 2022, Vol. 43
2022, Vol. 43

2. 中国科学院生态环境研究中心, 北京 100085;
3. 东北林业大学林学院, 哈尔滨 150040
 , WANG En-rui1 , CHANG Sheng1
, WANG En-rui1 , CHANG Sheng1 , WANG Shan-jun1 , JIN De-cai2 , FAN Yue-ting1 , ZHANG Kun-feng1,3 , XIE Qiong1 , FU Qing1
, WANG Shan-jun1 , JIN De-cai2 , FAN Yue-ting1 , ZHANG Kun-feng1,3 , XIE Qiong1 , FU Qing1
2. Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. School of Forestry, Northeast Forestry University, Harbin 150040, China
浮游植物是水环境中的初级生产者和食物链的基础, 它们分布广泛, 在生态系统的能量流动和物质循环中发挥着重要作用[1].浮游植物群落与河流和湖泊的水环境因子之间会互相影响, 由于浮游植物体积小、结构简单及对生境变化敏感, 因此其群落结构的变化可以在一定程度上反映环境因素的变化[2, 3], 对环境条件的变化具有指示作用, 同时环境条件的改变也直接或间接地影响到浮游植物的群落结构[4].
有研究表明, 水温、光照、营养盐和高锰酸盐指数是水体中浮游藻类的主要影响因子[5~8], 如郑丙辉等[7]的研究发现氮磷营养盐是大宁河水华敏感期的浮游植物的主要影响因子, 马沛明等[8]的研究发现水力停留时间和温度对浙江汤浦水库的藻类间演变和水华暴发起到主导作用.然而, 已有研究主要集中在单个湖泊或河道[7, 8], 对于多个水库群, 尤其是河道输水、库间调水的连通型水库群之间浮游植物群落结构和环境驱动因子的研究却鲜见报道.由于水库受河水输入影响, 容易导入大量的营养盐, 同时也因输水引发水库水动力的改变[9], 库区生境稳定性被打破, 造成底层藻类上浮.谢飞[10]曾对珠海市18座中营养水平以上的水库进行富营养化和浮游植物群落特征的研究, 对比调水型和非调水型水库的营养状态水平, 结果表明调水型水库的富营养化水平显著高于非调水型水库, 说明持续调水对水库的富营养化状态有直接影响.因此, 研究连通型调水水库群的浮游植物群落结构和环境因子的相关性、探究输水河道营养盐对水库浮游植物群落结构的影响具有重要意义.
珠海市位于珠江口西岸, 濒临南海, 地处北纬21°48′~22°27′、东经113°03′~114°19′之间, 珠海市各水库通过管网、泵站与河流相连, 形成了有特色的江库连通的调水水库系统[11], 而在咸潮时期, 由于靠近出海口, 近旁水道无法取水时, 自来水厂会在水库直接取水, 且水库为规避咸潮提前蓄水, 决定了珠海的供水系统为多取水点和多水库连调的水源特点(图 1)[12], 形成了江河输水、江库连通和库库连通的独特布局.

|
图 1 输水路径示意 Fig. 1 Water diversion route |
珠海市饮用水源也是澳门的主要供水水源, 其水质保障和饮水安全直接关乎粤港大湾区的社会稳定.本文选择珠海市4座连通型水库(大镜山水库、杨寮水库、竹仙洞水库和竹银水库)和输水河道磨刀门水道作为研究对象, 其中磨刀门水道是珠海及澳门的重要水源地, 枯水期咸潮入侵有威胁当地供水安全的风险[13], 4座水库为珠海市的骨干水源水库群, 主要承担储蓄河水和调配供水的任务, 虽然通过水量调度加大枯水期西江下游来水量, 对于抑制磨刀门水道的咸潮上溯效果显著[14, 15], 但另一方面将河流的营养物质引入水库, 增加了水库富营养化的风险[16].本研究分析了丰、枯两季的浮游植物分布特征和水环境质量, 探讨了浮游植物群落特征和环境驱动因子的关系, 以期为珠海市在水源水质安全保障和水环境管理提供基础资料和科学依据.
1 材料与方法 1.1 研究区概况珠海市在用的集中式饮用水水源地中, 大镜山水库(DJS)、杨寮水库(YL)、竹仙洞水库(ZXD)和竹银水库(ZY)属于从磨刀门水道取水的中部城区供水系统, 且相互连通、库间调水(图 1).其调水方式为:丰水期(4~9月)抽取西江磨刀门水道原水进入大镜山、竹仙洞和竹银水库, 杨寮水库水源由大镜山水库经正坑, 坑尾两个小型水库调入.枯水期(10月~次年3月, 调水主要集中于10~12月)通过从磨刀门水道抢淡, 输往最近的竹银水库, 再经竹银水库调水至竹仙洞和大镜山水库. 4个水库的基本情况见表 1.
|
|
表 1 各水库基本情况1) Table 1 Basic information of reservoirs |
由表 1可见, 4个水库集雨面积小、水库自产水远远无法满足供水需求.水库水力停留时间较短, 属于不稳定湖泊, 或介于湖泊与河流之间、或类似于河流, 库内生境并不稳定.
采样时间为2021年8月和10月, 共布设24个采样点(图 2), 在大镜山水库设置5个采样点(DJS1~DJS5), 杨寮水库设置9个采样点(YL1~YL9), 竹仙洞水库设置3个采样点(ZXD1~ZXD3), 竹银水库设置4个采样点(ZY1~ZY4).各采样点充分覆盖4个水库水体, 代表性强, 并覆盖了地方监测部门选定的日常监测点位, 其中DJS1、YL1、ZXD1和ZY1分别为饮用水水源取水口.

