 2022, Vol. 43
2022, Vol. 43

2. 中国科学院大学, 北京 100049;
3. 广东省农业科学院农业资源与环境研究所, 广州 510640;
4. 湖南省宁乡市农业技术推广中心, 宁乡 410600
 , ZHENG Xiao-dong1,3 , MAO Wan-qiong1,2 , CHEN Xiang-bi1
, ZHENG Xiao-dong1,3 , MAO Wan-qiong1,2 , CHEN Xiang-bi1 , HU Ya-jun1 , CHENG Ai-wu4 , HE Xun-yang1 , SU Yi-rong1
, HU Ya-jun1 , CHENG Ai-wu4 , HE Xun-yang1 , SU Yi-rong1
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Institute of Agricultural Resources and Environment, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China;
4. Agricultural Technology Promotion Center, Ningxiang 410600, China
土壤是陆地生态系统最大的碳库, 在全球碳循环中发挥着至关重要的作用[1].增加土壤有机碳的含量和稳定性是缓解全球气候变化、提升农田土壤肥力的重要途径[2].植物残体是土壤有机碳的主要来源, 微生物是驱动植物残体转化为土壤有机碳的引擎[3, 4].植物残体进入土壤后, 在微生物作用下从大分子聚合物逐级降解为小分子化合物, 并持续释放出可溶性有机碳(dissolved organic carbon, DOC)[5, 6].DOC被微生物吸收后, 一部分被矿化为气态碳离开土壤, 一部分经合成代谢被同化为细胞组成物质, 经生长、繁殖和死亡等迭代过程, 不断形成微生物源稳定有机碳在土壤中积累[7, 8].因此, 深入了解土壤有机碳转化和参与这一过程的微生物对调节土壤碳库至关重要.
真菌和细菌是土壤微生物的两大类群[9].由于自身元素组成以及生存策略的不同, 细菌和真菌对碳源底物的利用具有偏好性[10, 11].细菌在利用小分子化合物时占优势, 在植物残体降解前期占主导; 真菌利用大分子聚合物时具有优势, 在植物残体降解后期占主导[12].然而这种规律并不绝对, 有研究发现, 当葡萄糖添加量(以C/土计)为16~32 mg ·kg-1时, 真菌吸收葡萄糖比细菌快[13]; 微氧条件下, 枯草芽孢杆菌(Bacillus subtilis)在木质素降解方面比真菌更有优势[14].因此, 土壤真菌和细菌在植物残体转化为土壤有机碳过程中的作用强度与规律受控于具体的环境条件.
稻田和旱地作为我国东部粮食主产区两种主要的土地利用方式, 其土壤水分条件呈鲜明对比[15].稻田长期处于淹水条件, 土壤微生物类群以厌氧和兼性厌氧为主, 旱地主要处于自然排干的好氧条件, 表层土壤微生物类群以好氧微生物为主[16].厘清这两类巨大反差环境条件的土壤中有机质微生物降解的过程差异对农田土壤碳库管理有重要意义.本课题组前期研究表明, 我国东部4个气候区稻田土壤有机碳含量均显著低于旱地, 稻田土壤长期固持的有机碳中28% ~36%来自微生物残留物, 显著低于旱地(40% ~59%), 且稻田土壤中真菌/细菌残留物比值均低于旱地土壤[17].这表明, 稻田土壤微生物(尤其是真菌)合成代谢作用对有机碳积累的参与度低于旱地土壤.该结论是基于土壤中长期稳定的生物标志物(氨基糖和木质素酚)分析得出, 短期内细菌和真菌对秸秆类新鲜有机碳的利用是否符合上述规律, 尚不清楚.
本研究以亚热带典型稻田和旱地土壤为对象, 以13C标记的秸秆为材料, 通过短期(40 d)室内模拟培养试验结合13C-CO2监测和活性微生物(13C-PLFA)分析, 研究13C-秸秆在稻田和旱地土壤中的矿化动态, 解析真菌和细菌利用13C-秸秆的过程差异, 以期为深入揭示稻田和旱地土壤有机碳的稳定性差异及其微生物机制提供科学依据.
