 2022, Vol. 43
2022, Vol. 43



2. 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125;
3. 中国科学院大学, 北京 100049
2. Key Laboratory of Subtropical Agriculture Ecology, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
稻田在我国粮食安全中占有举足轻重的地位, 总面积达3 400万hm2, 主要分布在亚热带地区, 该区稻田占全国总面积的31.4%[1, 2].稻田土壤具有明显的碳汇功能, 开展稻田的固碳和温室气体排放研究, 对减缓温室气体排放和实现“双碳”计划具有重要意义[3, 4].
长期秸秆还田可以有效地提高土壤的养分和有机碳含量, 改变土壤物理、化学和生物环境, 从而影响水稻土有机碳的周转和积累[5, 6], 因此在我国得到大力推广, 且卓有成效. 2000~2010年间我国水稻的秸秆还田比例由不足25%增长到近40%[7]. Wang等[8]的研究对长期(>10 a)秸秆还田试验的荟萃分析显示, 水稻秸秆还田的碳固定效率为9.4%, 长期水稻秸秆还田可将土壤有机碳(SOC)含量提高约12%. Wang等[9]的荟萃分析也显示长期水稻秸秆还田可将土壤有机碳含量提高5.4%.秸秆还田不仅对土壤有机碳库有所贡献, 还会起到改良土壤物理结构, 改善土壤通透性和蓄水能力的作用, 其分解还会释放氮磷等养分元素[5, 6].然而, 目前秸秆还田的相关研究多集中在其对土壤有机碳库库容、养分含量以及其他物理、化学和生物条件等造成的影响, 以及秸秆在土壤中的分解、矿化和转化过程, 缺乏对长期秸秆还田情况下其他外源有机物进入土壤后的矿化和转化过程的研究.
水稻根系是稻田土壤有机碳的重要来源之一[10, 11], 水稻根系在土壤中的分解相当缓慢, 与其他新鲜碳源如秸秆和叶片等相比, 对土壤有机碳的贡献可能更大[12, 13].有研究显示, 约8.7%~20%水稻根系碳会转化为土壤有机碳[13, 14].水稻根系进入土壤中不仅可以补充土壤有机碳库, 还会增加土壤本身有机碳的矿化, 即产生正激发效应[14, 15], 且小于水稻地上部引起的激发效应[16].有机物在土壤中的分解和矿化受土壤物理、化学和生物等环境因素的影响, 如土壤肥力、水分条件和土壤质地等[15, 17, 18].然而, 目前关于外源有机物在土壤中的矿化研究多集中于秸秆和有机肥等有机物料或者单一小分子有机物, 针对水稻根系残留的研究极其缺乏[10], 尤其长期秸秆还田改造下的土壤中根系矿化过程的研究很少, 鉴于水稻根系碳在土壤有机碳中的重要贡献, 其分解和矿化过程及其影响因素等方面亟待深入研究.
因此, 本研究依托于湖南省宁乡县的长期定位试验, 通过室内培养试验和13C稳定同位素标记示踪技术, 探讨和量化了长期秸秆还田对水稻根系矿化特征及其激发效应方向和强度的影响, 以期为更好地了解稻田生态系统碳循环以及为稻田固碳减排提供理论指导.
1 材料与方法 1.1 供试土壤供试土壤采自湖南省宁乡县长期定位试验站(112°18′E, 28°07′N), 种植制度为早稻-晚稻-冬闲模式, 试验始于1986年, 该地区属于典型的亚热带季风湿润气候, 平均海拔36.1 m, 年均气温为16.8℃, 每年蒸发量均为1 353.9 mm, 每年无霜期274 d左右, 年均降雨量是1 553.7 mm.采样点土壤氮、磷和钾供应状况根据作物预期需求量.本试验选取田间3种长期不同施肥处理的土壤: ①无施肥处理(CK); ②化肥处理(CF), 早稻施肥量为142.5 kg·hm-2(以N计), 54.0 kg·hm-2(以P2O5计), 63.0 kg·hm-2(以K2O计), 晚稻施肥量为157.5 kg·hm-2(以N计), 43.2 kg·hm-2(以P2O5计), 81.0 kg·hm-2(以K2O计); ③化肥+秸秆还田处理(CFS), 早稻秸秆的还田量为2 775 kg·hm-2, 晚稻的还田量为3 600 kg·hm-2, 以CF处理的施氮量为标准, 不足用化肥补足.采集耕作层(0~20 cm)土壤样品, 人工挑出石块和肉眼可见动植物残体和根系等杂物, 过10目筛, 充分混匀.将混匀的土壤样品按照四分法取出一部分, 经风干过筛后测定土壤样品的基本理化性质(表 1).
