 2022, Vol. 43
2022, Vol. 43



2. 中国环境科学研究院湖泊水污染治理与生态修复技术国家工程实验室, 北京 100012
2. National Engineering Laboratory for Lake Pollution Control and Ecological Restoration, Chinese Research Academy of Environmental Sciences, Beijing 100012, China
浮游生物是在自然水体中自由浮游生活的一类水生生物, 包括浮游植物和浮游动物[1].浮游植物是水生态系统的初级生产者和食物链的基础环节, 能够利用光合作用合成有机物[2]; 浮游动物作为食物链的中间环节, 以浮游植物和水体中有机碎屑为食, 同时也作为水体中鱼类等高级生物的饵料[3].浮游生物作为湖泊水生态系统中重要的基础环节, 具有个体微小、细胞结构简单和生命周期短的特点, 因此对水体环境变化较为敏感, 能够对环境变化迅速作出响应[4], 其群落结构与环境因子的响应关系可作为水质状况评价指标[5].
不同的水文条件和水体理化因子, 造成浮游生物的物种组成、优势种、物种丰度和生物量等群落分布特征存在差异[6].同一湖泊不同湖区由于驳岸类型、地形、水深和水生植物覆盖度等因素的不同而造成生境异质性, 从而引起浮游生物群落结构的差异性, 且影响不同湖区浮游生物分布的主导因子也不同[7].沉水植物对环境因子和浮游生物群落具有一定影响: ①通过吸收水体DTN和DTP用于自身生长繁殖, 同时分泌助凝物对不可利用的氮磷进行吸附沉降从而降低水体氮磷浓度[8, 9].②为浮游动物提供栖息地和食物来源[10], 与浮游植物竞争光照和营养同时分泌化感物质, 抑制藻类生长避免水华蓝藻暴发[11, 12].
嘉兴南湖是中共一大会址, 红船精神的发源地, 是浙江省嘉兴市著名的红色旅游景点.南湖处于长江水系太湖流域[13], 周围河网密集, 受流域性水环境、旅游业和河岸绿化带施肥等因素影响, 导致南湖水体氮、磷营养盐浓度居高不下, 部分水域蓝藻频频暴发[14]. 2020年7月在南湖水生态问题严重的湖区分不同密度种植沉水植物, 工程实施后水体透明度大幅度提升, 氮磷营养盐浓度下降.但是, 有关修复湖区水生态方面研究较少, 导致对工程实施后南湖不同修复湖区水生态状况尚不清楚, 缺少工程参数.鉴于此, 本研究于2021年1月(竣工后)对南湖4个修复湖区水体理化指标和浮游生物进行了调查, 采用冗余分析(redundancy analysis, RDA)研究了不同湖区浮游植物与浮游动物群落分布的环境驱动因子, 通过聚类热图分析和主坐标分析(principal coordinate analysis, PCoA)研究不同湖区浮游生物群落差异的原因, 通过了解不同湖区生态系统特征, 选出最优工程参数, 以期为南湖水生态修复工程实施和环境保护提供数据支撑和参考.
1 材料与方法 1.1 研究区概况南湖地处太湖平原河网, 位于嘉兴市中心城区, 常年湖面面积约为0.58 km2, 是海盐塘、平湖塘和嘉善塘等多条河流干道的交汇处, 起到重要的枢纽作用, 四周地势低平河港纵横.嘉兴市地处北亚热带南缘, 属东亚季风区, 日照充足, 雨水丰沛, 气温适中, 年平均气温15.9℃.南湖景区的南部以及西部的湖区是游客经常休闲游览的区域, 水生态问题严重. 2020年7月在这些区域设置沉水植被修复示范区, 种植以苦草为主的沉水植物, 根据工程实施时沉水植物种植密度和方式的不同, 分为4个修复湖区进行研究. A区: 沉水植物覆盖30%, 全湖区均匀种植; B区: 沉水植物覆盖30%, 岸边高密度种植; C区: 沉水植物覆盖10%; D区: 沉水植物覆盖60%.
1.2 样品采集与处理 1.2.1 采样点布设本研究共布设24个采样点(图 1), 在A区与B区岸边及湖中心区域分别均匀布设7个点(A1~A7)和4个点(B1~B4); 在C区东西侧进出水口及中心区布设4个采样点(C1~C4); 在D区出入水口及沉水植物种植区布设7个点(D1~D7); 在未种植沉水植物区设置两个参照点S区(S1~S2).采样点布设原则为: 充分覆盖4个湖区水体和采样点采集的样品要对整个调查区域有较好的代表性和覆盖了地方常规检测点位.

|
图 1 南湖不同湖区采样点分布示意 Fig. 1 Distributions of sampling sites in different regions of Nanhu Lake |
水温(WT)、酸碱度(pH)和溶解氧(DO)使用多参数水质测定仪(YSI professional plus, 美国)测定.透明度(SD)使用塞氏盘法测定.采用有机玻璃采水器采集上覆水样品, 水样置于干净的样品瓶中, 于保温箱中4℃保存并尽快带回实验室.总氮(TN)、溶解性总氮(DTN)、氨氮(NH4+-N)、硝氮(NO3--N)、总磷(TP)、溶解性总磷(DTP)、无机磷(IP)和总有机碳(TOC)参照文献[15]的方法进行测定.