1 材料与方法 1.1 供试土壤和秸秆土壤样品采自湖南省常德市鼎城区长岭岗乡(24°56′~24°57′N, 108°03′~108°04′E)的典型稻田和旱地, 土壤母质为第四纪红土.稻田中水稻处于幼苗期(株高约5 cm), 未施肥; 旱地中油菜刚收割, 未施肥.采集表层土壤(0~15 cm), 过10目筛后备用.土壤基本理化性质见表 1.
|
|
表 1 土壤基本性质 Table 1 Basic properties of soils |
供试秸秆为13C标记的玉米秸秆.玉米幼苗置于以13C-CO2为碳源的标记箱中生长60 d后收获, 具体标记方法参考专利: ZL200610019742.4.除去根系, 玉米秸秆105℃下杀青30 min, 60℃烘干至恒重.将玉米秸秆剪成小段, 粉碎研磨, 过100目筛.秸秆碳含量为43.8%, 全氮含量为1.7%, 碳氮比为26, δ13C值为1 662‰.
1.2 试验设计与样品采集稻田和旱地土壤分别设置不添加秸秆的对照处理和13C-秸秆添加处理(添加量以C/土计, 为150 mg ·kg-1), 共4个处理, 分别表示为稻田、稻田+秸秆、旱地和旱地+秸秆, 每个处理4次重复; 本试验不考虑同位素分馏以及激发效应, 因而不设置添加非标记秸秆处理.称取相当于烘干土重50 g的鲜土装于自封袋中, 秸秆粉末多点添加, 密封袋口反复揉搓, 混匀后无损转至平底试管(直径40 mm, 高度120 mm)内.平底试管置于1 L的密闭大广口瓶中, 瓶口用带三通阀的橡胶塞塞紧.广口瓶底部加2 mL去CO2水, 以维持瓶内空气饱和湿度.稻田土壤保持淹水状态(试管内水层深度为1~2 cm), 旱地土壤湿度调节到45%田间持水量, 25℃黑暗条件下培养.于培养的第0.25、1、2、3、5、7、10、15、20、25、30、35和40 d采集气体样品, 于培养的第2、7和40 d进行土壤破坏性采样.
1.3 样品分析土壤基本理化性质采用常规方法分析[18].CO2含量采用气相色谱仪GC-6890 (Agilent Technologies, 美国)测定, CO2中的δ13C值采用稳定性同位素比值质谱仪MAT253(Thermo Scientific, 美国)测定[19].DOC含量采用0.05 mol ·L-1硫酸钾提取, 碳-自动分析仪Phoenix8000测定, 取部分浸提液冷冻干燥后测定δ13C值[20].微生物群落结构用土壤磷脂脂肪酸(phospholipid fatty acid, PLFA)表示.根据改进的Buyer方法提取土壤PLFA, 采用连接美国MIDI公司PLFA数据库的气相色谱仪GC-6890(Agilent Technologies, 美国)对提取物进行定量和定性分析[21]. i15:0、a15:0、i16:0、i17:0、a17:0指示革兰氏阳性菌(G+), 16:1ω9c、16:1ω7c、16:1ω5c、18:1ω7c、18:1ω5c、cy17:0、cy19:0指示革兰氏阴性菌(G-), 10Me16:0、10Me17:0, 10Me18:0指示放线菌, 18:1ω9c和18:2ω6c指示真菌[11, 19].细菌为革兰氏阳性菌、革兰氏阴性菌以及放线菌的总和.PLFA提取物对应的δ13C值采用气相色谱-燃烧-同位素比值质谱仪GC(Thermo Scientific Trace GC Ultra)-C-IRMS(MAT253)测定.
1.4 计算方法土壤释放CO2中来源于秸秆的比例[21]为:
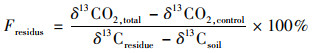
|
(1) |
式中, δ13CO2, total和δ13CO2, control分别为添加和不添加秸秆处理释放CO2的δ13C值, δ13Cresidue和δ13Csoil分别为秸秆和土壤的δ13C值.
以13C-CO2排放速率[13C-CO2, mg ·(kg ·d)-1]表示秸秆碳矿化速率:
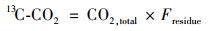
|
(2) |
式中, CO2, total为添加秸秆处理的总CO2排放速率.
以CO2中来自秸秆的碳累积量占添加碳总量比例估算秸秆矿化率.应用相似公式计算DOC中来自秸秆的碳含量(13C-DOC).