|
|
表 1 供试土壤基本理化性质1) Table 1 Physiochemical properties of the soil |
1.2 13C标记根系制备
在PVC盆中(直径17.2 cm, 高度16.7 cm)移栽25日龄的水稻幼苗, 每盆3株, 置于密闭植物生长箱(长×宽×高: 150 cm×100 cm×150 cm), 连续不断地输入CO2维持含量400 μL·L-1.在水稻营养生长期, 包括水稻分蘖期和伸长期, 对水稻连续输入13CO2(13C丰度2.8%)进行标记, 并在孕穗期开始时停止标记.将生长箱置于稻田中, 使标记植物和未标记对照物的环境条件相同.于成熟期进行破坏性采样, 将水稻根系与土壤分离.根系用水反复冲洗去除附着的土壤.根系于105℃杀青0.5 h后, 置于65℃的烘箱中烘干并粉碎保存.最终制得根系的13C丰度为2.152%, 含碳量为37.75%.
1.3 土壤培养试验对不同施肥土壤分别设置添加13C标记根系(CK+R、CF+R和CFS+R)与不加根(CK、CF和CFS)处理, 共6个处理, 3个重复, 另外设置添加未标记根系的自然丰度对照组.将风干土淹水至水层深约2~3 cm, 置于25℃的恒温培养暗室进行10 d预培养.预培养结束后, 将上清液倒掉, 将土样与13C标记根系按2.5g·kg-1(干土计)的碳添加量均匀混合, 称20 g(干土计)加入到500 mL的培养瓶中, 淹水至水层约为2~3 cm, 盖子松开保持通气状态, 置于25℃的恒温培养室中遮光培养120 d.
1.4 样品采集与分析每隔1~5 d进行气体采集, 用一次性针管注射器抽取培养瓶顶空气体35 mL, 注入已抽真空的12 mL贮藏玻璃瓶待测.每次采气结束后, 通过气泵将瓶中气体置换为新鲜空气, 密封后放回25℃恒温室继续培养.气体CO2含量用气相色谱法-温室气体气相色谱仪(7890A, 美国)测定; 13CO2用稳定性同位素质谱仪(MAT 253, 美国)测定.
1.5 数据处理根系和土壤本身有机碳(SOC)的CO2释放量计算公式:

|
(1) |
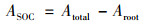
|
(2) |
式中, Atotal为各处理的CO2-C总释放量(mg·kg-1); δ13Ctotal为总释放量中13C丰度; ASOC为来源于SOC的CO2释放量; δ13CSOC为SOC的13C丰度(自然丰度对照); δ13Croot为根系的13C丰度; Aroot为根系的CO2释放量.根系的矿化率是根系CO2累积释放量与根系添加碳量的比值(质量分数).
激发效应和相对激发效应计算公式:

|
(3) |

|
(4) |
式中, ANonroot为不加根处理的CO2累积释放量.
所有数据计算均为3次重复的平均值, 运用Microsoft Excel 2010进行整理, 用Origin 2018绘图分析.用SPSS 21.0进行不同处理间的单因素方差分析(One-way ANOVA), 显著性水平为P < 0.05, 并用Tukey法进行多重比较.