|
图 2 采样点分布示意 Fig. 2 Distributions of sampling sites |
环境因子共12项, 包括水温(WT)、酸碱度(pH)、透明度(SD)、溶解氧(DO)、高锰酸盐指数、氨氮(NH4+-N)、硝酸盐(NO3-)、总磷(TP)、总氮(TN)、叶绿素a(Chl-a)、氯化物(Cl-)和总有机碳(TOC).样品分析方法参照文献[17].
浮游植物定性样品使用25号(64 μm)浮游生物网在水面成“∞”状巡回拖动3~5次, 然后打开浮游生物网的旋塞, 让网内浓缩水样流入50 mL圆形广口瓶中, 用1% ~1.5%样品体积的鲁哥氏液摇匀固定.使用光学显微镜鉴定拍照, 鉴定方法参考文献[18].定量样品使用采水器与水深0.5 m处采集1 L水样, 加入10 mL鲁哥氏液摇匀固定, 带回实验室静置48 h后浓缩至50 mL, 然后镜检计数.
1.3 数据处理本文应用Origin 2018和Excel 2016软件对数据进行统计、作图和分析; 使用ArcGIS软件对水库采样点分布进行绘制.采用Canoco5.0软件对物种数据和除WT、pH、DO和SD之外的环境数据进行lg(x+1)标准化处理, 去趋势分析(DCA)得出浮游植物丰、枯两季的第一轴长分别为1.6与1.4, 长度梯度均小于4, 因此都选用冗余分析(redundancy analysis, RDA)分析[19, 20].单因素方差分析(ANOVA)和非参数检验分析采用SPSS26.0完成.主坐标分析(PCoA)和相似性分析(ANOSIM)用R语言.
浮游植物群落特征指标采用Shannon-Wiener多样性指数、Margalef物种丰富度指数、Pielou均匀度指数、优势度、生物丰度和种类数比例进行分析[21].RDA分析中物种数据经过筛选, 本文用于排序的物种均为4个水库的优势种.各指数的计算公式如下.
Shannon-Wiener多样性指数(H′):
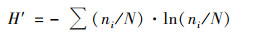
|
(1) |
Margalef物种丰富度指数(D):
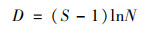
|
(2) |
Pielou均匀度指数(J):
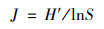
|
(3) |
优势度(Y):
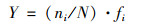
|
(4) |
式中, ni为第i个物种的个体总数, N为样品的总个体数, S为样品的种类总数, fi为样本个体的出现率, Y>0.02的物种认定为优势种.
2 结果与分析 2.1 环境因子4个水库各采样点的环境因子及相关性分析如表 2和表 3所示.监测结果显示, 两季各水库的温度变化不大, 在28.215~32.267℃之间, pH值总体为中性偏碱, 在7.239~8.831之间, 溶解氧均达到Ⅱ类水标准, 根据TLI分级标准, 这4个水库处于中营养状态.高锰酸盐指数、TP、TN、NH4+-N、NO3-、Cl-和TOC数据均满足正态分布, 对各指标做T检验齐性分析, 方差显示齐的指标进行ANOVA分析, 方差不齐的指标进行非参数检验分析.
|
|
表 2 各水库和磨刀门水道的水体理化因子均值1) Table 2 Mean value of water physical and chemical parameters in reservoirs and Modaomen waterway |
|
|
表 3 各水库和磨刀门水道重点环境因子差异性1) Table 3 Differences in mainly environmental factors in reservoirs and Modaomen Waterway |
丰水期, 大镜山水库与磨刀门水道的NH4+-N浓度水平相当, 竹仙洞水库与磨刀门水道的TN、NH4+-N和TOC浓度水平相当, 杨寮、竹银水库和磨刀门水道的NH4+-N和高锰酸盐指数较为接近, 而各水库与磨刀门水道的TP、NO3-和Cl-浓度均表现出显著差异(P < 0.05或P < 0.01); 各水库间除了杨寮和竹银水库的环境因子较为接近, 其他水库NO3-浓度表现出极为显著的差异(P < 0.01).
枯水期, 大镜山水库与磨刀门水道的TN、NH4+-N、NO3-和TOC浓度较为接近, 杨寮水库和磨刀门水道的高锰酸盐指数较为接近, 竹仙洞水库和磨刀门水道的高锰酸盐指数和NO3-浓度较为接近, 竹银水库和磨刀门水道除Cl-之外的其余主要环境因子浓度均较为接近, 而各水库和磨刀门水道的Cl-浓度均表现出显著差异(P<0.05或P<0.01), 且磨刀门水道的Cl-浓度远高于4个水库; 各水库之间除了竹仙洞和竹银水库的环境因子较为接近, 其他水库相互之间的TN、NH4+-N和TOC浓度均表现出显著差异(P<0.05或P<0.01).
总体来看, 在丰水期, 经过水库一段时间的滞留, 各水库库内水环境状况逐渐趋于稳定, 不仅表现出与原水较为明显的差异, 也呈现出一定的库间差异, 其中杨寮水库尤为明显, 虽然其来水源自大镜山水库输水, 但由于途经2座其他水库, 且水力停留时间不同, 杨寮水库和大镜山水库的环境因子依然差异较大.在枯水期, 磨刀门水道主要向竹银水库输水, 竹银水库向其他水库输水, 因此, 竹银水库的环境因子和磨刀门水道最为接近; 因咸潮上溯, 磨刀门水道枯水期Cl-浓度剧增, 但4个水库的Cl-浓度却与磨刀门水道表现出显著差异, 且与丰水期浓度相差不大, 分析认为, 这是由于水库库内自身具有水环境调节功能, 另一方面, 磨刀门水道高咸水集中在下游地区, 而枯水期以抢淡为主, 水库水质主要受上游平岗泵站调水影响.