各类群微生物的PLFA含量为该类群单体PLFA含量之和.
每种单体PLFA中秸秆碳所占比例[16, 20](Fi, %)为:
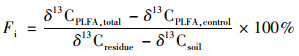
|
(3) |
式中, δ13CPLFA, total和δ13CPLFA, control分别为添加秸秆处理和对照处理中单体PLFA的δ13C值.
每种单体PLFA中来自秸秆的碳含量(Pi, nmol ·g-1)为:
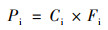
|
(4) |
式中, Ci为单体PLFA中C含量(nmol ·g-1).
各类群微生物PLFA中秸秆碳的同化量(13C-PLFA)为该类群单体PLFA中来自秸秆的碳含量之和.
各类群微生物的秸秆碳周转速率[22](Tr, % ·d-1)为:
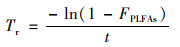
|
(5) |
式中, FPLFAs为秸秆碳的富集比例, 即该类群PLFA中13C占该类群总碳量的比例(%)[16], t为培养时间(d).
1.5 统计分析采用Excel 2016和SPSS 26.0进行数据正态性和方差齐性检验, 不同土地利用方式和不同秸秆处理间差异采用双因素方差分析检验, Duncan法进行多重比较(α=0.05).采用Origin2021绘制图表.数据表示为平均值±标准差.
2 结果与分析 2.1 秸秆碳矿化特征培养期间, 稻田和旱地的对照处理(不添加秸秆)CO2累积排放量分别为295.4 mg ·kg-1和383.5 mg ·kg-1; 秸秆添加使CO2累积排放量分别增加了40%和44%[图 1(a)].两种土壤的秸秆碳矿化速率(13C-CO2排放速率)均表现为先升高后降低最后趋于平缓, 且最大矿化速率均出现在第5 d.培养0.25~1 d, 稻田土壤中秸秆矿化速率[1.2~1.68 mg ·(kg ·d)-1, 以CO2计, 下同]显著高于旱地[0.41~0.45 mg ·(kg ·d)-1](P < 0.05). 2~20 d, 旱地中秸秆碳矿化速率高于稻田. 20 d后, 秸秆碳矿化速率在两种土壤中的差异不显著[图 1(b)].培养结束时, 稻田中秸秆碳的累积矿化率(11%)约为旱地(20%)的1/2[图 1(c)].

|
*表示稻田和旱地间差异显著(P < 0.05) 图 1 稻田和旱地土壤CO2累积排放量、秸秆的矿化速率和累积矿化率 Fig. 1 Cumulative emission of total CO2, mineralization rate, and proportion of the crop residue in paddy and upland soils |
土地利用方式和秸秆处理对DOC总含量无显著交互作用.培养期间, 稻田和旱地土壤中ω(总DOC)范围分别为23.4~51.3 mg ·kg-1和27.5~61.5mg ·kg-1, 总体以稻田低于旱地[图 2(a)].秸秆添加对DOC总含量无显著影响.培养期间, 两种土壤中秸秆碳转化为13C-DOC的量均呈明显上升趋势; 培养结束时, 稻田ω(13C-DOC)(0.39 mg ·kg-1)低于旱地(0.49 mg ·kg-1)[图 1(b), P < 0.05].

|
*表示相同处理下稻田和旱地间差异显著(P < 0.05) 图 2 稻田和旱地土壤中DOC总量和13C-DOC含量 Fig. 2 Contents of total DOC and 13C-DOC in paddy and upland soils |
培养期间, 各微生物类群总PLFA含量均以稻田高于旱地(图 3).培养前期(2 d), 稻田土壤中, 秸秆添加减少G-和放线菌含量, 但对PLFA总量无显著影响; 旱地土壤中, 秸秆添加增加真菌、放线菌、细菌含量和总PLFA量.培养中期(7 d), 秸秆添加对稻田土壤各类群微生物PLFA含量无显著影响; 旱地土壤中, 秸秆添加增加G+和放线菌PLFA.培养后期(40 d), 稻田土壤秸秆添加显著减少G+和细菌PLFA; 旱地土壤中秸秆添加显著增加细菌PLFA.

