地区连续采集生物气溶胶样品, 分析了样品中死活细菌月季分布特征, 重点讨论了雾天和沙尘天气对气溶胶中细菌存活率的影响.
1 材料与方法 1.1 采样地点及方法如图 1所示, 样品采集地点位于中国海洋大学崂山校区监测点(36°10′N, 120°30′E), 距海边约7.0 km, 周围的植被覆盖面积约占50%.样品利用FA-1撞击式生物采样器(辽阳康洁仪器研究所)28.3 L·min-1流量下采集30 min, 样品收集于直径80 mm、孔径0.2 μm灭菌的聚碳酸酯滤膜上.常规样品分别于每月的5、10、15、20和25日08:00采集生物气溶胶样品, 污染天气增加样品采集频率.FA-1采样器分六级, 切割粒径范围分别为:>7.0、4.7~7.0、3.3~4.7、2.1~3.3、1.1~2.1和0.65~1.1 μm.

|
图 1 生物气溶胶采样点示意 Fig. 1 Location of the sampling site for bioaerosols |
将样品膜放入20 mL生理盐水中, 以150 r·min-1振荡培养30 min, 制成菌悬液.将10 mL悬浮液用BacLightTM细菌活力试剂盒(L-13152, Thermo Fisher Scientific, USA)在黑暗条件下染色15 min[20].试剂盒主要包括36 nmol·L-1 SYTO-9和180 nmol·L-1碘化丙啶(PI).染色后, 利用0.2 μm黑色聚碳酸酯膜过滤器(Whatman Inc., USA)过滤.使用放大400倍的荧光显微镜(Olympus BX51, 日本)测定染色样品.SYTO-9染色的活细菌在450~480 nm的激发波长下测定, 呈绿色荧光, 而PI染色的死细菌在510~550 nm的激发波长下测定, 呈红色荧光[33].随机选取20个视野, 记录视野中具有细菌形态、直径小于1 μm的菌个数并根据下列公式进行计算[26].
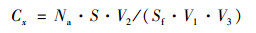
|
(1) |
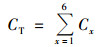
|
(2) |
式中, CT表示6级粒径中死/活细菌浓度之和(cells·m-3); Cx表示x粒径死/活细菌浓度(cells·m-3); Na表示各视野平均死/活细菌粒子数(cells); S表示核孔滤膜实际过滤面积(mm2); Sf表示显微镜视野面积(mm2); V1表示过滤样品量(L); V2表示氯化钠缓冲液体积(L); V3表示采样器空气流量(m3).
1.3 气象参数来源以及数据分析气象参数[温度(T)、相对湿度(RH)、风向(WD)、风速(WS)、能见度]和环境指标(PM2.5、PM10、SO2、NO2、CO、O3)分别来自青岛市气象局(http://qdqx.qingdao.gov.cn/)和青岛市环境保护局(http://www.qepb.gov.cn/m2/).青岛受海洋气候影响, 根据秋季(9~11月)、冬季(12月至次年2月)、春季(3~5月)以及夏季(6~8月)进行数据分类分析细菌浓度的季节变化.另外, 根据MICAPS资料天气类型, 将样品分为晴天、霾天、雾天和沙尘天样品, 其中霾天、雾天和沙尘天具体分类标准如表 1所示.Spearman相关性用于研究微生物与气象因素的相关性, 其中P>0.05表示无显著相关性, P<0.05则具有统计学意义.利用后向轨迹模型(HYSPLIT-4)[34]和全球数据同化系统(GDAS)气象数据(http://www.arl.noaa.gov/ready/hysplit4.html), 分析采样期间气团来源, 每条轨迹模拟后推72 h, 高度为1 000 m.
|
|
表 1 不同天气类型分类标准1) Table 1 Classification criteria for different weather types |
2 结果与讨论 2.1 青岛近海生物气溶胶中细菌浓度月季分布特征
2020年9月至2021年8月期间, 共采集生物气溶胶样品77套(图 2), 细菌浓度范围在3.19×104 cells·m-3~3.34×105 cells·m-3, 平均值为(1.06±0.68)×105 cells·m-3.其中, 活细菌浓度范围在3.16×103 cells·m-3~2.53×104 cells·m-3, 平均值为(8.20±4.88)×103 cells·m-3; 死细菌浓度在2.97×104 cells·m-3~3.19×105 cells·m-3, 平均值为(9.74±6.72)×104 cells·m-3. 2017年8月至2018年2月青岛地区采用相同方法测得生物气溶胶中细菌平均值为(9.21±9.07)×104 cells·m-3, 其中活细菌平均值为(1.89±1.83)×104 cells·m-3[20]; 2018年3月至2019年2月青岛地区生物气溶胶中细菌浓度为(8.45±6.37)×104 cells·m-3, 其中活细菌浓度为(1.09±0.71)×104 cells·m-3[21].以上结果表明, 由于气象条件的变化, 同一地区大气中细菌会随着年度变化而有所差异.