2 结果与分析 2.1 水稻土CO2累积释放特征在0~120 d培养期间, 不同施肥处理土壤前期CO2累积释放量快速增长, 之后增速逐渐减缓(图 1).添加根系土壤CO2的累积释放量显著高于不添加根系的土壤, 120 d时相较于CK、CF和CFS处理, 加根处理累积矿化量分别增加了726.3、617.4和724.5 mg·kg-1.不添加根系的土壤中, 培养前期(0~30 d)CFS处理的CO2累积释放量显著高于CK和CF处理, 之后CFS和CF处理间CO2累积释放量的差距逐渐减小至无显著差异, 在120 d时CF和CFS处理的CO2累积释放量分别比CK处理增加了19.6%和18.2%(P < 0.05).添加根系的土壤中, 培养期间CFS+R和CF+R处理的CO2累积释放量高于CK+R处理, 在培养结束时, CFS+R、CF+R和CK+R处理的CO2累积释放量分别为1 800.3、1 706.8和1 636.9 mg·kg-1.

|
CK: 不施肥, CF: 化肥, CFS: 秸秆还田配施化肥, CK+R: 不施肥+根系, CF+R: 化肥+根系, CFS+R: 秸秆还田配施化肥+根系; 不同小写字母表示120 d各不加根处理间或各加根处理间差异的显著性(P < 0.05), 下同 图 1 不同长期施肥处理水稻土添加和不添加根系条件下CO2累积释放量 Fig. 1 Cumulative CO2 emission with and without root addition under different fertilization |
相较于不施肥处理, 单施化肥和添加秸秆处理中根系矿化生成CO2的比例明显增加(图 2).培养前40 d, CFS+R处理的根系CO2累积释放量显著高于CK+R和CF+R处理, 之后3个处理的根系CO2释放速率同时加快, 且CF+R处理的CO2释放速率最快, 其根系CO2累积释放量与CFS+R处理的差距逐渐缩小, 并在76 d以后逐渐超过CFS+R处理的CO2累积释放量, 但是差异不显著.在90~120 d时各处理根系CO2释放速率变缓.在培养120 d结束时, CF+R、CFS+R和CK+R处理中根系CO2累积释放量分别为470.8、444.0和384.2 mg·kg-1, CF+R处理和CFS+R处理根系CO2累积释放量显著高于CK+R, 三者根系的矿化率分别为18.8%、17.8%和15.4%.

|
图 2 不同长期施肥水稻土根系来源的CO2累积释放量及其矿化率 Fig. 2 Cumulative CO2 emission from root and mineralized proportion of root under different long-term fertilization |
SOC矿化生成的CO2累积释放量在前期0~20 d内快速升高, 后期释放速率逐渐变缓.CFS+R处理的SOC来源CO2累积释放量略高于CK+R和CF+R处理, 但差异不显著[图 3(a)].但因CFS+R土壤的SOC含量较高, 其SOC的累积矿化率显著低于CK+R和CF+R处理. 120 d时, CK+R、CF+R和CFS+R处理的SOC累积矿化率分别为5.8%、5.4%和4.2%[图 3(b)].

|
图 3 不同长期施肥水稻土本身有机碳(SOC)来源的CO2累积释放量及其矿化率 Fig. 3 Cumulative CO2 emission and mineralized proportion of soil organic carbon (SOC) under different long-term fertilization |
水稻根系导致土壤CO2累积释放的正激发效应.培养前期(0~40 d), 3个处理的正CO2累积激发效应迅速增长, 且CFS+R处理最快, 40 d时CK+R、CF+R和CFS+R处理的CO2累积激发效应分别为234.2、184.9和256.8 mg·kg-1.随后CK+R和CFS+R处理的CO2累积激发效应增速减缓, CF+R处理的正CO2累积激发效应逐渐降低.在120 d培养结束后, CK+R、CF+R和CFS+R处理的CO2累积激发效应分别为386.8、156.2和316.5 mg·kg-1[图 4(a)].水稻根系导致的CO2相对累积激发效应[图 4(a)]与CO2累积激发效应[图 4(b)]不同, 3个处理的CO2相对累积激发效应0~40 d迅速增长, 之后均逐渐下降, 且40~120 d呈现出: CK+R>CFS+R>CF+R, 在120 d时, 3种处理的CO2相对累积激发效应分别为42.5%、29.6%和14.4%.