|
G+表示革兰氏阳性菌, G-表示革兰氏阴性菌; 不同小写字母表示土壤和秸秆处理交互作用显著下处理间差异显著(P < 0.05); *表示相同处理下稻田与旱地间差异显著(P < 0.05); #表示同一土壤中对照与秸秆添加处理间差异显著(P < 0.05) 图 3 不同处理下土壤微生物的特征PLFAs含量 Fig. 3 Contents of microbial PLFAs in upland and paddy soils with or without crop residues |
总体而言, 土地利用方式对细菌/真菌和G+/G-比值有显著影响(表 2).同一处理的细菌/真菌比值以稻田略高于旱地, 且不同时间点上变化幅度也以稻田(0.2~0.5)小于旱地(0.6~1).培养期间, 同一处理的G+/G-比值均以稻田低于旱地(P < 0.05); 整个培养期, 稻田对照处理的G+/G-比值随时间的变化不明显, 旱地对照处理呈上升趋势; 两种土壤中, 添加秸秆处理的G+/G-比值随培养时间的延长均呈上升趋势.
|
|
表 2 培养过程中不同处理土壤细菌/真菌、革兰氏阳性菌/革兰氏阴性菌比值的变化1) Table 2 Ratios of bacteria to fungi and gram-positive to gram-negative bacteria under different treatments during incubation |
2.4 利用秸秆碳的微生物群落组成及变化
如图 4(a)所示, 两种土壤中秸秆碳同化为微生物PLFA的总量(13C-PLFA总量)及各类群13C-PLFA含量均呈先降低后升高的趋势.培养前期(2 d), 13C-PLFA总量以稻田显著高于旱地(P < 0.05), 之后两者差异不显著.培养期内, 13C-真菌PLFA表现为稻田低于旱地; 13C-细菌、13C-G+和13C-G-的PLFA量均表现为稻田高于旱地.培养前期, 13C-放线菌PLFA以稻田高于旱地, 培养后期(40 d)则相反.

|
横坐标数字表示时间(d); G+表示革兰氏阳性菌, G-表示革兰氏阴性菌; *表示同一时间同一类群PLFA中稻田与旱地间差异显著(P < 0.05) 图 4 旱地和稻田土壤中PLFA秸秆碳的转化量、富集比例和周转速率 Fig. 4 Amount of 13C-PLFA, enrichment ratio of 13C in PLFA, and turnover rate of 13C in microbial PLFA in paddy and upland soils |
秸秆碳在PLFA总量和真菌PLFA中的富集比例均以稻田低于旱地[图 4(b)].培养前期, 总细菌、G+、G-和放线菌的13C-PLFA富集比例均表现为稻田高于旱地, 之后细菌、G+和G-的13C-PLFA富集比例在旱地和稻田间的差异不显著, 培养后期, 放线菌PLFA中13C的富集比例则以稻田低于旱地.总量和真菌PLFA中13C周转速率以稻田低于旱地[图 4(c)]; 细菌、G+、G-和放线菌的13C周转速率的变化规律与对应的富集比例一致.
3 讨论通常, 淹水引起的厌氧环境抑制微生物活性, 使稻田比旱地土壤有机碳矿化速率低[19].本研究培养0.25~1 d时, 稻田土壤中秸秆碳矿化速率高于旱地土壤[图 1(b)], 这可能是由两种土壤利用秸秆碳的主导微生物类群不同引起. 13C-PLFA分析表明稻田土壤中利用秸秆碳的主要微生物类群为细菌, 旱地土壤中则为真菌(图 5).一般而言, 细菌的碳源利用效率低于真菌[23], 即同化一单位秸秆碳释放的CO2量以细菌高于真菌, 意味着以细菌主导的稻田土壤微生物对新鲜碳源的利用效率低于以真菌主导的旱地土壤.由于稻田土壤微生物对秸秆碳的同化量大于旱地土壤[图 4(a)], 稻田土壤释放的CO2也比旱地土壤多. 2 d后, 随着秸秆输入的DOC被消耗[图 2(a)], 微生物矿化纤维素、木质素等组分需经过氧化、脱水以及一系列的自由基链反应[24], 旱地土壤的好氧环境比稻田土壤的淹水环境更有利, 因而旱地土壤中秸秆碳矿化速率高于稻田土壤(图 1).