|
图 2 2020年9月至2021年8月青岛大气生物气溶胶中细菌浓度分布 Fig. 2 Concentration variation in bacteria in bioaerosols samples in Qingdao from Sep. 2020 to Aug. 2021 |
青岛地区大气中活细菌浓度在春季和夏季较高(表 2), 平均值分别为(1.08±0.64)×104 cells·m-3和(1.01±0.31)×104 cells·m-3, 秋季次之, 为(7.84±3.65)×103 cells·m-3, 冬季最低, 为(4.94±2.47)×103 cells·m-3, 其中冬季与春季和夏季存在显著性差异(P<0.05).死细菌浓度在春季最高, 浓度为(1.32±0.85)×105cells·m-3, 冬季和秋季分别为(1.12±0.61)×105 cells·m-3和(7.42±1.92)×104 cells·m-3, 夏季最低, 浓度为(4.56±3.20)×104 cells·m-3.秋季与春季以及夏季与冬季、春季均呈现显著性差异(P<0.05).总细菌浓度的季节分布与死细菌相似, 呈现出春季和冬季最高, 夏季最低的季节变化, 且春季与夏季、秋季以及冬季与夏季存在显著差异.这与当地细菌来源、气象条件和空气环境的季节性变化有关.
|
|
表 2 2020年9月至2021年8月青岛大气生物气溶胶中细菌浓度以及相关参数季节变化 Table 2 Seasonal average of bacteria concentrations in atmospheric bioaerosols and related parameters in Qingdao from Sep. 2020 to Aug. 2021 |
春季细菌的高浓度首先源于微生物来源的增加, 土壤和植被等局地源对大气气溶胶中细菌浓度具有重要影响[35, 36], 春季植物生长、开花等生命活动会向空气中释放较多的微生物[37, 38], 春耕活动的增加, 也会造成大气中活/死细菌浓度均有增加.其次, 春季沙尘事件频繁发生, 在采样期间共出现3次, 沙尘期间风速最高达到14.42 m·s-1, 有助于空气中细菌的长距离传输, 极大地促进了大气中细菌浓度的增加[18].这3次沙尘事件细菌存活率较低, 在4.26% ~9.11%, 这可能是由于在长距离运输过程中, 细菌会受到紫外线辐射以及干燥环境影响[39], 从而大量死亡, 因此沙尘天造成死细菌浓度增加比例更高.再次, 相关性分析发现活细菌浓度与SO2和NO2呈负相关(P<0.05, 图 3), NO2在空气中会形成硝酸和亚硝酸, 可使蛋白质脱氨基[40], SO2也被认为是对微生物有毒的污染物[41], 春季SO2和NO2浓度低, 利于细菌存活.此外, 微生物失活率与紫外线辐射强度成正比[42], 春季太阳辐射强度较低, 也是造成活细菌浓度较高的原因.

|
椭圆方向表示正负相关, 其中向右表示正相关, 向左表示负相关, 椭圆宽度越窄表示相关性系数越接近±1, *表示在0.05水平上显著相关(双尾检验) 图 3 青岛生物气溶胶中细菌浓度与环境因素之间的Spearman's相关系数 Fig. 3 Spearman's correlation coefficient between the concentrations of bacteria in bioaerosol samples and environmental factors |
夏季活细菌的高浓度主要是因为大气环境相对适宜细菌存活.相关性分析发现气温与活细菌的浓度呈显著正相关(P<0.05, 图 3), 适宜的温度更易促进微生物的释放和生长[43], 大气中95%的细菌是嗜温细菌[44], 最佳生存温度在20~40℃之间[45], 而夏季平均温度为24.3℃, 适宜细菌生长繁殖.虽然夏季太阳辐射较强, 但是RH较高, 微生物表面的水层在一定程度上有助于抵抗太阳辐射造成的损害, 从而保持微生物的生存能力[46].因此细菌存活率在夏季最高, 为(21.72±7.51)%.相关性分析发现, 死细菌浓度与PM呈正相关, 冬季ρ(PM2.5)和ρ(PM10)偏高, 分别达到79 μg·m-3和139 μg·m-3, 这有利于细菌聚集在颗粒上, 免受恶劣的大气条件(如紫外线照射)的影响[47], 但是冬季低温本身就不利于细菌存活[21], 同时取暖等人类活动使得霾事件频发, 大气中有毒有害物质不利于细菌存活[48], 造成大气中细菌的大量死亡, 因此冬季细菌存活率最低, 平均值为(4.24±4.23)%.
2.1.2 青岛近海生物气溶胶中死/活细菌浓度的月变化图 4为青岛近海大气生物气溶胶中细菌的月均浓度分布.青岛大气中细菌浓度随月份有所波动.大气中活细菌月均浓度最高值出现在2021年5月, 为(1.48±0.72)×104 cells·m-3, 而最低值在2020年12月, 为(3.28±2.23)×103 cells·m-3.大气气溶胶中死细菌浓度的月变化与活细菌不同, 最高值出现在2021年3月, 为(1.53±0.91)×105 cells·m-3, 而最低值(3.54±1.53)×104 cells·m-3出现在2021年6月.总细菌的月变化和死细菌相似, 这是因为死细菌在细菌中占有很大比例, 平均值为88.34%.

|
图 4 2020年9月至2021年8月青岛生物气溶胶中细菌浓度及相关参数的月变化 Fig. 4 Monthly average concentrations of bacteria in bioaerosol samples and related parameters in Qingdao from Sep. 2020 to Aug. 2021 |
从9~11月, 气温逐渐下降, 从22℃下降到8℃, 不利于大气中细菌的生长[49], 细菌浓度逐渐降低.12月ρ(PM2.5)到了最高值102 μg·m-3, 有利于细菌在空气中附着从而浓度增加[50], 但此时青岛地区气温较低, 平均值为3.08℃, 同时燃煤、生物质燃烧以及车辆排放等人为活动会造成空气中有毒有害化学物质的增加[51], 如高浓度的K+和Na+可能会破坏细菌的结构和功能, 导致细菌的死亡[52], 相应活细菌浓度降低, 死细菌浓度升高.12月大气中细菌存活率达到了最低值, 为2.35%, 较高的SO2和NO2可能也是造成活细菌浓度偏低的一个重要原因.2021年1~2月, 大气中死/活细菌浓度在逐月下降, 在3月份均有所增加, 3月的死/活细菌浓度分别是2月的2.59倍和2.19倍, 此时PM浓度较高, 沙尘天气频发, PM10达到最高值为320 μg·m-3, 大气中细菌外来源的增加造成青岛地区细菌浓度的增加.5月份细菌浓度维持在较高水平.进入夏季后, 6~8月, 活细菌浓度则较高, 细菌存活率在6月和8月分别达到了(22.49±4.37)%和(23.73±3.86)%, 这与气象因素逐渐适宜细菌的生长和繁殖有关.