1.2.3 浮游动物与浮游植物浮游植物: 定性采集使用25号(孔径0.064 mm)浮游生物网在水下呈“∞”形拖拽5 min后将滤出液收集于小瓶内加入4%的甲醛溶液保存; 定量采集使用2 L有机玻璃采水器在水面下0.5 m和1 m处分层采水混合后收集1 L于聚乙烯瓶中, 带回实验室加入15 mL鲁哥试剂避光静置48 h, 虹吸上清液浓缩至30 mL用于镜检.浮游动物: 定性采集使用13号(孔径0.112 mm)浮游生物网, 处理方法同浮游植物; 定量采集使用2.5 L有机玻璃采水器在水下分层采集共10 L水过25号浮游生物网, 将滤出液收集于聚乙烯小瓶中加入4%甲醛溶液保存.浮游植物与浮游动物采集样液沉淀浓缩后分别使用0.1 mL和1 mL计数框进行镜检、计数和鉴定[16, 17].
1.3 数据处理与分析 1.3.1 多样性计算优势度和生物多样性计算采用Mcnaughton优势度(Y)、Shannon多样性指数(H′)和Pielou均匀度指数(J)对水体生物多样性进行计算分析, 计算公式如下[18, 19]:
Mcnaughton优势度(Y):
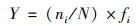
|
(1) |
Shannon-Wiener多样性指数(H'):
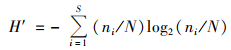
|
(2) |
Pielou均匀度指数(J):
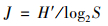
|
(3) |
式中, fi为物种i出现的频率, ni为物种i的个体数; N为全部样品的总个体数; S为样品中浮游生物物种总数.取优势度Y≥0.02的物种作为优势种. H′>3为清洁, 2~3为β-中污染, 1~2为α-中污染, < 1为重污染. J>0.8为清洁, 0.5~0.8为轻污染, 0.3~0.5为β-中污染, 0.1~0.3为α-中污染, < 0.1为重污染.
1.3.2 数据处理4个湖区环境因子、浮游生物数据相关性分析在SPSS 26.0软件中采用独立样本t检验和单因素方差分析.浮游生物群落分布与环境因子相关性分析使用R语言vegan包与ggplot2包进行排序分析并作图.除pH以外, 所有环境数据和浮游生物优势种丰度数据进行lg(x+1)标准化处理, 去趋势分析(DCA)得出浮游植物与浮游动物第一轴长分别为3.35与1.92; 长度梯度均小于4, 因此都选用RDA分析[20, 21].聚类热图和主坐标分析(PCoA)分别采用R语言heatmap(图)与pcoa(图)包完成; 浮游生物数据绘图使用软件Origin 2021; 采样点的绘制结合使用ArCIS 10.8和BigMap完成.
2 结果与分析 2.1 南湖修复区水体理化特征4个湖区及参照点水体理化指标及相关性分析详见表 1, A区和B区各项理化指标较为相近, 但DO、TP、TN、NO3--N和NH4+-N浓度与C、D和S区差异显著(P < 0.05或P < 0.01).所有采样点水体pH均值为8.49, 水体整体呈弱碱性; A区和B区的DO依次高于C、D和S区; S区ρ(TP)(0.052 mg·L-1)显著高于A、B、C和D区的(P < 0.05); C、D和S区的TN和NH4+-N浓度显著高于A区和B区.根据文献[22], A区、B区和C区水质达Ⅱ类水平; D区为Ⅲ类水; S区为Ⅳ类水(图 2).未种植沉水植物的S区与4个修复区环境因子差异显著(P < 0.05). S区的TN、TP、DTN和DTP浓度显著高于种植沉水植物的4个湖区(P < 0.05或P < 0.01), 说明沉水植物的种植能在一定程度上降低水体中氮磷营养盐的浓度, 即使在嘉兴温度最低的月份(8℃左右)沉水植物生长代谢速度变慢的情况下也能将水体氮磷浓度维持在一定的水平, 使修复区水体达Ⅲ类水标准.
|
|
表 1 不同湖区水体理化因子均值 Table 1 Mean values of water physicochemical parameters in different regions |

|
图 2 不同采样区水体理化值 Fig. 2 Physical indexes of water body in different areas |
所有采样点共检测出浮游植物6门63属, 其中绿藻门28种, 占种类总种数的44.44%; 硅藻门15种, 占总种数的23.81%; 蓝藻门、隐藻门和裸藻门均为6种, 各占总种数的9.52%; 甲藻门2种为D区特有.4个湖区与对照区门类占比与总体相似, 均以绿藻门与硅藻门为主(图 3). A区共5门35种, 绿藻门占比最多, 特有物种1种; B区共5门33种, 特有物种2种; C区共5门35种, 特有物种7种; D区共6门39种, 特有物种5种; S对照区共鉴定出4门17种, 特有种2种, 4个湖区浮游植物物种数均高于对照区.