2.2 浮游植物群落结构 2.2.1 浮游植物组成丰、枯两季4个水库共检测出8门73属150种浮游植物, 绿藻、硅藻、蓝藻和甲藻这4个门类占比重较大.其中丰水期检测出8门62属118种, 绿藻门的种类最为丰富, 为59种, 占总藻种数的50.0%, 其次是蓝藻门18种和硅藻门17种, 分别占15.2%和14.4%; 枯水期检测出8门61属104种, 绿藻门61种, 占总藻种数的58.6%, 其次为硅藻门16种和蓝藻门14种, 分别占15.3%和13.4%.
对比丰枯两季浮游植物的种类数占比(图 3), 可以发现4个水库的浮游植物种类占比趋于相似, 主要以绿藻和蓝藻为主.

|
图 3 各水库浮游植物种类数占比 Fig. 3 Composition of phytoplankton species in reservoirs |
丰水期, 大镜山和杨寮水库的浮游植物丰度占比极为相似, 蓝藻门均占绝对优势, 竹仙洞以蓝藻门为主, 其次为隐藻门, 竹银水库以绿藻门主, 其次为蓝藻门和硅藻门; 枯水期, 大镜山水库以蓝藻门为主, 杨寮水库以蓝藻门为主, 硅藻门和绿藻门次之, 竹仙洞水库以绿藻门为主, 其次为硅藻门和黄藻门, 竹银水库以绿藻门为主, 其次为硅藻门(图 4).

|
图 4 水库浮游植物相对丰度 Fig. 4 Relative abundance of plankton in reservoirs |
从浮游植物细胞丰度的空间分布来看(图 5), 4个水库浮游植物细胞丰度出现显著差异, 大镜山和杨寮水库的浮游植物细胞密度显著高于竹仙洞和竹银水库, 可见换水更为频繁的竹仙洞和竹银水库, 水体中浮游植物各种类丰度占比更均衡, 且因水力扰动大, 浮游植物细胞丰度也偏低.

|
图 5 水库浮游植物丰度空间分布 Fig. 5 Spatial distribution of phytoplankton abundance in reservoirs |
4个水库浮游植物优势种共6门22种, 主要为蓝藻门、绿藻门、硅藻门和隐藻门.其中大镜山和杨寮水库较为相似, 共有优势种为蓝藻门拟柱孢藻(Cylindrospermopsis sp.), 其优势度为0.89~0.96, 此外大镜山水库的特有优势种为蓝藻门尖头藻(Raphidiopsis sp.).竹仙洞和竹银水库较为相似, 优势种多为蓝藻门、绿藻门和硅藻门, 另外隐藻门、甲藻门和金藻门也为占优门类, 竹仙洞水库蓝藻门伪鱼腥藻(Pseudoanabaena sp.)优势度最大, 为0.84, 竹银水库优势度最大的是硅藻门的小环藻(Cyclotella sp.), 为0.39.由表 4中可见, 竹仙洞和竹银水库的优势种分布较大镜山和杨寮水库更为丰富.
|
|
表 4 不同水库浮游植物优势种及优势度1) Table 4 Dominant species and dominance index of plankton in different lakes |
2.2.4 浮游植物多样性分析
为探究4个水库的库内浮游植物多样性, 使用Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数对4个水库丰、枯两季进行α多样性分析(图 6).结果显示, 丰水期, 从多样性和均匀度指标表征上来看, 大镜山和杨寮水库都处于重污染型, 竹仙洞水库为重污染向α-中污染过渡型, 或重污染向α-中污染、β-中污染过渡型, 而竹银水库则处于β-中污染或无污染的清洁水体状态, 从丰富度指数看, 所有水库均处于α-中污染水体, 可见丰水期水质最好的是竹银水库, 基本为中污染或无污染, 其次为竹仙洞水库, 基本处于中污染状态, 而大镜山和杨寮水库水质差于竹仙洞和竹银水库, 处于重污染或中污染状态.枯水期, 从多样性和均匀度指标表征上来看, 大镜山处于重污染型, 杨寮水库处于重污染向中污染过渡型, 竹仙洞和竹银水库为β-中污染向清洁水体过渡型, 从丰富度指数看, 除了竹仙洞为重污染型, 其他3个水库均为中污染型.简言之, 竹银水库的浮游植物多样性最丰富, 指示水体最优, 竹仙洞水库次之, 杨寮水库再次之, 而4个水库中大镜山水库浮游植物多样性最差.

|
图 6 各水库浮游植物生物多样性指数 Fig. 6 Biodiversity index of phytoplankton in reservoirs |
为了解4个水库浮游植物群落结构的库间差异, PCoA分析(图 7)结果显示第一主成分(PCoA1)和第二主成分(PCoA2)的差异解释率在丰、枯两季分别为41.79%、21.72%和47.01%、19.72%, 前两轴在丰枯两季可以分别解释4个水库所有差异的63.51%和66.73%, 说明PCoA能够较好地反映4个水库浮游植物群落的β多样性.从降维后的样方分布可见, 大镜山和杨寮水库在丰、枯两季均有部分点位距离较近, 且都分别与竹仙洞和竹银水库距离较远, 竹仙洞和竹银水库两者之间在丰、枯两季距离都比较远.