2.2 青岛近海生物气溶胶中死/活细菌粒径月季分布特征 2.2.1 死/活细菌粒径分布的季节变化图 5为2020年9月至2021年8月青岛生物气溶胶中细菌粒径分布的季节变化.如图 5(a)所示, 青岛沿海地区大气中活细菌粒径分布的季节差异较大, 不同季节各粒径上的浓度占比也存在较大差异.秋季近似单峰分布, 峰值在4.7~7.0 μm, 占比为19.51%; 而冬季和夏季则呈现双峰分布, 冬季峰值位于1.1~2.1 μm和4.7~7.0 μm, 比例分别为21.32%和18.60%, 而夏季峰值为1.1~2.1 μm和>7.0 μm, 占比分别为17.62%和19.53%; 春季活细菌浓度在0.65~1.1 μm粒径上占比最低, 为12.73%, 在其他粒径上均匀分布, 占比在16.64% ~18.11%.死细菌的粒径分布主要分为近似偏态分布和双峰分布[图 5(b)], 其中秋季样品粒径近似偏态分布, 3.3~4.7 μm粒径占比较高, 为20.41%; 而冬季、春季和夏季均呈现出双峰分布, 冬季峰值位于1.1~2.1 μm和>7.0 μm, 占比分别为19.89%和18.65%, 春季峰值则位于2.1~3.3 μm和4.7~7.0 μm, 占比分别为20.56%和17.65%, 而夏季死细菌浓度峰值则位于2.1~3.3 μm和>7.0 μm, 占比分别为18.60%和20.31%.总细菌粒径分布与死细菌相似.总体而言, 大气中死/活细菌仍主要附着在粗粒径颗粒物上, 这与之前的研究保持一致[20, 53, 54].夏季活细菌在粗粒径上占比较低, 为67.62%, 这可能是由于青岛地区夏季气团主要来自海上[55], 海洋飞沫是生物气溶胶重要的来源[45], 有研究表明海面液滴的平均直径小于城市雾滴的平均直径[56].冬季活/死细菌在粗粒子中占比最低, 分别为62.3%和69.79%, 这与Yin等[21]的研究结果相似.冬季ρ(PM2.5)在4个季节中较高, 为(78.54±44.39) μg·m-3, 虽然PM10浓度也不低, 但是有研究表明PM2.5携带的微生物丰度要高于PM10[57], 这可能是引起冬季死/活细菌粗粒径占比最低的原因.

|
图 5 2020年9月至2021年8月青岛生物气溶胶中细菌的粒径分布季节变化 Fig. 5 Seasonal average of bacteria size distributions in bioaerosols samples in Qingdao from Sep. 2020 to Aug. 2021 |
本研究发现, 总细菌和死细菌粒径分布相似, 活细菌粒径分布略有不同.粒径分布总体趋势可分为两类, 即单峰分布和双峰分布, 不同月份的峰值情况也存在着差异性.如图 6(a)所示, 2020年9月和12月以及2021年的1、3和5月活细菌粒径分布呈现双峰分布, 峰值为1.1~2.1 μm和4.7~7.0 μm, 范围在18.26% ~23.59%; 而2021年的4、6和8月双峰则位于0.65~1.1 μm和4.7~7.0 μm.其余月份则呈现为单峰分布, 2020年10月峰值位于1.1~2.1 μm, 而2020年11月则位于3.3~4.7 μm, 2021年的2月和7月峰值均位于4.7~7.0 μm.结果显示, 不同月份大气中活细菌粒径分布差异较大.如图 6(b)和6(c)所示, 2020年的9、10、11月以及2021年8月青岛地区总细菌和死细菌粒径呈单峰分布, 高值出现在1.1~2.1、0.65~1.1、2.1~3.3和1.1~2.1 μm.而其余月份总细菌和死细菌粒径均呈现双峰分布, 2020年12月和2021年1月峰值为0.65~1.1 μm和4.7~7.0 μm, 且峰值比例在18.97% ~24.31%; 2021年2、3、5月及7月峰值为1.1~2.1 μm和4.7~7.0 μm, 峰值比例在18.12% ~25.02%; 2021年4月峰值位于1.1~2.1 μm和3.3~4.7 μm, 而2021年6月峰值则位于0.65~1.1 μm和3.3~4.7 μm.虽然在不同月份死活细菌的粒径分布存在一定的差异性, 但整体来说, 细菌主要附着在粗粒径上, 这是因为粗颗粒物中污染物富集低于细粒子, 能较好地为微生物提供庇护[47].其中12月活细菌在粗粒径上占比最低, 为60.81%, 该月是青岛地区霾天多发期, 有研究表明, 在霾天活细菌在粗粒径中占比最低[21].