|
图 3 不同湖区浮游动植物各门类物种数 Fig. 3 Composition of zooplankton and phytoplankton species in different areas |
浮游动物共检出4门56属, 其中原生动物23种, 占总种数的41.07%; 轮虫17种, 占总种数的30.36%; 桡足类9种, 占总种数的16.07%; 枝角类7种, 占总种数的12.50%.各采样区物种占比与总体相似均以小型浮游动物类型的轮虫与原生动物为主(图 3), A区共4门33种, 特有种5种; B区共4门27种, 特有种3种; C区共4门24种, 特有种1种; D区共4门36种, 特有种9种; S区共3门11种, 特有种4种, 修复区浮游动物物种数均高于对照区, 大型浮游动物枝角类和桡足类物种数明显增多.
2.2.2 生物量与丰度A、B、C和D区浮游植物生物量与丰度占比较为相似(图 4), 裸藻门生物量占比最大, 蓝藻门、绿藻门、硅藻门在丰度上占有优势; S区浮游植物多样性较为单一, 丰度占比主要以绿藻门和硅藻门为主, 绿藻门生物量占比最大.所有采样区浮游动物, 轮虫生物量占据绝对优势; A区原生动物丰度占比大于轮虫, B区和C区原生动物与轮虫丰度占比基本持平, D区原生动物丰度占比小于轮虫, S区原生动物丰度占比最高, A区和D区枝角类生物量占比较其它湖区高(图 4).

|
(a)浮游植物相对生物量, (b)浮游植物相对丰度, (c)浮游动物相对生物量, (d)浮游动物相对丰度 图 4 不同采样区浮游生物相对生物量与相对丰度 Fig. 4 Relative biomass and abundance of plankton in different areas |
4个湖区浮游植物优势种主要为硅藻门、绿藻门和蓝藻门(表 2).不同湖区优势种略有差异, A区共5种优势种, 其中假鱼腥藻(Pseudanabaena sp.)优势度最大; B区共6种优势种, 优势度最高的是假鱼腥藻(Pseudanabaena sp.)和空星藻(Coelastrum sp.), 特有优势种为浮丝藻(Planktothrix sp.)、空星藻(Coelastrum sp.)和螺旋纤维藻(Ankistrodesmus spiralis); C区共9种优势种主要以绿藻门为主, 优势度最高的是细鞘丝藻(Leptolyngbya sp.), 特有优势种为: 细鞘丝藻(Leptolyngbya sp.)、纤维藻(Ankistrodesmus sp.)、肾形藻(Nephrocytium agardhianum)、三角四角藻(Tetraedronoidrae)、平滑四星藻(Tetrastrum sp.)和啮齿隐藻(Cryptomonas erosa); D区共7种优势种, 针杆藻(Synedra sp.)优势度最大, 特有优势种有长胞藻(Dolichospermum sp.)、隐球藻(Aphanocapsa sp.)和弓形藻(Schroederia sp.).此外, 硅藻门的小环藻(Cyclotella sp.)和针杆藻(Synedra sp.)是4个湖区共有的优势种.
|
|
表 2 不同湖区浮游生物优势种及优势度 Table 2 Dominant species and dominance index of plankton in different lake regions |
浮游动物优势种主要为原生动物与轮虫(表 2). A区共6种, 侠盗虫属(Strobilidium sp.)优势度最高; B区共8种, 小巨头轮虫(Cephalodella exigna)优势度最高; C区共6种, 原生动物纤毛虫未定种优势度最高; D区共7种, 小巨头轮虫(Cephalodella exigna)优势度最高.毛板壳虫(Coleps hirtus)和尖头异尾轮虫[Trichocerca (Diurella) tigris]为B区特有优势种; 尖趾单趾轮虫(Monostyla closterocerca)为C区特有优势种; 鳞状叶轮虫(Notholca squamula)、尖尾鞍甲轮虫(Lepadella acuminata)和桡足类无节幼体为D区特有优势种.4个采样区共同优势种有侠盗虫属(Strobilidium sp.)、纤毛虫未定种、疣毛轮属(Synchaeta sp.)和小巨头轮虫(Cephalodella exigna).
2.2.4 生物多样性指数4个沉水植物种植区浮游生物多样性指数无显著性差异(P>0.05), 浮游植物Shannon指数与Pielou指数评价结果为清洁, 显著高于浮游动物轻污染和β-中污染(图 5). Shannon指数主要通过计算不同物种丰度反映各物种间分布的丰富度, Pielou指数反映各物种分布的均匀程度[18, 23], 而浮游植物各门类物种丰度占比相较于浮游动物各门类丰度占比分布得更加均匀(图 4), 浮游动物丰度占比主要以轮虫和原生动物为主, 物种丰富度较为单一, 且各湖区浮游植物的丰度数值高于浮游动物, 因此浮游植物水质评价结果优于浮游动物, 说明浮游植物物种分布均匀度和物种丰富度要高于浮游动物, 而浮游动物群落结构较为简单.