|
图 7 水库浮游植物群落PCoA分析 Fig. 7 PCoA of phytoplankton in reservoirs |
为进一步验证水库间浮游植物群落的差异显著性, 对各水库浮游植物物种数据采用基于Bray-Curtis距离的ANOSIM分析方法.结果表明, 无论丰、枯两季, 大镜山和杨寮水库的差异性均不显著(P>0.05), 而大镜山和杨寮水库分别与竹仙洞和竹银水库两两之间存在显著差异(P<0.05).这说明大镜山和杨寮水库之间的调水使库内浮游植物群落结构趋于接近, 而竹仙洞和竹银水库则呈现出各自不同的浮游植物群落特征(表 5).
|
|
表 5 浮游植物群落Bray-Curtis距离的ANOSIM检验 Table 5 ANOSIM test of phytoplankton community based on Bray-Curtis distance |
2.3 浮游植物与环境因子间的关系
分别对4个水库丰枯两季的浮游植物优势种和环境因子进行RDA分析, 结果如图 8所示.丰水期, 第一轴与第二轴分别能解释29.22%和19.55%的浮游植物群落差异, 大镜山和杨寮水库样方聚集于第二、三象限, 与TOC靠近, 而竹仙洞样方主要聚集于第四象限, 与NO3-和TP靠近, 竹银水库样方距离环境因子较远; 枯水期, 第一轴与第二轴分别能解释31.79%和14.51%的浮游植物群落差异, 竹仙洞和大镜山水库样方分别聚集于第一、二象限, 且与环境因子较远, 杨寮水库样方聚集于第二、三象限, 与TOC、高锰酸盐指数、Chl-a和NH4+-N靠近, 竹银水库样方聚集于第四象限, 与NO3-、TN和Cl-靠近.

|
蓝色箭头表示物种, 编码z1~z22对应的物种名见表 4; 红色箭头表示环境因子; 点状图形表示不同水库的样方分布 图 8 浮游植物群落与环境因子的RDA分析 Fig. 8 RDA plot of plankton community and environmental factors |
通过蒙特卡洛置换检验(表 6), 可以看出丰水期NO3-、TOC和TP与浮游植物群落特征具有显著相关性(P<0.05), 且在第一序轴NO3-和TP呈正相关, TOC呈负相关, 在第二序轴所有环境因子都呈负相关; 枯水期NO3-、高锰酸盐指数、Cl-和NH4+-N与浮游植物群落特征具有显著相关性(P<0.05), 且在第一序轴NO3-和Cl-呈正相关, 高锰酸盐指数和NH4+-N呈负相关, 在第二序轴除NO3-呈正相关外, 其余环境因子都呈负相关.
|
|
表 6 各环境因子与浮游植物群落的相关性1) Table 6 Correlations of each environmental factor and phytoplankton communities |
3 讨论 3.1 河库和库间调水特征对浮游植物群落结构的影响
有研究表明, 对于亚热带地区来说, 水动力学特征是影响浮游植物群落结构的重要因素[22].本文分析显示, 大镜山和杨寮水库浮游植物均以蓝藻为主, 而竹仙洞和竹银水库表现为蓝藻、绿藻和硅藻共同占优的特征, 这与已有研究对珠三角西江水系浮游生物调查结果类似[10, 11], 其水力停留时间的差异性是主要原因.水力停留时间是集中反映水体动力学变化过程的重要指标, 也是反映平流损失率的重要参数[23], 一般认为只有当水力停留时间超过14 d时[24], 平流损失率低于浮游植物生长率, 才有利于浮游植物的逐渐积累.水力停留时间也是影响蓝藻生长的关键因素之一[25, 26], 这是因为调水水库中, 水体稳定性往往受到水力扰动较大, 尤其是具有漂浮机制的蓝藻, 调水带来的扰动对分布在水体中层的绿藻和硅藻影响较小.水力停留时间长则意味着水库具有相对稳定的环境, 大镜山水库为营养型水库(326.8 d), 换水周期较长加之珠海终年水温较高, 有利于蓝藻的稳定生长, 而蓝藻门在大量增长时会分泌他感物质[27, 28], 改变周围pH及浊度, 进而抑制其他藻类的生长[29].黄洪波[30]指出水力停留时间是影响大镜山水库浮游植物生物丰度的关键因子, 本文的研究结果也证实了这点.杨寮水库为过渡型水库(111.6 d), 竹仙洞水库为过流型水库(6.8 d), 邹红菊[11]发现水力停留时间长的杨寮水库蓝藻优势极高, 而水力停留时间短的竹仙洞水库则为蓝藻、绿藻和硅藻共同占优, 与本研究的结果一致, 这主要是因为竹仙洞水库水力停留时间仅有不到7 d, 水体一般不分层, 水体混合性好, 不利于在稳定分层水体中占优势的蓝藻生存[31, 32].竹银水库属于过渡型水库(93.9 d), 一般认为大于100 d的水力停留时间有利于水体分层, 竹银水库的库内水体相对混合性较好.郭川等[33]对竹银水库的健康评价认为, 竹银水库以蓝藻、硅藻和绿藻为优势类群, 其水华警报预警为低风险.本文的研究结果也揭示了水力停留时间较长的湖泊型水库中蓝藻多为主要优势类群, 而类似于河流或介于河流与湖泊之间的水库浮游植物群落结构更加趋于多种类群共同占优.