|
图 6 2020年9月至2021年8月青岛生物气溶胶中细菌月均粒径分布 Fig. 6 Monthly averaged size distributions of bacteria in bioaerosol samples in Qingdao from Sep. 2020 to Aug. 2021 |

|
*表示两者浓度之间存在显著性差异(P<0.05) 图 7 2020年9月至2021年8月青岛不同天气下生物气溶胶中细菌浓度箱式图 Fig. 7 Concentrations of bacteria in bioaerosol samples under various weather conditions in Qingdao from Sep. 2020 to Aug. 2021 |
为了对比分析大气污染对细菌浓度的影响, 本研究根据采样期间的天气类型[20, 58], 将77份生物气溶胶样品分为晴天、雾天、霾天和沙尘天这4类, 样品数量分别为2、5、14和7个.图 7比较了不同天气类别下的细菌浓度, 结果显示青岛地区大气中细菌的平均浓度受到天气类型的影响.霾天活细菌浓度平均值是最低的, 为(4.34±2.55)×103 cells·m-3, 雾天活细菌浓度平均值为(7.15±2.61)×103 cells·m-3, 均低于晴天浓度[(7.93±5.05)×103 cells·m-3], 分别降低了43.15%和9.84%.沙尘天活细菌浓度为(16.19±5.49)×103 cells·m-3, 是晴天的2.04倍.死细菌浓度在雾天最低, 平均值为(4.39±2.08)×104 cells·m-3, 霾天和沙尘天死细菌浓度则是晴天的1.50倍和2.93倍, 平均值分别为(12.12±5.99)×104 cells·m-3和(23.74±6.41)×104 cells·m-3.其中, 沙尘天活/死细菌浓度与晴天、雾天和霾天均存在显著性差异(P<0.05), 霾天死细菌浓度与雾天也存在显著性差异(P<0.05).总细菌不同天气下浓度分布与死细菌相似.可见, 特殊天气对细菌的影响依赖于细菌类别和污染天气类型, 沙尘天气会使死活细菌浓度均升高, 雾天会使死活细菌浓度均有所降低, 霾天会使活细菌浓度降低而死细菌浓度增加.此外, 本研究发现雾天细菌存活率最高为16.26%, 晴天、霾天以及沙尘天细菌存活率分别为11.12%、4.10%和6.85%.
霾天低风速使得气溶胶的垂直和水平扩散均弱于其他天气, 易导致空气污染物的积累, 作为微生物载体的PM浓度较高[59].霾天下PM2.5和PM10的浓度分别达到了115 μg·m-3和172 μg·m-3, 是晴天的3.8倍和2.15倍.本研究进一步计算了单位质量PM10颗粒上的细菌负荷发现, 霾天下死/活细菌负荷均值分别为702 cells·μg-1和32 cells·μg-1, 均低于晴天998 cells·μg-1和98 cells·μg-1, 同时, 死细菌浓度与PM呈正相关关系(图 3), 这些结果均表明大气中死细菌浓度的增加是由于污染物在不利扩散条件下积累造成的[21, 60].青岛地区冬季供暖煤炭燃烧、车辆尾气排放等造成SO2和NO2的浓度高于其他天气[61], 同时, 霾天温度较低, 在-1~9℃, 平均值为3.9℃, 远低于细菌适宜生存温度[45], 因此霾天活细菌浓度较低, 造成大气中细菌存活率的降低.相较于晴天, 雾天PM更低, ρ(PM2.5)和ρ(PM10)仅为23 μg·m-3和36 μg·m-3(表 3), 雾天死活细菌负荷分别为1 213 cells·μg-1和197 cells·μg-1, 均高于晴天, 表明雾天大气中的液滴确实为大气中细菌的生长和繁殖提供了适宜的场所.微生物载体的减少可能是造成大气中细菌浓度降低的一个原因[21].雾天湿度达到了94%, 高湿度会促进空气中微生物的湿沉降[62], 影响空气中微生物的浓度和群落结构, 部分革兰氏阴性细菌不容易在雾天下存活[63], 造成总细菌浓度低于晴天.相较于霾天, 雾天对微生物有毒有害的化学物质浓度较低[64], SO2和NO2浓度最低, 使得大气中死细菌浓度较低, 雾天平均温度最接近细菌适宜生存的温度, 细菌存活率最高, 达到16.26%. 2020年9月和2021年8月期间青岛市发生三起沙尘事件, 均出现在春季.沙尘期间ρ(PM10)高达541.43 μg·m-3, ρ(PM2.5)为114.15 μg·m-3, 沙尘事件期间的WS(8.71 m·s-1)明显高于其他天气, 可扬起地面尘土, 引起生物气溶胶的远距离传输[65], 运输一些来自遥远地区的生物气溶胶, 从而改变大气中的细菌种类[66], 造成细菌浓度增加.沙尘期间(图 8), 气团途经蒙古戈壁沙漠以及内蒙古浑善达克沙地, 经过京津冀工业区达到青岛, 沙尘气团中携带的高浓度粗颗粒物在传输和扩散过程中易携带微生物[67], 造成死/活细菌浓度的增加.计算得出死细菌在PM10上的负荷较沙尘前明显升高, 进一步证明沙尘天气大气细菌浓度的增加是由于外源微生物的输入.相较于晴天, 沙尘天死细菌浓度增加量远超于活细菌浓度的增加量, 这是由于沙尘运输的细菌细胞会暴露在紫外线辐射、低温和干燥等大气不利环境因素中, 并受到超出其自身压力保护能力的损害[28].
|
|
表 3 2020年9月至2021年8月不同天气下青岛地区环境因素 Table 3 Environmental factors in Qingdao from Sep. 2020 to Aug. 2021 under different weathers |

|
图 8 青岛2021年4月16日沙尘期气团的72 h后向轨迹 Fig. 8 The 72 h air mass back trajectories in Qingdao during the dust periods in 16 Apr. 2021 |
本研究发现, 不同天气条件下青岛大气活细菌粒径分布均呈现出双峰分布(图 9).晴天和霾天峰值均位于1.1~2.1 μm和4.7~7.0 μm, 活细菌主要附着在粗颗粒上, 但是各粒径占比不同, 霾天活细菌向细粒径迁移, 活菌在粗粒径中的比例为63.48%, 要低于晴天的68.29%, 这与霾天PM2.5浓度的增加有关[33].相比之下, 雾天活细菌峰值分别位于1.1~2.1 μm和>7.0 μm, 占比分别为20.39%和21.60%, 高RH促进了颗粒沉积, 相较于晴天, 雾天活细菌粗粒径上占比下降了2.67%.沙尘天活细菌峰值则位于2.1~3.3 μm和4.7~7.0 μm, 占比分别为20.90%和19.24%, 活细菌粒径明显向粗粒径迁移, 占比提高到74.25%, 同样的结果出现在Yin等[21]研究中, 这与沙尘天粗颗粒的增加密切相关.