|
图 5 不同采样区浮游动植物生物多样性指数 Fig. 5 Biodiversity index of zooplankton and phytoplankton in different areas |
浮游植物优势种(P1~P17)与环境因子RDA分析显示, 排序轴1和轴2对浮游植物优势种的解释量分别为54.45%和23.56%[图 6(a)].蒙特卡罗检验显著性分析结果显示, NO3--N和DO与浮游植物分布呈极显著相关(P < 0.01), WT和pH与浮游植物分布呈显著相关(P < 0.05). A区和B区浮游植物群落排序特征更为接近, 与pH、WT和DO呈现正相关, 与TP和NO3--N呈负相关; C区浮游植物群落与NH4+-N和NO3--N呈现正相关, 与pH、WT、DO、TP和IP呈负相关; D区浮游植物群落与TP、NO3--N和IP呈正相关, 与DO呈负相关.长胞藻、隐球藻和弓形藻等数量随着TP、NO3--N、IP、pH和WT的增加而升高, 同时受NH4+-N和DO升高的抑制; 假鱼腥藻等数量随着pH、WT和DO的增加而升高, 同时受TP、NO3--N和NH4+-N的抑制.

|
A、B、C和D表示不同湖区; P表示浮游植物优势种; Z表示浮游动物优势种 图 6 浮游生物群落与环境因子的RDA分析 Fig. 6 RDA plot of plankton community and environmental factors |
浮游动物优势种(Z1~Z12)与环境因子RDA分析显示, 排序轴1和轴2对浮游动物优势种的解释量分别为38.87%和20.65%[图 6(b)]. DO和浮游动物分布呈极显著相关(P < 0.01), TP、NO3--N和NH4+-N与浮游植物分布呈显著相关(P < 0.05). A区和B区浮游动物群落排序特征更为接近, 与pH、WT和DO呈现正相关, 与NH4+-N、NO3--N、TP和IP呈负相关; C区浮游动物群落在原点附近分布较为集中, 和各环境因子相关性不是很强; D区浮游动物群与NH4+-N、NO3--N、TP和IP呈正相关, 与pH、WT和DO呈负相关.疣毛轮属、鳞状叶轮虫、尖尾鞍甲轮虫和桡足类无节幼体等数量随着NH4+-N、NO3--N、TP和IP的增加而升高, 同时受pH、WT和DO升高的抑制; 侠盗虫属、钟虫、毛板壳虫和尖头异尾轮虫等数量随着pH、WT和DO的增加而升高, 同时受NH4+-N、NO3--N、TP和IP的抑制.
3 讨论 3.1 不同湖区浮游植物与浮游动物群落结构特征各湖区浮游植物种类组成均以绿藻门和硅藻门为主, 浮游动物均以原生动物和轮虫为主, 这与多位学者对嘉兴河网和南湖水系浮游生物调查结果相类似[13, 24~26].修复区较未修复区浮游生物物种数明显增多且各门类物种数占比发生变化, 未修复区绿藻生物量占优而修复区裸藻门生物量占优, 修复区桡足类和枝角类浮游动物物种数明显增多(图 3和图 4), 说明沉水植物的种植能抑制绿藻的繁殖, 同时有利于大型浮游动物桡足类和枝角类的生长.研究区浮游植物优势种以硅藻门的小环藻与针杆藻占比最多, 硅藻门主要为狭冷性物种.狭冷性物种能够在低温条件下通过调节生理代谢适应外部环境, 具有较高的生态宽位, 从而在低温度且光照强度较弱的条件下取得竞争优势[27].浮游动物优势种以原生动物门类的侠盗虫属、纤毛虫未定种以及轮虫门类的疣毛轮属、小巨头轮虫为主.浮游动物作为水体中杂食性鱼类的食物来源, 枝角类与桡足属于大型浮游动物更容易被捕食, 而轮虫与原生动物属于小型浮游动物具有一定生存优势[28].裸藻作为轮虫喜食的饵料[29], 在浮游植物生物量占比最大[图 4(a)], 因此有利于轮虫的生物量积累从而大量繁殖形成优势种.
4个修复区与南湖中心区水体对流程度不同, 南湖中心区由于游船的扰动、上游高浓度水体流入和沉积物氮磷释放等作用的影响, 与修复区相比, 其水体透明度较低且氮磷浓度较高, 因此修复区与南湖的对流程度越大, 对流区域氮磷值越大、透明度越低. A区和B区相邻, 与南湖中心区相对独立, 对流程度最低, 受南湖中心区水体影响最小; C区呈长条状, 东侧开口与南湖中心区对流程度相对较大; D区南侧开口靠近南湖渡船码头, 游船的扰动作用使其与南湖中心区对流程度最大, 这些生境的差异也会对不同修复区浮游生物群落分布造成影响.
对4个沉水植物种植区浮游生物群落相似性及差异性进行聚类分析与PCoA分析(图 7).浮游植物群落相似性分析显示[图 7(a)和7(c)], A区与B区群落特征较为相似(P>0.05); C区浮游植物分布与其它湖区差异显著(P < 0.05), 这可能与C区进行生态补水有关, 外源水源的输入造成水流波动对浮游植物群落结构产生一定影响; D区各采样点分类不明显, 浮游植物分布组内差异较大组间差异不显著, D区靠近码头, 游船对水体的扰动作用使D区靠近码头的水域和远离码头的水域产生生境异质从而使其浮游植物结构产生差异.浮游动物群落相似性分析显示[图 7(b)和7(d)], A区与D区浮游动物群落结构差异显著(P < 0.05), 其它湖区浮游动物分布较为相似(P>0.05). A区湖面较小、水深较浅且受南湖中心水体对流影响小, 理化因子与D区差异显著(表 1), 生境差异性导致浮游动物群落差异显著.