|
*表示同一时间点同一类群PLFA的旱地和稻田间差异显著(P < 0.05) 图 5 稻田和旱地土壤中利用秸秆碳的各类群PLFA的相对丰度 Fig. 5 Relative abundance of 13C incorporated in microbial PLFAs in paddy and upland soils |
根据自身生存策略, 可将土壤微生物划分为r型和K型[3].r型微生物生长快, 一般专性利用易分解碳源; K型微生物生长慢, 可兼性利用易分解碳源和难分解碳源[25, 26].有研究认为, G-细菌是r型微生物[27], 在秸秆降解的前期占主导; 而真菌、放线菌和大部分G+细菌是K型微生物[12, 20], 在秸秆降解的后期占主导.然而, 本研究结果与上述规律不完全一致.秸秆添加2 d, 旱地土壤中真菌和放线菌含量显著增加且细菌/真菌比值显著降低(图 3和表 2), 说明旱地土壤中真菌和放线菌是对新鲜碳源反应较灵敏的微生物类群, 而稻田土壤中对添加秸秆反应较灵敏的是G+细菌和放线菌.即旱地和稻田土壤中对新鲜碳源输入反应最快的均为K型微生物.这可能是由于本研究输入的玉米秸秆C/N比更接近K型微生物生物量C/N比, 大大刺激K型微生物(尤其是真菌)生长[26].放线菌对秸秆添加响应灵敏, 但其对秸秆碳的同化量、周转速率均远低于真菌和G+细菌(图 4), 造成放线菌对秸秆碳利用的贡献小(图 5, 占13C-PLFA总量的1% ~5%).G+/G-反映土壤微生物受到的饥饿压力, 其值越高表明土壤养分越贫瘠[21].旱地土壤的G+/G-始终高于稻田土壤(表 2), 说明旱地土壤中养分不如稻田土壤丰富.细菌/真菌比值可表征土壤生态系统的稳定性[28].秸秆添加后, 旱地土壤微生物对新鲜碳源的竞争激烈, 细菌/真菌比值变化幅度大; 相比之下, 稻田土壤本身SOC含量较高、养分较充足(表 1), 加上微生物活性受到淹水抑制(图 4), 微生物类群之间的竞争活动较弱, 群落结构更稳定, 因而细菌/真菌变化幅度小(表 2).培养结束时, 尽管稻田土壤中微生物总PLFA含量高于旱地(图 3), 但两种土壤中微生物的秸秆碳同化总量相当(图 4), 说明稻田土壤中参与秸秆碳降解的活性微生物占比少[29].整个培养期稻田土壤中真菌对秸秆碳的富集比例远低于旱地(图 5), 细菌对秸秆碳的富集比例在培养中后期与旱地土壤相似(图 4), 说明稻田淹水对真菌的抑制程度强于细菌.
本研究发现, 旱地土壤中真菌在秸秆分解中发挥主要作用, 而稻田土壤中G+和G-细菌是参与秸秆分解的主要微生物类群(图 5).残体本身的性质和土壤环境的不同会导致参与植物残体分解微生物群落不同[25, 27].本研究供试秸秆来源和添加量一致, 供试土壤来自同一成土母质, 因此底物丰缺和水分环境差异可能是旱地和稻田土壤中新鲜碳源微生物利用主要类群差异的主因(图 4).具体而言, 旱地土壤中SOC含量较稻田土壤低, 前者微生物受到更强的碳源限制, 当秸秆加入后, 微生物对外源碳的利用能力较稻田土壤微生物强[图 4(a), 旱地总量PLFA的周转速率大于稻田][30].秸秆碳在旱地土壤真菌PLFA中的周转速率是稻田土壤的2.2~8.9倍[图 4(c)], 被微生物同化秸秆碳在真菌PLFA的分配比例高达76%(图 5).这与前人旱作好氧条件下真菌主导外源有机碳利用和固持的结论相符[16].绝大多数真菌是好氧的、细菌则包括好氧、厌氧和兼性好氧类群[16], 因而淹水对真菌的抑制作用大于细菌, 导致稻田土壤中真菌PLFA低于G+、G-和放线菌(图 3), 进而抑制真菌对秸秆的矿化分解, 使细菌成为稻田中降解秸秆的主导菌群(图 4和图 5).细菌和真菌新陈代谢和细胞功能不同[31], 由于细菌生物量C/N比(4 ∶1)低于真菌(10 ∶1)[28], 前者养分需求高, 自身生物量周转时间为2.3~33 d; 后者养分需求低, 周转时间为130~150 d[29].因此, 以细菌主导秸秆碳降解过程的稻田土壤中, 微生物对秸秆碳的固持时间短于以真菌主导的旱地土壤.与旱地土壤相比, 稻田土壤中秸秆碳较低的矿化率(图 1)及相当的微生物同化量(图 3), 说明淹水条件下较多秸秆没有被微生物分解代谢也没有被用于合成代谢, 更多的秸秆碳可能直接以土壤物理化学保护的形式沉积在土壤中[4, 6].