|
图 9 2020年9月至2021年8月青岛不同天气下生物气溶胶中细菌粒径分布 Fig. 9 Size distributions of bacteria in bioaerosol samples under various weather conditions from Sep. 2020 to Aug. 2021 in the Qingdao coastal region |
晴天死细菌在0.65~1.1 μm粒径上占比最低, 为11.61%, 其他粒径段死细菌占比处于18.82% ~19.59%, 而雾天、霾天以及沙尘天下死细菌粒径分布均呈现出双峰分布, 其中雾天和霾天峰值均位于1.1~2.1 μm和>7.0 μm, 沙尘天则出现在2.1~3.3 μm和4.7~7.0 μm.总体来说, 死细菌仍主要附着在粗粒径上, 晴天平均占比为75.95%, 雾天、霾天和沙尘天粗模态下的死活菌比例均有所下降, 分别为71.21%、69.83和73.62%. Gong等[20]同样发现霾天下死细菌在粗粒径上的占比低于晴天, 细模态下的微生物最有可能随着细颗粒浓度的增加而增加.Yin等[21]发现雾天下死细菌在粗粒径上的占比同样低于晴天, 这与高湿度造成的粒子沉降有关.总细菌粒径分布则与死细菌相似.
3 结论(1) 青岛地区大气中细菌浓度由于受微生物来源、气象条件及颗粒物浓度等环境因素的影响具有明显的季节差异, 死细菌浓度为:春季>冬季>秋季>夏季的趋势, 活细菌为:春季>夏季>秋季>冬季.细菌存活率则表现为夏季最高, 春季和秋季次之, 冬季最低. 相关性分析发现, 活细菌浓度与温度呈显著正相关, 与NO2、SO2和CO呈显著负相关; 死细菌与PM显著正相关.
(2) 大气中细菌浓度随着月份波动, 其中死细菌月均浓度最高值出现在2021年3月, 最低值出现在2021年6月; 活细菌月均浓度最高值出现在2021年5月, 而最低值在2020年12月.
(3) 青岛地区细菌粒径分布主要呈现双峰分布和单峰分布两类, 存在一定的月季差异, 但细菌主要分布在粗粒径上.
(4) 不同污染天气下生物气溶胶中细菌存在明显差异性, 沙尘天细菌浓度显著高于霾天、雾天以及晴天, 但细菌存活率较低.霾天细菌存活率最低, 为4.10%, 雾天细菌存活率最高, 为16.26%.不同污染天气死/活细菌粒径分布均为双峰分布, 但峰值随天气类型而变化.
| [1] | Després V R, Huffman J A, Burrows S M, et al. Primary biological aerosol particles in the atmosphere: a review[J]. Tellus B: Chemical and Physical Meteorology, 2012, 64(1). DOI:10.3402/tellusb.v64i0.15598 |
| [2] |
祁建华, 高会旺. 生物气溶胶研究进展: 环境与气候效应[J]. 生态环境, 2006, 15(4): 854-861. Qi J H, Gao H W. Environment and climate effect of bioaerosol: A review[J]. Ecology and Environment, 2006, 15(4): 854-861. DOI:10.3969/j.issn.1674-5906.2006.04.038 |
| [3] | Griffin D W. Atmospheric movement of microorganisms in clouds of desert dust and implications for human health[J]. Clinical Microbiology Reviews, 2007, 20(3): 459-477. DOI:10.1128/CMR.00039-06 |
| [4] | Cao C, Jiang W J, Wang B Y, et al. Inhalable microorganisms in Beijing's PM2.5 and PM10 pollutants during a severe smog event[J]. Environmental Science & Technology, 2014, 48(3): 1499-1507. |
| [5] | Yadav S, Gettu N, Swain B, et al. Bioaerosol impact on crop health over India due to emerging fungal diseases (EFDs): an important missing link[J]. Environmental Science and Pollution Research, 2020, 27(11): 12802-12829. DOI:10.1007/s11356-020-08059-x |
| [6] | Núñez A, de Paz G A, Rastrojo A, et al. Monitoring of airborne biological particles in outdoor atmosphere. Part 1: importance, variability and ratios[J]. International Microbiology, 2016, 19(1): 1-13. |
| [7] | Kourtev P S, Hill K A, Shepson P B, et al. Atmospheric cloud water contains a diverse bacterial community[J]. Atmospheric Environment, 2011, 45(30): 5399-5405. DOI:10.1016/j.atmosenv.2011.06.041 |
| [8] | Möhler O, DeMott P J, Vali G, et al. Microbiology and atmospheric processes: the role of biological particles in cloud physics[J]. Biogeosciences, 2007, 4(6): 1059-1071. DOI:10.5194/bg-4-1059-2007 |
| [9] | Sindoni O I, Saija R, Iatí M A, et al. Optical scattering by biological aerosols: experimental and computational results on spore simulants[J]. Optics Express, 2006, 14(15): 6942-6950. DOI:10.1364/OE.14.006942 |
| [10] | Burrows S M, Elbert W, Lawrence M G, et al. Bacteria in the global atmosphere-Part 1: review and synthesis of literature data for different ecosystems[J]. Atmospheric Chemistry and Physics, 2009, 9(23): 9263-9280. DOI:10.5194/acp-9-9263-2009 |
| [11] | Creamean J M, Suski K J, Rosenfeld D, et al. Dust and biological aerosols from the Sahara and Asia influence precipitation in the western U.S.[J]. Science, 2013, 339(6127): 1572-1578. DOI:10.1126/science.1227279 |
| [12] | Behzad H, Gojobori T, Mineta K. Challenges and opportunities of airborne metagenomics[J]. Genome Biology and Evolution, 2015, 7(5): 1216-1226. DOI:10.1093/gbe/evv064 |
| [13] |
陈盟, 祁建城, 杜耀华, 等. 活/死菌检测方法的研究进展[J]. 军事医学, 2018, 42(9): 715-720. Chen M, Qi J C, Du Y H, et al. Research progress in methods for live/dead detection of bacteria[J]. Military Medical Sciences, 2018, 42(9): 715-720. |
| [14] |
唐倩倩, 王剑平, 叶尊忠, 等. 免疫磁分离技术在E. coli O157∶H7检测中的应用[J]. 光谱学与光谱分析, 2009, 29(10): 2614-2618. Tang Q Q, Wang J P, Ye Z Z, et al. Application of immunomagnetic separation to E.coli O157∶H7 detection[J]. Spectroscopy and Spectral Analysis, 2009, 29(10): 2614-2618. DOI:10.3964/j.issn.1000-0593(2009)10-2614-05 |
| [15] |