|
(a)浮游植物丰度聚类; (b)浮游动物丰度聚类; (c)浮游植物PCoA; (d)浮游动物PCoA; P18表示直链藻; P19表示华美十字藻; P20表示囊裸藻; P21表示四角藻; P22表示蹄形藻; P23表示裸藻; Z13表示急游虫; Z14表示鼠异尾轮虫; Z15表示方块鬼轮虫; Z16表示轮虫未定种; Z17表示江苏似铃壳虫; Z18表示螅状独缩虫; Z19表示瓷甲异尾轮虫; Z20表示斜管虫 图 7 不同湖区浮游生物聚类热图与PCoA分析 Fig. 7 Hierarchical clustering tree heat map and PCoA of plankton in different lake regions |
对各湖区浮游动植物丰度、生物量均值和物种数进行分析显示(图 8), 与未种植沉水植物的S区相比, 沉水植物种植区浮游植物生物量显著下降(P < 0.05), 浮游植物与浮游动物物种数显著增加(P < 0.05). S区氮磷浓度显著高于4个修复区(表 1), 氮磷营养水平的初始增加会导致浮游植物种类的增加, 但当营养水平超过某一临界值时, 浮游植物种类减少[30].有研究表明在沉水植物成功修复的富营养化湖泊中浮游植物生物量减少、浮游生物多样性增加[31, 32], 浮游生物各门类物种分布更均匀, 从而有利于提高浮游生物物种丰富度和生态系统稳定性.在4个修复湖区中, B区浮游植物丰度最高; D区浮游植物生物量、浮游动物丰度、浮游动物生物量、浮游动植物物种数是最高的. A、B和C区浮游植物物种数较为相近, A区浮游动物物种数仅次于D区, C区浮游动物物种数最低.此外, A区和D区桡足类浮游动物的生物量占比较其它湖区要高(图 4).同样是30%的沉水植物覆盖率, A区整个湖区均匀种植比B区在岸边高密度种植修复时间更快.综合来看, 四个沉水植物修复区中D区修复效果最好, 其次为A区. D区沉水植物种植覆盖率为60%, A区沉水植物种植覆盖率为30%, A区能用相对较少的沉水植物覆盖率得到较好的修复效果.考虑到经济成本, 沉水植物覆盖率30%, 全湖区均匀种植的方式更适合在在实际工程实施中运用.

|
图 8 各采样区浮游动植物丰度、生物量和物种数 Fig. 8 Abundance, biomass, and number of species of plankton in different lake regions |
水生生物群落分布特征因水体水文条件和水体理化因子的不同而存在差异, 不同湖区影响浮游生物群落分布的主导环境因子也不同[33, 34].RNA分析显示, A区与B区浮游动植物分布主要与DO、WT和pH物理因素有关, 这两个湖区氮磷等营养盐水平较低、溶解氧充足(表 1), 硅藻门优势种较多.浮游生物的生长繁殖均需要消耗DO, 高溶解氧有利于浮游生物生长并形成稳定的生态系统[35].水温影响着浮游生物群落的大小结构和功能, 水温升高有利于小型物种的增殖从而改变浮游生物群落结构, 并增加初级生产力和群落更新速度[36].Gogoi等[37]的研究表明温度升高会导致蓝藻比例的增加, 绿藻和硅藻更适合在较低温度下生存.水体pH在一定范围内增大有利于浮游植物进行光合作用吸收溶解性无机碳[38, 39], 但pH值过大又会抑制浮游植物的光合作用[40], 同时沉水植物的生长也会反过来影响水体酸碱度[41]. C区浮游植物分布与NH4+-N显著正相关, 浮游动物分布受环境因子影响较弱.该区绿藻门优势种占比最多, 这与适宜温度范围内氨氮浓度增高有利于绿藻生长研究相符[42].与其它3个湖区相比C区的氮磷比最高, Zhang等[6]的研究表明高氮磷比环境下更适合绿藻的繁殖. D区浮游动植物群落分布与氮磷营养盐呈明显的正相关关系, 该区氮磷营养盐浓度显著高于其它修复区(表 1).随着氮磷营养盐含量的改变, 水体中的养分极大地影响浮游植物的演替, 刺激浮游植物生物量的增加[37].氮磷营养盐通过对浮游植物的直接影响而对浮游动物群落分布产生间接影响[43]. Li等[44]对太湖浮游生物的研究也表明, 硝氮和氨氮是影响浮游动物分布的重要指标. D区与南湖中心水体对流和游船扰动沉积物氮磷释放等造成的营养盐输入也是影响浮游生物群落分布的重要因素[45].
4 结论(1) 调查期间, 南湖共鉴定出浮游植物6门63属, 以绿藻和硅藻型为主; 共检测出浮游动物4门56属, 主要以原生动物和轮虫这两种小型浮游动物为主, 每个湖区代表优势种不同.
(2) 与未种植沉水植物的区域相比, 4个沉水植物修复区浮游植物生物量显著减少(P < 0.05), 浮游植物与浮游动物的物种数显著增加(P < 0.05), 氮磷营养盐浓度显著下降.