|
图 4 不同长期施肥水稻土中根系的CO2累积激发效应和CO2相对累积激发效应 Fig. 4 Cumulative and relative cumulative priming effects of roots in paddy soils with different long-term fertilization |
培养结束时, CO2累积排放量23.5%~27.6%来源于根系, 72.4%~76.5%来源于SOC, 其中, 因根系引起的正CO2累积激发效应贡献了CO2累积排放量的8.6%~20.9%(图 5).CF+R处理的根系来源CO2累积排放量占比最高, CFS+R处理次之.CF+R处理的SOC来源CO2累积排放量占比为72.4%, 其中8.6%源于根系引起的正CO2累积激发效应.CFS+R处理的正CO2累积激发效应占比是CF+R处理的近两倍(15.6%), 且低于CK+R处理(20.9%), 见图 5.

|
图 5 120 d添加根系水稻土CO2排放来源与比例 Fig. 5 Partitioning of C sources for CO2 emissions in paddy soil with added roots at 120 d |
农业土壤中土壤有机碳的积累对维持土壤肥力和作物生产力至关重要, 水稻根系是稻田土壤有机碳碳库中重要的输入碳源[11]. Liu等[11]总结了连续标记和脉冲标记试验结果, 发现水稻光合碳约10%~13%分配在根系之中.其中8.7%[13]或20%[14]水稻根系碳会转化为土壤有机碳, 取决于土壤水分条件.根系中木质素和软木脂等难分解有机碳组分的含量较高, 因此根系的矿化率普遍低于秸秆或叶片等作物其他部位的残留物[10, 12, 19, 20].本文经过120 d淹水培养, 15.4%~18.8%的根系碳矿化为CO2(图 2).类似地, Kramer等[21]经过32 d培养, 玉米根系矿化率约为12%, 小于玉米叶片矿化率(45%).Viswanath等[22]在保持田间持水量的情况下, 室内培养150 d后, 水稻秸秆和小麦秸秆的矿化率分别为81%和73%, 均高于本文和Kramer等[21]的根系矿化率, 虽然矿化率一定程度上与培养时间和土壤水分状况有关, 但仍能显示出根系矿化率普遍低于秸秆矿化率. Zhu等[16]的研究中, 虽然水稻根系矿化率为45.4%, 高于秸秆31.9%, 但是其根系碳的平均滞留时间大于秸秆碳.刘晖等[23]的室外培养试验显示, 在培养5个月时, 水稻根系累积矿化率低于秸秆碳, 培养1 a时, 水稻根系和秸秆碳累积矿化率相近.根系产生CO2在总释放量中的占比也小于水稻秸秆的CO2占比.本文根系产生CO2累积释放量不足总释放量的30%(图 5), 这与Guo等[10]的研究结果一致. Guo等[10]的研究中培养前14 d, 11%~19%的CO2来源于根系, 28 d以后不足5%.类似地, 玉米根系产生CO2占比为12%~17%[21].而水稻秸秆CO2释放量占总释放量可达30%~80%[24].
水稻根系在土壤中会引起正激发效应.因为根系碳相对土壤有机碳更易被微生物利用, 当外源有机碳的可利用性较高时, 会刺激微生物群落为了达到碳氮平衡而对土壤本身有机碳进行矿化分解, 从而挖掘更多矿质氮, 因而产生正激发效应.本文根系CO2相对累积激发效应为14.4%~42.5%, 且培养前40 d CO2相对累积激发效应更高(图 4). Zhu等[16]的研究结果与本文相似, 水稻根系的激发效应在培养50 d后达到最大, 为43%, 之后下降至2.9%.然而, 秸秆的激发效应大于根系, 可达52%~378%[16].这与根系和秸秆的易利用和难利用等多种组分形态有关[21].