王晓丹, 赵新华. 配水管网细菌活性影响因素[J]. 环境科学, 2017, 38(2): 622-628. Wang X D, Zhao X H. Influencing factors of bacterial activity in water distribution networks[J]. Environmental Science, 2017, 38(2): 622-628. DOI:10.13227/j.hjkx.201604029 |
| [16] | Haglund A L, Lantz P, Törnblom E, et al. Depth distribution of active bacteria and bacterial activity in lake sediment[J]. FEMS Microbiology Ecology, 2003, 46(1): 31-38. DOI:10.1016/S0168-6496(03)00190-9 |
| [17] | Nazarenko Y, Zhen H J, Han T, et al. Nanomaterial inhalation exposure from nanotechnology-based cosmetic powders: a quantitative assessment[J]. Journal of Nanoparticle Research, 2012, 14(11). DOI:10.1007/s11051-012-1229-2 |
| [18] | Murata K, Zhang D Z. Transport of bacterial cells toward the Pacific in Northern Hemisphere westerly winds[J]. Atmospheric Environment, 2014, 87: 138-145. DOI:10.1016/j.atmosenv.2013.12.038 |
| [19] | Zhen Q, Deng Y, Wang Y Q, et al. Meteorological factors had more impact on airborne bacterial communities than air pollutants[J]. Science of the Total Environment, 2017, 601-602: 703-712. DOI:10.1016/j.scitotenv.2017.05.049 |
| [20] | Gong J, Qi J H, Beibei E, et al. Concentration, viability and size distribution of bacteria in atmospheric bioaerosols under different types of pollution[J]. Environmental Pollution, 2020, 257. DOI:10.1016/j.envpol.2019.113485 |
| [21] | Yin Y D, Qi J H, Gong J, et al. Distribution of bacterial concentration and viability in atmospheric aerosols under various weather conditions in the coastal region of China[J]. Science of the Total Environment, 2021, 795. DOI:10.1016/j.scitotenv.2021.148713 |
| [22] | Bowers R M, McCubbin I B, Hallar A G, et al. Seasonal variability in airborne bacterial communities at a high-elevation site[J]. Atmospheric Environment, 2012, 50: 41-49. DOI:10.1016/j.atmosenv.2012.01.005 |
| [23] | Fang Z G, Ouyang Z Y, Zheng H, et al. Culturable airborne bacteria in outdoor environments in Beijing, China[J]. Microbial Ecology, 2007, 54(3): 487-496. DOI:10.1007/s00248-007-9216-3 |
| [24] | Wang W F, Ma Y T, Ma X, et al. Seasonal variations of airborne bacteria in the Mogao Grottoes, Dunhuang, China[J]. International Biodeterioration & Biodegradation, 2010, 64(4): 309-315. |
| [25] | Genitsaris S, Stefanidou N, Katsiapi M, et al. Variability of airborne bacteria in an urban Mediterranean area (Thessaloniki, Greece)[J]. Atmospheric Environment, 2017, 157: 101-110. DOI:10.1016/j.atmosenv.2017.03.018 |
| [26] | Li M F, Qi J H, Zhang H D, et al. Concentration and size distribution of bioaerosols in an outdoor environment in the Qingdao coastal region[J]. Science of the Total Environment, 2011, 409(19): 3812-3819. DOI:10.1016/j.scitotenv.2011.06.001015 |
| [27] | Park J, Yoon C, Lee K. Comparison of modeled estimates of inhalation exposure to aerosols during use of consumer spray products[J]. International Journal of Hygiene and Environmental Health, 2018, 221(6): 941-950. DOI:10.1016/j.ijheh.2018.05.005 |
| [28] | Hara K, Zhang D Z. Bacterial abundance and viability in long-range transported dust[J]. Atmospheric Environment, 2012, 47: 20-25. DOI:10.1016/j.atmosenv.2011.11.050 |
| [29] | Lighthart B, Shaffer B T. Viable bacterial aerosol particle size distributions in the midsummer atmosphere at an isolated location in the high desert chaparral[J]. Aerobiologia, 1995, 11(1): 19-25. DOI:10.1007/BF02136140 |
| [30] | Fang Z G, Ouyang Z Y, Zheng H, et al. Concentration and size distribution of culturable airborne microorganisms in outdoor environments in Beijing, China[J]. Aerosol Science and Technology, 2008, 42(5): 325-334. DOI:10.1080/02786820802068657 |
| [31] | Gao M, Jia R Z, Qiu T L, et al. Seasonal size distribution of airborne culturable bacteria and fungi and preliminary estimation of their deposition in human lungs during non-haze and haze days[J]. Atmospheric Environment, 2015, 118: 203-210. DOI:10.1016/j.atmosenv.2015.08.004 |
| [32] |
李婉欣, 路瑞, 谢铮胜, 等. 西安市秋冬季不同空气质量下可培养微生物气溶胶浓度和粒径分布[J]. 环境科学, 2017, 38(11): 4494-4500. Li W X, Lu R, Xie Z S, et al. Concentration and size distribution characteristics of culturable bioaerosols at various air quality levels during fall and winter in Xi'an, China[J]. Environmental Science, 2017, 38(11): 4494-4500. DOI:10.13227/j.hjkx.201703072 |