4个水库的浮游植物群落相似性及差异性分析表明, 大镜山和杨寮水库群落特征较为相似, 且分别与竹仙洞和竹银水库存在两两之间的显著差异.这主要是因为杨寮水库的水源来自大镜山水库输水, 且换水周期长, 受外源输入扰动的影响不大, 因此杨寮水库表现出与大镜山水库较为接近的类群特征.大镜山水库、竹仙洞水库和竹银水库则分别从磨刀门水道输水, 磨刀门水道上下游水质差别较大, 加之输水路线不同, 导致理化因子存在显著差异(表 3), 竹仙洞和竹银水库各自换水周期较短, 且在枯水期通过竹仙洞水库抢淡输水, 外源输入扰动频繁, 因此水流动力造成的生境异质性是水库之间浮游群落差异性的主要原因.
3.2 各水库水质生物学评价分析各水库理化性质, 以总磷作为评价因子, 大镜山水库、杨寮水库和竹仙洞水库均为Ⅲ类水质, 而竹银水库为Ⅲ~Ⅳ类水质; 以总氮作为评价因子, 大镜山水库和杨寮水库(由于有异常点, 取均值评价)为Ⅲ~Ⅳ类水质, 竹仙洞水库为Ⅳ~Ⅴ类水质, 竹银水库为Ⅲ~Ⅴ类水质(图 9).如以其他水质指标评价, 4个水库的水质均达到地表水水环境质量Ⅲ类或Ⅱ类水质标准.对比水质生物学评价结果(图 6), 多样性指数和均匀度指数与理化因子(除TP和TN外)指示的水质状况表现一致, 这说明用多样性指数和均匀度指数表征水质更加科学.

|
图 9 磨刀门水道输水点和各水库的总氮和总磷浓度 Fig. 9 Total nitrogen and phosphorus content of water point and each reservoir |
氮磷指标表明的各水库水质状况与生物学水质评价表现出不一致的指示, 这是因为一方面各水库的氮盐和磷盐超标主要由于磨刀门水道的输水(输水点分别为MDM1、MDM4和MDM10), 输水带来的水力扰动影响库内生境的变化, 尤其是竹仙洞水库受到频繁的水力扰动, 导致优势种变化较快.另一方面营养盐对浮游植物的影响主要集中在氮磷的限制上[34], 有研究表明营养盐水平的变化会导致浮游植物群落组成的改变[35], 在一些水体中氮是影响浮游植物生长的限制因子[36, 37], 在另外一些水体中磷为限制因子[1, 2], 当ρ(总磷)超过0.01 mg ·L-1时, 蓝藻的生长很大程度上受物理因子的影响, 蓝藻的生长速度更易受水体稳定性影响[38], 所有水库的ρ(总磷)均超过0.01 mg ·L-1, 这意味着总磷对蓝藻生长的关系不占主导, 而蓝藻生长又涉及到是否抑制其他浮游植物的生长, 因此磷盐的超标并未引发库内浮游植物的多样性变化.当ρ(无机氮盐)超过0.1 mg ·L-1时, 浮游植物生长将不受氮浓度的限制[32, 39], 各水库中ρ(无机氮盐)均超过0.1 mg ·L-1, 因此氮盐的大量输入虽导致水体水质类别恶化, 却无法限制浮游植物的多样性繁殖.
3.3 浮游植物与环境因子的关系有研究表明, 浮游植物群落分布受水体水文条件、物理和化学等诸多因素的影响[40~44].RDA分析显示, 大镜山水库和杨寮水库丰枯两季浮游植物主要优势种均为蓝藻门的拟柱孢藻, 其分布与TOC都呈现出显著正相关(P<0.01), 这是因为TOC代表水体中有机质(DOM)的含量, DOM通过吸收紫外光从而对浮游植物光合作用产生影响[45], 不断消耗的碳酸盐导致水体中碳氮比降低, 而拟柱孢藻具有固氮能力, 在氮利用上被认为是广幅种[46], 高浓度的氮环境拟柱孢藻比其他蓝藻生长更快, 具有较强的竞争优势[47].丰水期, 竹仙洞的浮游植物分布与NO3-和TP都呈现出显著正相关(P<0.01), 这是因为优势种伪鱼腥藻对磷含量有较宽的适应性[48, 49], 竹仙洞水库的浮游植物在枯水期主要受水力扰动影响, 与水库中环境因子相关性不强.丰水期, 竹银水库的浮游植物与环境因子相关性不强, 枯水期优势种主要为绿藻门和硅藻门, 浮游植物群落分布与NO3-和Cl-呈显著正相关, 与TP呈正相关, Zhang等[50]的研究也表明高氮磷比环境下更适合绿藻的繁殖.陈悦[51]对硅藻门的小环藻属研究表明TP是影响硅藻增长速度的重要指标.竹银水库枯水期抢淡调水是造成营养盐输入的主要因素.
4 结论(1) 丰枯两季, 珠海市大镜山水库、杨寮水库、竹仙洞水库和竹银水库群共检测出浮游植物8门73属150种, 以绿藻、蓝藻和硅藻为主. 4个水库种类分布较为趋近, 优势种分布大镜山水库和杨寮水库较为相似, 竹仙洞和竹银水库与前两者各不相同.
(2) 影响4个水库浮游植物多样性的主要因素为磨刀门水道输水带来的水力扰动和水库水力停留时间.大镜山水库水力停留时间最长, 浮游植物多样性最差, 竹仙洞水库水力停留时间最短, 浮游植物优势种分布最广
(3) 水质生物学评价表明, 大镜山水库丰枯两季均处于重污染状态; 杨寮水库丰水期处于重污染状态, 枯水期处于重污染状态; 竹仙洞丰水期处于中污染状态, 枯水期处于寡污-清洁状态; 竹银水库丰枯两季均处于寡污-清洁状态.