3.2 秸秆还田对水稻根系矿化和激发效应的影响秸秆还田是提高土壤碳库和土壤肥力的重要措施, 一直以来得到大力推广.不同的荟萃分析研究显示长期水稻秸秆还田可将土壤有机碳含量提高5.4%~12%[8, 9].本文土壤经过约30 a的秸秆还田, 土壤有机碳含量与CK相比提高了48%(表 1).有机质含量对土壤养分含量、微生物群落结构和活性和生物酶活性等具有显著影响[14, 25].本文秸秆还田土壤较CK全氮、碱解氮和全磷含量分别增长了53%、55%和177%.秸秆还田还会改良土壤物理结构, 改善土壤通透性, 本文秸秆还田土壤较CK的黏粒和粉粒含量分别降低了18%和14%(表 1).因此, 秸秆还田利于土壤微生物生长和对有机物的分解矿化.宁赵等[26]的研究显示秸秆还田会增加土壤微生物生物量和生长速率, 提高微生物对羧酸类、氨基酸和碳水化合物的利用速率.因此, 本文不加根的秸秆还田土壤CO2累积释放量比CK多18.1%(图 1), 加根的秸秆还田处理的根系产生CO2累积释放量比CK增加了22.5%(图 2).另外, 水稻土中有机质的转化过程以真菌为主[27, 28], 长期秸秆还田土壤中真菌含量显著高于无秸秆还田土壤[27].真菌相比于细菌更擅长和偏好分解难降解有机物[27, 28], 因此秸秆还田处理相较于CK处理对根系的分解能力更强, 根系产生CO2和矿化率更高(图 2).
与单施化肥相比, 本文秸秆还田处理在培养前期根系CO2释放量更高, 之后因单施化肥处理的根系CO2释放速率更快, 培养结束时二者根系CO2累积释放量无显著差异(图 2).这可能是由于培养初期, 如前面讨论所述, 因秸秆还田土壤微生物生物量和活性更高, 根系矿化更快, 但由于矿化过程中对土壤养分如氮磷的消耗, 导致养分计量比逐渐不平衡, 限制了矿化速率, 而化肥土壤相对秸秆还田土壤的氮磷养分占比更大[5], 因此根系的矿化能够持续高速进行, 最终导致两个处理间的累积矿化率相近.宁赵等[26]的研究也显示化肥和秸秆还田处理的外源碳代谢特征相似.
土壤本身有机碳的矿化率为4.2%~5.8%(图 3), 小于根系矿化率, 表明土壤中难分解有机碳组分更多.秸秆还田土壤本身有机碳的CO2累积释放量略高于CK和化肥处理, 但差异不显著.但因秸秆还田土壤有机碳含量较高, 其累积矿化率显著低于CK和化肥处理(图 3), 说明秸秆还田土壤有机碳的易矿化程度更低.秸秆还田处理的根系CO2相对累积激发效应小于CK处理(图 4), 这可能是由于秸秆还田土壤养分含量相对更高, 根系矿化过程中微生物通过分解土壤本身有机碳来获取速效氮磷养分的需求强度不如CK大.化肥处理的CO2累积激发效应小于秸秆还田处理(图 4), 也是出于养分元素计量比的原因, 化肥处理的土壤中氮磷养分的相对比例更高, 因此分解土壤本身有机碳的强度不高.另外, 因真菌分解难降解有机物的能力较强且秸秆还田土壤真菌含量更高[27, 28], 即在外源有机碳刺激下对土壤本身有机碳的挖掘能力优于化肥土壤, 这可能也是导致更大激发效应的原因之一.
4 结论(1) 经过120 d的室内培养, 秸秆还田和化肥处理根系的矿化率分别为18.8%和17.8%, 显著高于CK处理的15.4%.
(2) 3个处理的土壤本身有机碳产生的CO2累积排放量无显著差异, 但是对于土壤有机碳矿化率, 秸秆还田处理显著低于化肥和CK处理.秸秆还田土壤中根系引发的CO2相对累积激发效应为29.6%, 显著低于CK处理的42.5%, 但高于化肥处理的14.4%.