| [33] | Dong L J, Qi J H, Shao C C, et al. Concentration and size distribution of total airborne microbes in hazy and foggy weather[J]. Science of the Total Environment, 2016, 541: 1011-1018. DOI:10.1016/j.scitotenv.2015.10.001 |
| [34] | Wang Y Q, Zhang X Y, Draxler R R. TrajStat: GIS-based software that uses various trajectory statistical analysis methods to identify potential sources from long-term air pollution measurement data[J]. Environmental Modelling & Software, 2009, 24(8): 938-939. |
| [35] | Gandolfi I, Bertolini V, Bestetti G, et al. Spatio-temporal variability of airborne bacterial communities and their correlation with particulate matter chemical composition across two urban areas[J]. Applied Microbiology and Biotechnology, 2015, 99(11): 4867-4877. DOI:10.1007/s00253-014-6348-5 |
| [36] | Smets W, Moretti S, Denys S, et al. Airborne bacteria in the atmosphere: presence, purpose, and potential[J]. Atmospheric Environment, 2016, 139: 214-221. DOI:10.1016/j.atmosenv.2016.05.038 |
| [37] |
宫静, 祁建华, 李鸿涛. 青岛近海生物气溶胶中总微生物的分布特征[J]. 环境科学, 2019, 40(8): 3477-3488. Gong J, Qi J H, Li H T. Distribution of total microbes in atmospheric bioaerosols in the coastal region of Qingdao[J]. Environmental Science, 2019, 40(8): 3477-3488. DOI:10.13227/j.hjkx.201901262 |
| [38] | Mu F F, Li Y P, Lu R, et al. Source identification of airborne bacteria in the mountainous area and the urban areas[J]. Atmospheric Research, 2020, 231. DOI:10.1016/j.atmosres.2019.104676 |
| [39] | Qi J H, Liu X H, Yao X H, et al. The concentration, source and deposition flux of ammonium and nitrate in atmospheric particles during dust events at a coastal site in northern China[J]. Atmospheric Chemistry and Physics, 2018, 18(2): 571-586. DOI:10.5194/acp-18-571-2018 |
| [40] | Ehrlich R, Miller S. Effect of NO2 on airborne venezuelan equine encephalomyelitis virus[J]. Applied Microbiology, 1972, 23(3): 481-484. DOI:10.1128/am.23.3.481-484.1972 |
| [41] | Hameed A A A, Khoder M I, Ibrahim Y H, et al. Study on some factors affecting survivability of airborne fungi[J]. Science of the Total Environment, 2012, 414: 696-700. DOI:10.1016/j.scitotenv.2011.10.042 |
| [42] | Pan Y L, Kalume A, Wang C J, et al. Atmospheric aging processes of bioaerosols under laboratory-controlled conditions: a review[J]. Journal of Aerosol Science, 2021, 155. DOI:10.1016/j.jaerosci.2021.105767 |
| [43] | Guo K Q, Qian H, Ye J, et al. Assessment of airborne bacteria and fungi in different-type buildings in Nanjing, a hot summer and cold winter moist Chinese city[J]. Building and Environment, 2021, 205. DOI:10.1016/j.buildenv.2021.108258 |
| [44] | Tsai F C, Macher J M. Concentrations of airborne culturable bacteria in 100 large US office buildings from the BASE study[J]. Indoor Air, 2005, 15(S9): 71-81. DOI:10.1111/j.1600-0668.2005.00346.x |
| [45] | Xie Z S, Fan C L, Lu R, et al. Characteristics of ambient bioaerosols during haze episodes in China: a review[J]. Environmental Pollution, 2018, 243: 1930-1942. DOI:10.1016/j.envpol.2018.09.051 |
| [46] | Zhong X, Qi J H, Li H T, et al. Seasonal distribution of microbial activity in bioaerosols in the outdoor environment of the Qingdao coastal region[J]. Atmospheric Environment, 2016, 140: 506-513. DOI:10.1016/j.atmosenv.2016.06.034 |
| [47] | Bowers R M, Clements N, Emerson J B, et al. Seasonal variability in bacterial and fungal diversity of the Near-Surface atmosphere[J]. Environmental Science & Technology, 2013, 47(21): 12097-12106. |
| [48] | Ji L, Zhang Q, Fu X W, et al. Feedback of airborne bacterial consortia to haze pollution with different PM2.5 levels in typical mountainous terrain of Jinan, China[J]. Science of the Total Environment, 2019, 695. DOI:10.1016/j.scitotenv.2019.133912 |
| [49] | Tang J W. The effect of environmental parameters on the survival of airborne infectious agents[J]. Journal of the Royal Society Interface, 2009, 6(S6): 737-746. |
| [50] | Jeon E M, Kim H J, Jung K, et al. Impact of Asian dust events on airborne bacterial community assessed by molecular analyses[J]. Atmospheric Environment, 2011, 45(25): 4313-4321. DOI:10.1016/j.atmosenv.2010.11.054 |