本课题组前期基于区域调研结合生物标志物(氨基糖和木质素酚)的研究也证实了稻田土壤中积累的植物源有机碳高于旱地土壤, 而源于微生物合成代谢产物的微生物残留物比旱地土壤低, 尤其是真菌残留物[17].有研究表明, 细菌死亡后其残留物(生物标志物为胞壁酸)稳定性比真菌残留物(生物标志物为氨基葡萄糖)低[3, 32].因而, 本研究稻田土壤秸秆降解过程由细菌主导而旱地由真菌主导的结果, 暗示了稻田土壤中残留的微生物源碳(细菌残留物为主)稳定性低于旱地土壤(真菌残留物为主).前人基于有机质分组(颗粒有机质和矿物结合态有机质)的研究发现, 不稳定的颗粒有机碳占比以稻田高于相邻旱地, 而稳定性较高的矿物结合态有机碳占比以旱地高于稻田[33].总之, 本研究短期监测结果和以往长期有机碳积累组分分析均表明, 稻田土壤有机碳积累能力强是缘于淹水抑制了微生物(尤其是真菌)矿化, 但其积累的有机碳稳定性比旱地土壤弱, 警示了稻田淹水管理对保护土壤碳库的重要性.
4 结论稻田和旱地土壤中参与秸秆降解的微生物类群差异明显, 稻田土壤主要以细菌为主, 而旱地土壤以真菌为主.尽管稻田土壤中真菌、细菌生物量大于旱地土壤, 但淹水抑制了稻田土壤好氧微生物(尤其是真菌)的活动, 导致真菌对秸秆碳利用的参与度低于旱地, 使稻田土壤中作物残体矿化为CO2的程度弱于旱地土壤.
| [1] | Lal R. Soil carbon sequestration to mitigate climate change[J]. Geoderma, 2004, 123(1-2): 1-22. DOI:10.1016/j.geoderma.2004.01.032 |
| [2] | Liang C, Schimel J P, Jastrow J D. The importance of anabolism in microbial control over soil carbon storage[J]. Nature Microbiology, 2017, 2(8). DOI:10.1038/nmicrobiol.2017.105 |
| [3] |
汪景宽, 徐英德, 丁凡, 等. 植物残体向土壤有机质转化过程及其稳定机制的研究进展[J]. 土壤学报, 2019, 56(3): 528-540. Wang J K, Xu Y D, Ding F, et al. Process of plant residue transforming into soil organic matter and mechanism of its stabilization: a review[J]. Acta Pedologica Sinica, 2019, 56(3): 528-540. |
| [4] |
邵帅, 何红波, 张威, 等. 土壤有机质形成与来源研究进展[J]. 吉林师范大学学报(自然科学版), 2017, 38(1): 126-130. Shao S, He H B, Zhang W, et al. Soil organic matter formation and origin: a review[J]. Journal of Jilin Normal University (Natural Science Edition), 2017, 38(1): 126-130. |
| [5] | Roth V N, Lange M, Simon C, et al. Persistence of dissolved organic matter explained by molecular changes during its passage through soil[J]. Nature Geoscience, 2019, 12(9): 755-761. DOI:10.1038/s41561-019-0417-4 |
| [6] | Lehmann J, Kleber M. The contentious nature of soil organic matter[J]. Nature, 2015, 528(7580): 60-68. DOI:10.1038/nature16069 |
| [7] |
梁超, 朱雪峰. 土壤微生物碳泵储碳机制概论[J]. 中国科学: 地球科学, 2021, 64(5): 680-695. Liang C, Zhu X F. The soil microbial carbon pump as a new concept for terrestrial carbon sequestration[J]. Science China Earth Sciences, 2021, 64(4): 545-558. |
| [8] | Kalbitz K, Schmerwitz J, Schwesig D, et al. Biodegradation of soil-derived dissolved organic matter as related to its properties[J]. Geoderma, 2003, 113(3-4): 273-291. DOI:10.1016/S0016-7061(02)00365-8 |
| [9] |
吴宪, 王蕊, 胡菏, 等. 潮土细菌及真菌群落对化肥减量配施有机肥和秸秆的响应[J]. 环境科学, 2020, 41(10): 4669-4681. Wu X, Wang R, Hu H, et al. Response of bacterial and fungal communities to chemical fertilizer reduction combined with organic fertilizer and straw in fluvo-aquic soil[J]. Environmental Science, 2020, 41(10): 4669-4681. |
| [10] | Deng S H, Zheng X D, Chen X B, et al. Divergent mineralization of hydrophilic and hydrophobic organic substrates and their priming effect in soils depending on their preferential utilization by bacteria and fungi[J]. Biology and Fertility of Soils, 2021, 57(1): 65-76. DOI:10.1007/s00374-020-01503-7 |