(3) 淹水水稻土CO2释放量中23.47%~27.59%来源于根系, 其余来源于土壤, 其中, 激发效应引起的CO2释放量占比在CK处理中最大, 秸秆还田处理次之, 化肥处理最小.
| [1] | Zhou P, Sheng H, Li Y, et al. Lower C sequestration and N use efficiency by straw incorporation than manure amendment on paddy soils[J]. Agriculture, Ecosystems & Environment, 2016, 219: 93-100. |
| [2] | Liu S L, Huang D Y, Chen A L, et al. Differential responses of crop yields and soil organic carbon stock to fertilization and rice straw incorporation in three cropping systems in the subtropics[J]. Agriculture, Ecosystems & Environment, 2014, 184: 51-58. |
| [3] | Mackay A. Climate change 2007: impacts, adaptation and vulnerability. contribution of working group Ⅱ to the fourth assessment report of the intergovernmental panel on climate change[J]. Journal of Environmental Quality, 2008, 37(6): 2407-2407. DOI:10.2134/jeq2008.0015br |
| [4] | Pan G X, Li L Q, Wu L S, et al. Storage and sequestration potential of topsoil organic carbon in China's paddy soils[J]. Global Change Biology, 2004, 10(1): 79-92. DOI:10.1111/j.1365-2486.2003.00717.x |
| [5] | Young M D, Ros G H, De Vries W. Impacts of agronomic measures on crop, soil, and environmental indicators: a review and synthesis of meta-analysis[J]. Agriculture, Ecosystems & Environment, 2021, 319. |
| [6] | Huang W, Wu J F, Pan X H, et al. Effects of long-term straw return on soil organic carbon fractions and enzyme activities in a double-cropped rice paddy in South China[J]. Journal of Integrative Agriculture, 2021, 20(1): 236-247. DOI:10.1016/S2095-3119(20)63347-0 |
| [7] |
赵永存, 徐胜祥, 王美艳, 等. 中国农田土壤固碳潜力与速率: 认识、挑战与研究建议[J]. 中国科学院院刊, 2018, 33(2): 191-197. Zhao Y C, Xu S X, Wang M Y, et al. Carbon sequestration potential in chinese cropland soils: review, challenge, and research suggestion[J]. Bulletin of Chinese Academy of Sciences, 2018, 33(2): 191-197. |
| [8] | Wang J Z, Wang X J, Xu M G, et al. Crop yield and soil organic matter after long-term straw return to soil in China[J]. Nutrient Cycling in Agroecosystems, 2015, 102(3): 371-381. DOI:10.1007/s10705-015-9710-9 |
| [9] | Wang Y L, Wu P N, Mei F J, et al. Does continuous straw returning keep China farmland soil organic carbon continued increase? a meta-analysis[J]. Journal of Environmental Management, 2021, 288. |
| [10] | Guo T F, Zhang Q, Ai C, et al. Microbial utilization of rice root residue-derived carbon explored by DNA stable-isotope probing[J]. European Journal of Soil Science, 2021, 72(1): 460-473. DOI:10.1111/ejss.12970 |
| [11] | Liu Y L, Ge T D, Zhu Z K, et al. Carbon input and allocation by rice into paddy soils: a review[J]. Soil Biology and Biochemistry, 2019, 133: 97-107. DOI:10.1016/j.soilbio.2019.02.019 |
| [12] | Abiven S, Recous S, Reyes V, et al. Mineralisation of C and N from root, stem and leaf residues in soil and role of their biochemical quality[J]. Biology and Fertility of Soils, 2005, 42(2): 119-128. DOI:10.1007/s00374-005-0006-0 |
| [13] | Lu Y H, Watanabe A, Kimura M. Carbon dynamics of rhizodeposits, root- and shoot-residues in a rice soil[J]. Soil Biology and Biochemistry, 2003, 35(9): 1223-1230. DOI:10.1016/S0038-0717(03)00184-6 |
| [14] | Zhu Z K, Ge T D, Hu Y J, et al. Fate of rice shoot and root residues, rhizodeposits, and microbial assimilated carbon in paddy soil - part 2: turnover and microbial utilization[J]. Plant and Soil, 2017, 416: 243-257. DOI:10.1007/s11104-017-3210-4 |