(4) 影响4个水库浮游植物群落结构的环境因子为NO3-、TOC、TP、Cl-和NH4+-N.营养盐主要来自磨刀门水道输水带来的外源输入, 改善磨刀门水道水质是提升4个水库水质的关键举措.
| [1] | Manickam N, Bhavan P S, Santhanam P, et al. Phytoplankton biodiversity in the two perennial lakes of Coimbatore, Tamil Nadu, India[J]. Acta Ecologica Sinica, 2020, 40(1): 81-89. DOI:10.1016/j.chnaes.2019.05.014 |
| [2] | Cui L, Lu X X, Dong Y L, et al. Relationship between phytoplankton community succession and environmental parameters in Qinhuangdao coastal areas, China: a region with recurrent brown tide outbreaks[J]. Ecotoxicology and Environmental Safety, 2018, 159: 85-93. DOI:10.1016/j.ecoenv.2018.04.043 |
| [3] | Pérez J R, Loureiro S, Menezes S, et al. Assessment of water quality in the Alqueva Reservoir (Portugal) using bioassays[J]. Environmental Science and Pollution Research, 2010, 17(3): 688-702. DOI:10.1007/s11356-009-0174-9 |
| [4] | Liu C Q, Liu L S, Shen H T. Seasonal variations of phytoplankton community structure in relation to physico-chemical factors in Lake Baiyangdian, China[J]. Procedia Environmental Sciences, 2010, 2: 1622-1631. DOI:10.1016/j.proenv.2010.10.173 |
| [5] |
胡旻琪, 张玉超, 马荣华, 等. 巢湖2016年蓝藻水华时空分布及环境驱动力分析[J]. 环境科学, 2018, 39(11): 4925-4937. Hu M Q, Zhang Y C, Ma R H, et al. Spatial and temporal dynamics of floating algal blooms in lake Chaohu in 2016 and their environmental drivers[J]. Environmental Science, 2018, 39(11): 4925-4937. |
| [6] |
余茂蕾, 洪国喜, 许海, 等. 湖泊蓝藻水华对连通河道水质的影响[J]. 环境科学, 2019, 40(2): 603-613. Yu M L, Hong G X, Xu H, et al. Effects of cyanobacterial blooms in eutrophic lakes on water quality of connected rivers[J]. Environmental Science, 2019, 40(2): 603-613. |
| [7] |
郑丙辉, 张佳磊, 王丽婧, 等. 大宁河水华敏感期浮游植物与环境因子关系[J]. 环境科学, 2011, 32(3): 641-648. Zheng B H, Zhang J L, Wang L J, et al. Exploration of relationships between phytoplankton and related environmental factors in the Daning river during sensitive period of algal blooms[J]. Environmental Science, 2011, 32(3): 641-648. |
| [8] |
马沛明, 施练东, 张俊芳, 等. 浙江汤浦水库浮游植物季节演替及其影响因子分析[J]. 环境科学, 2016, 37(12): 4560-4569. Ma P M, Shi L D, Zhang J F, et al. Succession of phytoplankton assemblages and its influencing factors in Tangpu reservoir, Zhejiang province[J]. Environmental Science, 2016, 37(12): 4560-4569. |
| [9] |
王立明, 林超, 刘德文. 水动力学条件下潘家口水库富营养化的控制[J]. 水资源保护, 2009, 25(4): 8-11. Wang L M, Lin C, Liu D W. Study on eutrophication control of Panjiakou reservoir under hydrodynamic conditions[J]. Water Resources Protection, 2009, 25(4): 8-11. DOI:10.3969/j.issn.1004-6933.2009.04.003 |
| [10] | 谢飞. 珠海供水水库富营养化现状与浮游植物群落特征[D]. 广州: 暨南大学, 2014. |
| [11] | 邹红菊. 珠海市三座调水水库浮游植物群落动态与藻类水华预警研究[D]. 广州: 暨南大学, 2010. |
| [12] |
谢琼, 付青, 昌盛, 等. 珠海市江库连通条件下饮用水源水质分布特征及水资源调配措施[J]. 环境工程技术学报, 2022, 12(4): 1075-1085. Xie Q, Fu Q, Chang S, et al. Water quality distribution characteristics and water resources allocation measures of river-reservoir connected drinking water sources in Zhuhai city[J]. Journal of Environmental Engineering Technology, 2022, 12(4): 1075-1085. |
| [13] |
涂新军, 吴海鸥, 陈晓宏, 等. 滨海地区基于避咸蓄淡模式的供水安全临界流量研究——以粤港澳大湾区珠澳供水为例[J]. 湖泊科学, 2022, 34(1): 194-206. Tu X J, Wu H O, Chen X H, et al. Critical streamflow for water supply safety based on the withdrawal avoiding saltwater model in coastal areas——taking the water supply for Zhuhai and Macao in the Guangdong-Hong Kong-Macao Greater Bay Area as a case[J]. Journal of Lake Sciences, 2022, 34(1): 194-206. |
| [14] |
刘智森. 骨干水库调度破解珠江五大水问题[J]. 人民珠江, 2007, 28(1): 44-47. Liu Z S. United deployment of the key reservoirs solved the five major water problems in the Pearl River Basin[J]. Pearl River, 2007, 28(1): 44-47. DOI:10.3969/j.issn.1001-9235.2007.01.009 |
| [15] |
刘斌, 刘丽诗, 吴炜, 等. 基于取淡与流量控制的压咸调度方案研究[J]. 水文, 2013, 33(4): 84-86, 74. Liu B, Liu L S, Wu W, et al. Pearl river water diversion research based on drinking water supply target and flow control[J]. Journal of China Hydrology, 2013, 33(4): 84-86, 74. |
| [16] |