(3) RDA分析表明, 影响浮游植物群落分布的环境因子为NO3--N、DO、WT和pH; 影响浮游动物群落分布的环境因子为DO、NO3--N、NH4+-N和TP.环境因子主要是通过影响浮游植物而对浮游动物产生间接影响.
(4) D区修复效果最好, 其次为A区.考虑到经济成本, 在实际工程实施中更适合采用A区30%的沉水植物覆盖率, 均匀种植的方式对水体进行修复.
致谢: 朱天顺、陈亮、车霏霏、王书航、赵丽、王鹏飞和王雯雯等在写作上提供指导, 段繁曦、刘虹、王晨、张华涛和邢健宇等在采样和实验方面提供帮助, 在此一并致谢!
| [1] | Zhao K, Wang L Z, You Q M, et al. Influence of cyanobacterial blooms and environmental variation on zooplankton and eukaryotic phytoplankton in a large, shallow, eutrophic lake in China[J]. Science of the Total Environment, 2021, 773. DOI:10.1016/j.scitotenv.2021.145421 |
| [2] |
许志, 陈小华, 沈根祥, 等. 上海河道浮游植物群落结构时空变化特征及影响因素分析[J]. 环境科学, 2020, 41(8): 3621-3628. Xu Z, Chen X H, Shen G X, et al. Spatial and temporal variation of phytoplankton community structure and its influencing factors in Shanghai River Channels[J]. Environmental Science, 2020, 41(8): 3621-3628. DOI:10.3969/j.issn.1000-6923.2020.08.043 |
| [3] |
杜彩丽, 杨丽, 赵诣, 等. 淀山湖浮游动物群落时空分布特征及其与环境因子的关系[J]. 环境科学, 2019, 40(10): 4513-4522. Du C L, Yang L, Zhao Y, et al. Temporal and spatial variation of zooplankton community structure and its relationship with environmental factors in Dianshan Lake, Shanghai[J]. Environmental Science, 2019, 40(10): 4513-4522. |
| [4] | Zhang X J, Wang G Q, Tan Z X, et al. Effects of ecological protection and restoration on phytoplankton diversity in impounded lakes along the eastern route of China's South-to-North water diversion project[J]. Science of the Total Environment, 2021, 795. DOI:10.1016/j.scitotenv.2021.148870 |
| [5] | Valentin J L, Leles S G, Tenenbaum D R, et al. Frequent upwelling intrusions and rainfall events drive shifts in plankton community in a highly eutrophic estuary[J]. Estuarine, Coastal and Shelf Science, 2021, 257. DOI:10.1016/j.ecss.2021.107387 |
| [6] | Zhang H H, Zong R R, He H Y, et al. Biogeographic distribution patterns of algal community in different urban lakes in China: insights into the dynamics and co-existence[J]. Journal of Environmental Sciences, 2021, 100: 216-227. DOI:10.1016/j.jes.2020.07.024 |
| [7] |
王振方, 张玮, 杨丽, 等. 异龙湖不同湖区浮游植物群落特征及其与环境因子的关系[J]. 环境科学, 2019, 40(5): 2249-2257. Wang Z F, Zhang W, Yang L, et al. Characteristics of phytoplankton community and its relationship with environmental factors in different regions of Yilong Lake, Yunnan Province, China[J]. Environmental Science, 2019, 40(5): 2249-2257. |
| [8] | Xia T, Zhu L, Liu Z N, et al. Construction of symbiotic system of filamentous algae and submerged plants and its application in wastewater purification[J]. Journal of Water Process Engineering, 2021, 43. DOI:10.1016/j.jwpe.2021.102240 |
| [9] | Chao C X, Wang L G, Li Y, et al. Response of sediment and water microbial communities to submerged vegetations restoration in a shallow eutrophic lake[J]. Science of the Total Environment, 2021, 801. DOI:10.1016/j.scitotenv.2021.149701 |
| [10] | Fu H, Özkan K, Yuan G X, et al. Abiotic and biotic drivers of temporal dynamics in the spatial heterogeneity of zooplankton communities across lakes in recovery from eutrophication[J]. Science of the Total Environment, 2021, 778. DOI:10.1016/j.scitotenv.2021.146368 |
| [11] | Zhang X B, Guo K D, Lu C, et al. Effects of origin and water depth on morphology and reproductive modes of the submerged plant Vallisneria natans[J]. Global Ecology and Conservation, 2020, 24. DOI:10.1016/j.gecco.2020.e01330 |
| [12] | Barrow J L, Beisner B E, Giles R, et al. Macrophytes moderate the taxonomic and functional composition of phytoplankton assemblages during a nutrient loading experiment[J]. Freshwater Biology, 2019, 64(8): 1369-1381. DOI:10.1111/fwb.13311 |
| [13] |
姜登岭, 赵昊, 邬喜红, 等. 嘉兴城市河网区高、低水位期浮游植物群落及其与环境因子的典范对应分析[J]. 环境化学, 2020, 39(9): 2540-2550. Jiang D L, Zhao H, Wu X H, et al. Model correspondence analysis of phytoplankton community and its environmental factors at high and low water levels in urban river network area of Jiaxing City[J]. Environmental Chemistry, 2020, 39(9): 2540-2550. |
| [14] |