| [51] | Zhang Y Y, Lang J L, Cheng S Y, et al. Chemical composition and sources of PM1 and PM2.5 in Beijing in autumn[J]. Science of the Total Environment, 2018, 630: 72-82. DOI:10.1016/j.scitotenv.2018.02.151 |
| [52] | Xu C H, Wei M, Chen J M, et al. Bacterial characterization in ambient submicron particles during severe haze episodes at Ji'nan, China[J]. Science of the Total Environment, 2017, 580: 188-196. DOI:10.1016/j.scitotenv.2016.11.145 |
| [53] |
雒月云, 毛怡心, 庄思琪, 等. 北京市不同季节室外细菌气溶胶分布特征及其环境影响因素分析[J]. 环境科学研究, 2022, 35(2): 556-565. Luo Y Y, Mao Y X, Zhuang S Q, et al. Seasonal distribution characteristics of bacterial aerosols and their correlations with environmental factors in Beijing, China[J]. Research of Environmental Sciences, 2022, 35(2): 556-565. DOI:10.13198/j.issn.1001-6929.2021.12.02 |
| [54] | Wang Y F, Wang C H, Hsu K L. Size and seasonal distributions of airborne bioaerosols in commuting trains[J]. Atmospheric Environment, 2010, 44(35): 4331-4338. DOI:10.1016/j.atmosenv.2010.08.029 |
| [55] |
袁刚, 祁建华, 丁雪. 青岛近海不同污染过程下大气颗粒态氮磷浓度分布特征[J]. 环境科学, 2021, 42(3): 1280-1297. Yuan G, Qi J H, Ding X. Concentrations and patterns of atmospheric particulate nitrogen and phosphorus during different weather conditions in Qingdao coastal region[J]. Environmental Science, 2021, 42(3): 1280-1297. DOI:10.13227/j.hjkx.202007241 |
| [56] | Zhang X J, Musson-Genon L, Dupont E, et al. On the Influence of a simple microphysics parametrization on radiation fog modelling: a case study during ParisFog[J]. Boundary-Layer Meteorology, 2014, 151(2): 293-315. DOI:10.1007/s10546-013-9894-y |
| [57] | Wei M, Xu C H, Xu X M, et al. Size distribution of bioaerosols from biomass burning emissions: characteristics of bacterial and fungal communities in submicron (PM1) and fine (PM2.5) particles[J]. Ecotoxicology and Environmental Safety, 2019, 171: 37-46. DOI:10.1016/j.ecoenv.2018.12.026 |
| [58] | QX/T 113-2010, 霾的观测和预报等级[S]. |
| [59] | Betha R, Behera S N, Balasubramanian R. 2013 Southeast Asian smoke haze: fractionation of particulate-bound elements and associated health risk[J]. Environmental Science & Technology, 2014, 48(8): 4327-4335. |
| [60] | Zhai Y B, Li X, Wang T F, et al. A review on airborne microorganisms in particulate matters: composition, characteristics and influence factors[J]. Environment International, 2018, 113: 74-90. DOI:10.1016/j.envint.2018.01.007 |
| [61] | Zheng B, Tong D, Li M, et al. Trends in China's anthropogenic emissions since 2010 as the consequence of clean air actions[J]. Atmospheric Chemistry and Physics, 2018, 18(19): 14095-14111. DOI:10.5194/acp-18-14095-2018 |
| [62] | Dueker M E, O'Mullan G D, Weathers K C, et al. Coupling of fog and marine microbial content in the near-shore coastal environment[J]. Biogeosciences, 2012, 9(2): 803-813. DOI:10.5194/bg-9-803-2012 |
| [63] | Theunissen H J, Toom N A L, Burggraaf A, et al. Influence of temperature and relative humidity on the survival of Chlamydia pneumoniae in aerosols[J]. Applied and Environmental Microbiology, 1993, 59(8): 2589-2593. DOI:10.1128/aem.59.8.2589-2593.1993 |
| [64] | Rajput P, Singh D K, Singh A K, et al. Chemical composition and source-apportionment of sub-micron particles during wintertime over Northern India: new insights on influence of fog-processing[J]. Environmental Pollution, 2018, 233: 81-91. DOI:10.1016/j.envpol.2017.10.036 |
| [65] | Mazar Y, Cytryn E, Erel Y, et al. Effect of dust storms on the atmospheric microbiome in the eastern mediterranean[J]. Environmental Science & Technology, 2016, 50(8): 4194-4202. |
| [66] |
汪瑶, 祁建华, 张婷, 等. 青岛多种天气下生物气溶胶中细菌群落特征[J]. 中国海洋大学学报(自然科学版), 2021, 51(6): 88-98. Wang Y, Qi J H, Zhang T, et al. Bacterial community characteristics in different weather conditions of atmospheric bioaerosols in Qingdao[J]. Periodical of Ocean University of China, 2021, 51(6): 88-98. DOI:10.16441/j.cnki.hdxb.20200135 |
| [67] | Amarloei A, Fazlzadeh M, Jafari A J, et al. Particulate matters and bioaerosols during Middle East dust storms events in Ilam, Iran[J]. Microchemical Journal, 2020, 152. DOI:10.1016/j.microc.2019.104280 |