| [11] | Xia Y H, Chen X B, Zheng X D, et al. Preferential uptake of hydrophilic and hydrophobic compounds by bacteria and fungi in upland and paddy soils[J]. Soil Biology and Biochemistry, 2020, 148. DOI:10.1016/j.soilbio.2020.107879 |
| [12] | Fontaine S, Mariotti A, Abbadie L. The priming effect of organic matter: a question of microbial competition?[J]. Soil Biology and Biochemistry, 2003, 35(6): 837-843. DOI:10.1016/S0038-0717(03)00123-8 |
| [13] | Reischke S, Rousk J, Bååth E. The effects of glucose loading rates on bacterial and fungal growth in soil[J]. Soil Biology and Biochemistry, 2014, 70: 88-95. DOI:10.1016/j.soilbio.2013.12.011 |
| [14] | Xu W Y, Fu S F, Yang Z M, et al. Improved methane production from corn straw by microaerobic pretreatment with a pure bacteria system[J]. Bioresource Technology, 2018, 259: 18-23. DOI:10.1016/j.biortech.2018.02.046 |
| [15] | Wei L, Ge T D, Zhu Z K, et al. Comparing carbon and nitrogen stocks in paddy and upland soils: accumulation, stabilization mechanisms, and environmental drivers[J]. Geoderma, 2021, 398. DOI:10.1016/j.geoderma.2021.115121 |
| [16] |
王娟. 稻田土壤碳氮转化与微生物群落结构及活性之间的联系机制[D]. 杭州: 浙江大学, 2015. 5-6. Wang J. The relationship of microbial community structure and activity to carbon and nitrogen transformations in paddy soil[D]. Hangzhou: Zhejiang University, 2015. 5-6. |
| [17] | Chen X B, Hu Y J, Xia Y H, et al. Contrasting pathways of carbon sequestration in paddy and upland soils[J]. Global Change Biology, 2021, 27(11): 2478-2490. DOI:10.1111/gcb.15595 |
| [18] | 鲍士旦. 土壤农化分析[M]. ((第三版)). 北京: 中国农业出版社, 2000. |
| [19] | Qiu H S, Ge T D, Liu J Y, et al. Effects of biotic and abiotic factors on soil organic matter mineralization: experiments and structural modeling analysis[J]. European Journal of Soil Biology, 2018, 84: 27-34. DOI:10.1016/j.ejsobi.2017.12.003 |
| [20] | Zhu Z K, Ge T D, Hu Y J, et al. Fate of rice shoot and root residues, rhizodeposits, and microbial assimilated carbon in paddy soil-part 2: turnover and microbial utilization[J]. Plant and Soil, 2017, 416(1-2): 243-257. DOI:10.1007/s11104-017-3210-4 |
| [21] |
徐学池, 苏以荣, 王桂红, 等. 秸秆还田配施氮肥对喀斯特农田微生物群落及有机碳矿化的影响[J]. 环境科学, 2019, 40(6): 2912-2919. Xu X C, Su Y R, Wang G H, et al. Straw returning plus nitrogen fertilizer affects the soil microbial community and organic carbon mineralization in karst farmland[J]. Environmental Science, 2019, 40(6): 2912-2919. |
| [22] | Zang H D, Blagodatskaya E, Wen Y, et al. Carbon sequestration and turnover in soil under the energy crop Miscanthus: repeated 13C natural abundance approach and literature synthesis[J]. GCB Bioenergy, 2018, 10(4): 262-271. DOI:10.1111/gcbb.12485 |