王书航, 郑朔方, 尚晓, 等. 平原河网景观湖泊水质提升关键问题分析与对策研究——以嘉兴南湖为例[J]. 环境工程技术学报, 2020, 10(6): 891-896. Wang S H, Zheng S F, Shang X, et al. Key issues of water quality improvement in the landscape lake of plain river network and corresponding countermeasures: a case study of Nanhu Lake in Jiaxing City[J]. Journal of Environmental Engineering Technology, 2020, 10(6): 891-896. |
| [15] | 国家环境保护总局. 水和废水监测分析方法[M]. ((第四版)). 北京: 中国环境科学出版社. |
| [16] | 胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [17] | 赵文. 水生生物学[M]. 北京: 中国农业出版社, 2005. |
| [18] |
孙军, 刘东艳. 多样性指数在海洋浮游植物研究中的应用[J]. 海洋学报, 2004, 26(1): 62-75. Sun J, Liu D Y. The application of diversity indices in marine phytoplankton studies[J]. Acta Oceanologica Sinica, 2004, 26(1): 62-75. |
| [19] | Huang Z Q, Shi X N. Correlation between ecotourism and the evolution law of zooplankton communities in Dongting Lake tourist area of China's Hunan[J]. Applied Ecology and Environmental Research, 2020, 18(2): 2959-2974. DOI:10.15666/aeer/1802_29592974 |
| [20] | Lv H, Yang J, Liu L M, et al. Temperature and nutrients are significant drivers of seasonal shift in phytoplankton community from a drinking water reservoir, subtropical China[J]. Environmental Science and Pollution Research, 2014, 21(9): 5917-5928. DOI:10.1007/s11356-014-2534-3 |
| [21] | Liu X, Lu X H, Chen Y W. The effects of temperature and nutrient ratios on Microcystis blooms in Lake Taihu, China: an 11-year investigation[J]. Harmful Algae, 2011, 10(3): 337-343. DOI:10.1016/j.hal.2010.12.002 |
| [22] | GB 3838-2002, 地表水环境质量标准[S]. |
| [23] |
杨潇, 马吉顺, 张欢, 等. 鄱阳湖不同水文期浮游生物群落结构特征和影响因素及水质评价[J]. 水生生物学报, 2021, 45(5): 1093-1103. Yang X, Ma J S, Zhang H, et al. Community structure and the water quality during different hydrological periods in Poyang Lake[J]. Acta Hydrobiologica Sinica, 2021, 45(5): 1093-1103. |
| [24] |
马迎群, 迟明慧, 赵艳民, 等. 嘉兴南湖浮游动物群落结构及影响因素研究[J]. 环境科学与技术, 2019, 42(12): 180-186. Ma Y Q, Chi M H, Zhao Y M, et al. Characteristics of zooplankton groups in Nanhu Lake catchment and its influencing factors in Jiaxing city[J]. Environmental Science & Technology, 2019, 42(12): 180-186. |
| [25] |
马迎群, 赵艳民, 迟明慧, 等. 嘉兴南湖水系浮游植物群落结构及其环境影响因子[J]. 水生态学杂志, 2021, 42(2): 42-48. Ma Y Q, Zhao Y M, Chi M H, et al. Phytoplankton community structure and its relationship with environmental factors in the Nanhu Lake catchment, Jiaxing city[J]. Journal of Hydroecology, 2021, 42(2): 42-48. |
| [26] |
季宏康, 范帆, 王宝印, 等. 嘉兴南湖水系水污染特征分析及治理策略[J]. 环境污染与防治, 2018, 40(5): 581-587. Ji H K, Fan F, Wang B Y, et al. Analysis and countermeasures of water pollution characteristics in South Lake water system of Jiaxing[J]. Environmental Pollution & Control, 2018, 40(5): 581-587. |
| [27] |
王司阳, 张笑欣, 田世民, 等. 黄河流域干旱区湖泊冰封期浮游植物群落结构特征及影响因子研究[J]. 水利学报, 2020, 51(9): 1070-1079. Wang S Y, Zhang X X, Tian S M, et al. Study on phytoplankton community structure characteristics and its influencing factors of lakes in arid regions of the Yellow River basin during ice-sealing period[J]. Journal of Hydraulic Engineering, 2020, 51(9): 1070-1079. |
| [28] | Ersoy Z, Brucet S, Bartrons M, et al. Short-term fish predation destroys resilience of zooplankton communities and prevents recovery of phytoplankton control by zooplankton grazing[J]. PLoS One, 2019, 14(2). DOI:10.1371/journal.pone.0212351 |
| [29] |
周绪申, 李娜, 孙博闻, 等. 白洋淀浮游生物群落结构的季节变化及其与环境因子的关系[J]. 水利水电技术(中英文), 2021, 52(8): 110-119. Zhou X S, Li N, Sun B W, et al. Seasonal variation of plankton community structure in the Lake Baiyangdian and the relationship with environmental factors[J]. Water Resources and Hydropower Engineering, 2021, 52(8): 110-119. |
| [30] | Dembowska E A, Napiórkowski P, Mieszczankin T, et al. Planktonic indices in the evaluation of the ecological status and the trophic state of the longest lake in Poland[J]. Ecological Indicators, 2015, 56: 15-22. |