胡培, 邱静. 水动力影响下的调水水库营养物质变化规律研究——以珠海市大镜山水库为例[J]. 中国农村水利水电, 2015(9): 58-61. Hu P, Qiu J. Research on the nutrients change of pumped storage reservoir under hydrodynamics: a case study of Dajingshan reservoir in Zhuhai city[J]. China Rural Water and Hydropower, 2015(9): 58-61. DOI:10.3969/j.issn.1007-2284.2015.09.014 |
| [17] | 国家环境保护总局. 水和废水监测分析方法[M]. 北京: 中国环境科学出版社, 1990. |
| [18] | 胡鸿钧, 魏印心. 中国淡水藻类: 系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [19] | Liu X, Lu X H, Chen Y W, et al. The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: an 11-year investigation[J]. Harmful Algae, 2011, 10(3): 337-343. DOI:10.1016/j.hal.2010.12.002 |
| [20] | Lv H, Yang J, Liu L M, et al. Temperature and nutrients are significant drivers of seasonal shift in phytoplankton community from a drinking water reservoir, subtropical China[J]. Environmental Science and Pollution Research, 2014, 21(9): 5917-5928. DOI:10.1007/s11356-014-2534-3 |
| [21] |
孟睿, 何连生, 过龙根, 等. 长江中下游草型湖泊浮游植物群落及其与环境因子的典范对应分析[J]. 环境科学, 2013, 34(7): 2588-2596. Meng R, He L S, Guo L G, et al. Canonical correspondence analysis between phytoplankton community and environmental factors in macrophtic lakes of the middle and lower reaches of Yangtze River[J]. Environmental Science, 2013, 34(7): 2588-2596. |
| [22] | Reynolds C S. What factors influence the species composition of phytoplankton in lakes of different trophic status?[J]. Hydrobiologia, 1998, 369: 11-26. |
| [23] | Straskraba M. Retention time as a key variable of reservoir limnology[A]. In: Tundisi J G, Straskraba M (Eds. ). Theoretical Reservoir Ecology and its Applications[M]. Sao Carlos: International Institute of Ecology, Brazilian Academy of Sciences, Backhuys Publishers, 1999. |
| [24] | Kawara O, Yura E, Fujii S, et al. A study on the role of hydraulic retention time in eutrophication of the Asahi River Dam reservoir[J]. Water Science & Technology, 1998, 37(2): 245-252. |
| [25] |
刘格辛, 吴孟李, 尹娟, 等. 西江调水对珠海市供水水库富营养化的影响及其管理对策[J]. 生态科学, 2013, 32(4): 494-499. Liu G X, Wu M L, Yin J, et al. Influence of pumping water from Xijiang river on eutrophication of water supply reservoirs and its management strategy in Zhuhai city[J]. Ecological Science, 2013, 32(4): 494-499. |
| [26] | Monsen N E, Cloern J E, Lucas L V, et al. A comment on the use of flushing time, residence time, and age as transport time scales[J]. Limnology and Oceanography, 2002, 47(5): 1545-1553. DOI:10.4319/lo.2002.47.5.1545 |
| [27] |
王振方, 张玮, 杨丽, 等. 异龙湖不同湖区浮游植物群落特征及其与环境因子的关系[J]. 环境科学, 2019, 40(5): 2249-2257. Wang Z F, Zhang W, Yang L, et al. Characteristics of phytoplankton community and its relationship with environmental factors in different regions of Yilong Lake, Yunnan Province, China[J]. Environmental Science, 2019, 40(5): 2249-2257. |
| [28] |
宗梅, 郑西强, 王翔宇. 巢湖万年埠沿岸浮游植物与环境因子关系[J]. 安徽农业科学, 2016, 44(27): 102-105. Zong M, Zheng X Q, Wang X Y. The relationship between phytoplankton and related environmental factors in the littoral zone of wannianbu, Chaohu Lake[J]. Journal of Anhui Agricultural Sciences, 2016, 44(27): 102-105. DOI:10.3969/j.issn.0517-6611.2016.27.035 |
| [29] |
王丽卿, 许莉, 卢子园, 等. 淀山湖浮游植物数量消长及其与环境因子的关系[J]. 环境科学, 2011, 32(10): 2868-2874. Wang L Q, Xu L, Lu Z Y, et al. Dynamic of phytoplankton abundance and the relationship with environmental factors in Dianshan Lake, Shanghai[J]. Environmental Science, 2011, 32(10): 2868-2874. |
| [30] |
黄洪波. 调水水库的富营养化特征研究[J]. 广东化工, 2011, 38(2): 99-100, 121. Huang H B. Study of eutrophication of two pumped storage reservoirs[J]. Guangdong Chemical Industry, 2011, 38(2): 99-100, 121. DOI:10.3969/j.issn.1007-1865.2011.02.047 |
| [31] | Reynolds C S, Huszar V, Kruk C, et al. Towards a functional classification of the freshwater phytoplankton[J]. Journal of Plankton Research, 2002, 24(5): 417-428. DOI:10.1093/plankt/24.5.417 |
| [32] | Hodgson D A. The ecology of cyanobacteria: their diversity in time and space, Edited by B. A. Whitton and M. Potts[J]. Journal of Paleolimnology, 2002, 28(3): 383-384. DOI:10.1023/A:1021694308518 |
| [33] |
郭川, 代晓炫, 饶伟民, 等. 珠海市竹银水库健康评价研究[J]. 广东水利水电, 2021(3): 12-17. Guo C, Dai X X, Rao W M, et al. Study on the health assessment of the Zhuyin reservoir in Zhuhai[J]. Guangdong Water Resources and Hydropower, 2021(3): 12-17. |