| [23] |
陈智, 于贵瑞. 土壤微生物碳素利用效率研究进展[J]. 生态学报, 2020, 40(3): 756-767. Chen Z, Yu G R. Advances in the soil microbial carbon use efficiency[J]. Acta Ecologica Sinica, 2020, 40(3): 756-767. |
| [24] |
陈昕, 姜成浩, 罗安程. 秸秆微生物降解机理研究[J]. 安徽农业科学, 2013, 41(23): 9728-9731, 9740. Chen X, Jiang C H, Luo A C, et al. Study on the microbial degradation mechanism of straw[J]. Journal of Anhui Agricultural Sciences, 2013, 41(23): 9728-9731, 9740. DOI:10.3969/j.issn.0517-6611.2013.23.070 |
| [25] |
魏圆云, 崔丽娟, 张曼胤, 等. 土壤有机碳矿化激发效应的微生物机制研究进展[J]. 生态学杂志, 2019, 38(4): 1202-1211. Wei Y Y, Cui L J, Zhang M Y, et al. Research advances in microbial mechanisms underlying priming effect of soil organic carbon mineralization[J]. Chinese Journal of Ecology, 2019, 38(4): 1202-1211. |
| [26] | Mazzilli S R, Kemanian A R, Ernst O R, et al. Priming of soil organic carbon decomposition induced by corn compared to soybean crops[J]. Soil Biology and Biochemistry, 2014, 75: 273-281. DOI:10.1016/j.soilbio.2014.04.005 |
| [27] |
仇存璞, 陈晓芬, 刘明, 等. 两种典型水稻土中秸秆碳转化的微生物过程[J]. 中国农业科学, 2019, 52(13): 2268-2279. Qiu C P, Chen X F, Liu M, et al. Microbial transformation process of straw-derived c in two typical paddy soils[J]. Scientia Agricultura Sinica, 2019, 52(13): 2268-2279. DOI:10.3864/j.issn.0578-1752.2019.13.007 |
| [28] | de Vries F T, Hoffland E, van Eekeren N, et al. Fungal/bacterial ratios in grasslands with contrasting nitrogen management[J]. Soil Biology and Biochemistry, 2006, 38(8): 2092-2103. DOI:10.1016/j.soilbio.2006.01.008 |
| [29] | Gunina A, Dippold M, Glaser B, et al. Turnover of microbial groups and cell components in soil: 13C analysis of cellular biomarkers[J]. Biogeosciences, 2017, 14(2): 271-283. DOI:10.5194/bg-14-271-2017 |
| [30] | Zhu Z K, Ge T D, Xiao M L, et al. Belowground carbon allocation and dynamics under rice cultivation depends on soil organic matter content[J]. Plant and Soil, 2017, 410(1-2): 247-258. DOI:10.1007/s11104-016-3005-z |
| [31] | Fabian J, Zlatanovic S, Mutz M, et al. Fungal-bacterial dynamics and their contribution to terrigenous carbon turnover in relation to organic matter quality[J]. The ISME Journal, 2017, 11(2): 415-425. DOI:10.1038/ismej.2016.131 |
| [32] | Xia Y H, Chen X B, Hu Y J, et al. Contrasting contribution of fungal and bacterial residues to organic carbon accumulation in paddy soils across eastern China[J]. Biology and Fertility of Soils, 2019, 55(8): 767-776. DOI:10.1007/s00374-019-01390-7 |
| [33] |
章晓芳, 郑生猛, 夏银行, 等. 红壤丘陵区土壤有机碳组分对土地利用方式的响应特征[J]. 环境科学, 2020, 41(3): 1466-1473. Zhang X F, Zheng S M, Xia Y H, et al. Responses of soil organic carbon fractions to land use types in hilly red soil regions, China[J]. Environmental Science, 2020, 41(3): 1466-1473. |