| [31] | Peng X, Yi K L, Lin Q W, et al. Annual changes in periphyton communities and their diatom indicator species, in the littoral zone of a subtropical urban lake restored by submerged plants[J]. Ecological Engineering, 2020, 155. DOI:10.1016/j.ecoleng.2020.105958 |
| [32] |
苏小妹, 薛庆举, 万翔, 等. 小型湖泊修复区与未修复区对生态修复的响应与评价[J/OL]. 中国环境科学, 1-12. https://doi.org/10.19674/j.cnki.issn1000-6923.20210803.001, 2021-10-22. Su X M, Xue Q J, Wan X, et al. Responses and assessment of water ecological status in restored and unrestored areas of a small urban lake after ecological restoration[J/OL]. China Environmental Science, 1-12. https://doi.org/10.19674/j.cnki.issn1000-6923.20210803.001, 2021-10-22. |
| [33] | Zhao C S, Pan X, Yang S T, et al. Principal hydrology and water quality factors driving the development of plankton communities in a pilot city of China[J]. Ecohydrology, 2020, 13(4). DOI:10.1002/eco.2207 |
| [34] | Rao K, Zhang X, Wang M, et al. The relative importance of environmental factors in predicting phytoplankton shifting and cyanobacteria abundance in regulated shallow lakes[J]. Environmental Pollution, 2021, 286. DOI:10.1016/j.envpol.2021.117555 |
| [35] |
陈莎, 谢青, 付梅, 等. 三峡库区典型支流水库浮游动植物群落结构特征及其与环境因子的关系[J]. 环境科学, 2021, 42(5): 2303-2312. Chen S, Xie Q, Fu M, et al. Structural characteristics of zooplankton and phytoplankton communities and its relationship with environmental factors in a typical tributary reservoir in the Three Gorges Reservoir Region[J]. Environmental Science, 2021, 42(5): 2303-2312. |
| [36] | Rasconi S, Gall A, Winter K, et al. Increasing water temperature triggers dominance of small freshwater plankton[J]. PLoS One, 2015, 10(10). DOI:10.1371/journal.pone.0140449 |
| [37] | Gogoi P, Das S K, Das Sarkar S, et al. Environmental factors driving phytoplankton assemblage pattern and diversity: insights from Sundarban eco-region, India[J]. Ecohydrology & Hydrobiology, 2021, 21(2): 354-367. |
| [38] |
马煜, 陆欣鑫, 范亚文. 松花江哈尔滨段浮游植物群落格局及其与环境因子的相关性[J]. 生态学报, 2021, 41(1): 224-234. Ma Y, Lu X X, Fan Y W. Correlation between phytoplankton community patterns and environmental factors in Harbin section of the Songhua River[J]. Acta Ecologica Sinica, 2021, 41(1): 224-234. |
| [39] |
马宝珊, 徐滨, 魏开金, 等. 安宁河中游浮游植物群落结构及其与环境因子的关系[J]. 生态学杂志, 2020, 39(10): 3332-3341. Ma B S, Xu B, Wei K J, et al. Phytoplankton community structure and its relation to environmental conditions in the middle Anning River, China[J]. Chinese Journal of Ecology, 2020, 39(10): 3332-3341. |
| [40] |
路枫, 李磊, 齐青松, 等. 哈尔滨城市河网丰水期浮游植物群落分布特征及驱动因子[J]. 环境科学, 2021, 42(7): 3253-3262. Lu F, Li L, Qi Q S, et al. Distribution characteristics and influencing factors of phytoplankton community in Harbin Urban River Network during wet season[J]. Environmental Science, 2021, 42(7): 3253-3262. |
| [41] | Peng X, Lin Q W, Liu B Y, et al. Effect of submerged plant coverage on phytoplankton community dynamics and photosynthetic activity in situ[J]. Journal of Environmental Management, 2022, 301. DOI:10.1016/j.jenvman.2021.113822 |
| [42] |
蒋雪蕾, 周晓见, 林佳宁, 等. 黄海浒苔绿潮生态效应研究进展[J]. 海洋环境科学, 2021, 40(4): 647-652. Jiang X L, Zhou X J, Lin J N, et al. Research progress in the ecological consequences of Ulva prolifera green tides in the Yellow Sea[J]. Marine Environmental Science, 2021, 40(4): 647-652. |
| [43] |
陈佳琪, 赵坤, 曹玥, 等. 鄱阳湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 2020, 40(18): 6644-6658. Chen J Q, Zhao K, Cao Y, et al. Zooplankton community structure and its relationship with environmental factors in Poyang Lake[J]. Acta Ecologica Sinica, 2020, 40(18): 6644-6658. |
| [44] | Li C C, Feng W Y, Chen H Y, et al. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu-a large freshwater lake in China[J]. Environmental Pollution, 2019, 245: 1050-1057. |
| [45] |
张晶晶, 王玉珏, 李凡, 等. 小清河专项治理对莱州湾环境和浮游植物的影响[J]. 环境科学, 2022, 43(4): 1997-2006. Zhang J J, Wang Y J, Li F, et al. Effects of pollution control of Xiaoqing River on environment factors and phytoplankton community in the Laizhou Bay[J]. Environmental Science, 2022, 43(4): 1997-2006. |