| [34] | Schindler D W. Evolution of phosphorus limitation in lakes: Natural mechanisms compensate for deficiencies of nitrogen and carbon in eutrophied lakes[J]. Science, 1997, 195(4275): 260-262. |
| [35] | Hallegraeff G M. A review of harmful algal blooms and their apparent global increase[J]. Phycologia, 1993, 32(2): 79-99. DOI:10.2216/i0031-8884-32-2-79.1 |
| [36] | Temponeras M, Kristiansen J, Moustaka-Gouni M. Seasonal variation in phytoplankton composition and physical-chemical features of the shallow Lake Doirani, Macedonia, Greece[J]. Hydrobiologia, 2000, 424(1-3): 109-122. |
| [37] | Nalewajko C, Murphy T P. Effects of temperature, and availability of nitrogen and phosphorus on the abundance of Anabaena and Microcystis in Lake Biwa, Japan: an experimental approach[J]. Limnology, 2001, 2(1): 45-48. DOI:10.1007/s102010170015 |
| [38] | Steinberg C E W, Hartmann H M. Planktonic bloom-forming cyanobacteria and the eutrophication of lakes and rivers[J]. Freshwater Biology, 1988, 20(2): 279-287. DOI:10.1111/j.1365-2427.1988.tb00452.x |
| [39] |
邹红菊, 胡韧, 韩博平. 鹤地水库浮游植物群落的结构与动态[J]. 热带亚热带植物学报, 2010, 18(2): 196-202. Zou H J, Hu R, Han B P. Structure and dynamics of phytoplankton community in Hedi reservoir, south China[J]. Journal of Tropical and Subtropical Botany, 2010, 18(2): 196-202. DOI:10.3969/j.issn.1005-3395.2010.02.014 |
| [40] | Berge T, Daugbjerg N, Andersen B B, et al. Effect of lowered pH on marine phytoplankton growth rates[J]. Marine Ecology Progress, 2010, 416: 79-91. DOI:10.3354/meps08780 |
| [41] | Jiang Y J, He W, Liu W X, et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu)[J]. Ecological Indicators, 2014, 40: 58-67. DOI:10.1016/j.ecolind.2014.01.006 |
| [42] | Kasprzak P, Lathrop R C, Carpenter S R. Influence of different sized Daphnia species on chlorophyll concentration and summer phytoplankton community structure in eutrophic Wisconsin lakes[J]. Journal of Plankton Research, 1999, 21(11): 2161-2174. DOI:10.1093/plankt/21.11.2161 |
| [43] | Rao K, Zhang X, Wang M, et al. The relative importance of environmental factors in predicting phytoplankton shifting and cyanobacteria abundance in regulated shallow lakes[J]. Environmental Pollution, 2021, 286. DOI:10.1016/j.envpol.2021.117555 |
| [44] | Zhao C S, Pan X, Yang S T, et al. Principal hydrology and water quality factors driving the development of plankton communities in a pilot city of China[J]. Ecohydrology, 2020, 13(4). DOI:10.1002/eco.2207 |
| [45] | 白英臣. 天然溶解有机质和环境污染物的相互作用——以铜、汞离子和卡马西平为例[D]. 贵阳: 中国科学院地球化学研究所, 2007. |
| [46] | Moisander P H, Cheshire L A, Braddy J, et al. Facultative diazotrophy increases cylindrospermopsis raciborskii competitiveness under fluctuating nitrogen availability[J]. FEMS Microbiology Ecology, 2012, 79(3): 800-811. DOI:10.1111/j.1574-6941.2011.01264.x |
| [47] | Ammar M, Comte K, Tran T D C, et al. Initial growth phases of two bloom-forming cyanobacteria (Cylindrospermopsis raciborskii and Planktothrix agardhii) in monocultures and mixed cultures depending on light and nutrient conditions[J]. Annales de Limnologie-International Journal of Limnology, 2014, 50(3): 231-240. DOI:10.1051/limn/2014096 |
| [48] | Tian C, Pei H Y, Hu W R, et al. Variation of cyanobacteria with different environmental conditions in Nansi Lake, China[J]. Journal of Environmental Sciences, 2012, 24(8): 1394-1402. DOI:10.1016/S1001-0742(11)60964-9 |
| [49] | Gao J S, Zhu J, Wang M W, et al. Dominance and growth factors of Pseudanabaena sp. in drinking water source reservoirs, southern China[J]. Sustainability, 2018, 10(11). DOI:10.3390/su10113936 |
| [50] | Zhang H H, Zong R R, He H Y, et al. Biogeographic distribution patterns of algal community in different urban lakes in China: insights into the dynamics and co-existence[J]. Journal of Environmental Sciences, 2021, 100: 216-227. DOI:10.1016/j.jes.2020.07.024 |
| [51] | 陈悦. 深圳福田红树林区浮游植物生态特征及主要环境因子对孟氏小环藻胞外多糖的影响[D]. 厦门: 厦门大学, 2017. |