| [15] | Kuzyakov Y. Priming effects: interactions between living and dead organic matter[J]. Soil Biology and Biochemistry, 2010, 42(9): 1363-1371. DOI:10.1016/j.soilbio.2010.04.003 |
| [16] | Zhu Z K, Zeng G J, Ge T D, et al. Fate of rice shoot and root residues, rhizodeposits, and microbe-assimilated carbon in paddy soil - Part 1: decomposition and priming effect[J]. Biogeosciences, 2016, 13(15): 4481-4489. DOI:10.5194/bg-13-4481-2016 |
| [17] | Saidy A R, Smernik J A, Baldock K, et al. Effects of clay mineralogy and hydrous iron oxides on labile organic carbon stabilisation[J]. Geoderma, 2012, 173-174: 104-110. DOI:10.1016/j.geoderma.2011.12.030 |
| [18] | Sodano M, Said-Pullicino D, Fiori A F, et al. Sorption of paddy soil-derived dissolved organic matter on hydrous iron oxide-vermiculite mineral phases[J]. Geoderma, 2016, 261: 169-177. DOI:10.1016/j.geoderma.2015.07.014 |
| [19] | Amin B A Z, Chabbert B, Moorhead D, et al. Impact of fine litter chemistry on lignocellulolytic enzyme efficiency during decomposition of maize leaf and root in soil[J]. Biogeochemistry, 2014, 117: 169-183. DOI:10.1007/s10533-013-9856-y |
| [20] | Insalud N, Bell R W, Colmer T D, et al. Morphological and physiological responses of rice (Oryza sativa) to limited phosphorus supply in aerated and stagnant solution culture[J]. Annals of Botany, 2006, 98(5): 995-1004. DOI:10.1093/aob/mcl194 |
| [21] | Kramer S, Dibbern D, Moll J, et al. Resource partitioning between bacteria, fungi, and protists in the detritusphere of an agricultural soil[J]. Frontiers in Microbiology, 2016, 7. |
| [22] | Viswanath T, Pal D, Purakayastha T J. Elevated CO2 reduces rate of decomposition of rice and wheat residues in soil[J]. Agriculture, Ecosystems & Environment, 2010, 139(4): 557-564. |
| [23] |
刘晖, 刘杰, 张玉玲, 等. 外源水稻根系和茎叶碳氮在稻田土壤中释放的特征[J]. 土壤学报, 2021, 58(4): 989-997. Liu H, Liu J, Zhang Y L, et al. Release of exogenous carbon and nitrogen in rice root, stem and leaf in paddy soil[J]. Acta Pedologica Sinica, 2021, 58(4): 989-997. |
| [24] | Pan F X, Li Y Y, Chapman S J, et al. Microbial utilization of rice straw and its derived biochar in a paddy soil[J]. Science of the Total Environment, 2016, 559: 15-23. DOI:10.1016/j.scitotenv.2016.03.122 |
| [25] |
战厚强, 颜双双, 王家睿, 等. 水稻秸秆还田对土壤磷酸酶活性及速效磷含量的影响[J]. 作物杂志, 2015(2): 78-83. Zhan H Q, Yan S S, Wang J R, et al. Effects of rice straw returning on activities of soil phosphatase and available P values in soil[J]. Crops, 2015(2): 78-83. |
| [26] |
宁赵, 程爱武, 唐海明, 等. 长期施肥下水稻根际和非根际土壤微生物碳源利用特征[J]. 环境科学, 2019, 40(3): 1475-1482. Ning Z, Cheng A W, Tang H M, et al. Microbial carbon source metabolic profile in rice rhizosphere and non-rhizosphere soils with different long-term fertilization management[J]. Environmental Science, 2019, 40(3): 1475-1482. |
| [27] |
宁赵, 陈香碧, 唐海明, 等. 不同施肥处理下水稻根际和非根际土壤中氨基糖积累特征[J]. 应用生态学报, 2019, 30(1): 189-197. Ning Z, Cheng X B, Tang H M, et al. Characteristics of amino sugar accumulation in rhizosphere and non-rhizosphere soil of rice under different fertilization treatments[J]. Chinese Journal of Applied Ecology, 2019, 30(1): 189-197. |
| [28] | Luo Y, Xiao M L, Yuan H Z, et al. Rice rhizodeposition promotes the build-up of organic carbon in soil via fungal necromass[J]. Soil Biology and Biochemistry, 2021, 160. |