 2021, Vol. 42
2021, Vol. 42



2. 中国环境科学研究院环境基准与风险评估国家重点实验室, 北京 100012
2. State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012, China
氮是生态系统重要的营养元素, 为生命体生长提供必不可少的物质和能量.由于人类活动的干扰, 全球氮循环失衡, 引发了温室气体排放、水体富营养化和土壤酸化等一系列环境问题[1].反硝化作为氮循环的关键过程, 是一个兼性厌氧的微生物过程, 通过氧化中间体将NO3-或NO2-还原成N2或N2O[2, 3].其中, 亚硝酸盐还原酶(nitrite reductase, Nir)具有催化NO2-还原的能力, 是限制土壤反硝化过程的关键酶和限速步骤[4, 5].Nir有两种结构不同而功能相似的同工酶: nirK基因编码的含铜亚硝酸盐还原酶和nirS基因编码的同源二聚体细胞色素CD1亚硝酸盐还原酶.Mosier等[6]的研究发现nirS型反硝化细菌比nirK型分布更广泛, 其微生物活性、多样性和丰富度也更高; Zhao等[7]的研究发现nirS型反硝化细菌对土壤含氮气体排放的贡献高于nirK型.因此, nirS型反硝化细菌标记物常用来研究土壤反硝化细菌群落组成、多样性和丰度.
反硝化细菌群落与环境条件紧密相关: pH、温度、有机碳和无机氮含量等环境参数均可能影响反硝化菌群的结构与功能[8~10]; 植物根系微环境具有丰富的根系分泌物如有机酸、氨基酸和糖类等可作为微生物的底物或对微生物产生刺激效应, 同时因根系的泌氧作用形成好氧-兼氧-厌氧的氧气浓度梯度也对氮循环相关微生物的耦合产生重要影响[2, 3].不同的生态系统中反硝化过程的主控因素可能不同: 热带森林土壤中, NO3-、微生物量碳和全氮是反硝化细菌群落的主控因子[11]; NH4+、NO2-、总磷、NO3-和溶解氧对景观水体反硝化微生物多样性影响较大[12]; Wu等[3]的研究发现沉水植物的根际有利于洪湖沉积物的氮排放.然而, 目前对于生态系统氮排放及反硝化细菌群落的研究大多数是基于人为影响显著的环境, 而对于人为干扰较低的自然生态系统的研究较少.青藏高原平均海拔在4 000 m以上, 素有“地球第三极”之称, 是地球上分布面积最大、纬度最低、海拔最高且形成时代最新的巨型地貌单元; 是长江和黄河等10余条河流的发源地; 拥有独特的高寒湿地生态系统, 面积超过1×105 km2, 占全国湿地面积的近30%[13, 14].相比于低海拔湿地, 青藏高原的高寒湿地海拔高, 氧气含量低, 人为影响较少, 氮污染负荷较小, 且拥有较为独特的植被[15, 16].对于青藏高原地区氮循环相关微生物的研究, Zhang等[17]的研究发现青藏高原高海拔河流沉积物的氨氧化细菌和古菌丰度随着海拔升高而降低; Zhao等[18]的研究发现青藏高原旱地土壤的厌氧氨氧化菌多样性高于高寒湿地.因此, 进一步探究在青藏高原高寒湿地这一人为影响低的特殊生境下, 反硝化速率以及反硝化细菌群落结构, 将有助于完善人们对自然生态系统氮循环的理解.
本研究通过15N同位素示踪技术测定青藏高原不同季节和不同海拔的高寒湿地植物根际与非根际土壤的反硝化速率及其对总脱氮的贡献; 通过高通量测序技术调查分析nirS型反硝化细菌的群落结构; 通过冗余分析和相关分析探讨反硝化细菌群落组成及反硝化速率与环境因子的关系.
1 材料与方法 1.1 土壤样品采集2017年5月(春季)和8月(夏季)在青藏高原黄河源区不同海拔的高寒湿地: 唐克(Tangke, TK)、久治(Jiuzhi, JZ)、玛多(Maduo, MD)和达日(Dari, DR), 采集植物的根际和非根际土壤, 各采样地位置和概况如图 1和表 1所示.根际土壤样品采样时, 选择3棵相似大小的当地优势植物, 将植株整棵连同土壤挖出(0~20 cm), 用刷子刷下附着于根系表面的土壤作为根际样品, 得到3个根际土壤样品; 采集根际土壤样品的植株附近20 m内, 在无植被覆盖土壤采集同深度土壤作为非根际土壤, 非根际土壤采集3个平行样品.春季与夏季采集根际土壤的植物为相同地点的同一种植物但并非同一植株, 每个季节采集24个土壤样品, 于无菌密封袋中保存, 放置于冰盒带回驻地, 部分在驻地进行泥浆培养实验, 部分转入-80℃保存用于后续土壤理化指标测定和分子生物学分析.

|
图 1 青藏高原黄河源区不同海拔高寒湿地采样点示意 Fig. 1 Location of sampling sites in the alpine wetlands of the Yellow River source region on the Qinghai-Tibet Plateau |
|
|
表 1 春季和夏季青藏高原黄河源区不同海拔高寒湿地采样地概况和优势植物种 Table 1 Environmental conditions and dominant plant species at the sampling sites along an altitudinal gradient and among seasons on the Qinghai-Tibet Plateau |
1.2 土壤理化性质测定
土壤样品剔除石块、植物细根等, 过2 mm筛.新鲜土壤用2 mol·L-1的KCl溶液浸提后, 使用AA3流动分析仪(SEAL, Germany)测定土壤中的铵态氮(NH3-N)、硝态氮(NO3--N)和亚硝态氮(NO2--N)含量.土壤pH采用pH计(SevenGo DuoTM SG23, Mettler Toledo)在1∶2.5质量比的土水悬浊液下测定.室温风干的土壤样品, 研磨后用Vario EL CN元素分析仪(Elementar, Germany)测定土壤有机碳(soil organic carbon, SOC)含量.
1.3 同位素示踪培养经由15N同位素-泥浆培养实验测定反硝化速率, 该实验在驻地进行.新鲜采集的土壤样品以1∶5质量比加入去离子水, 置于12 mL的小瓶(Labco, High Wycombe, Buckinghamshire, UK)中, 用氦气吹扫30 min去除氧气, 用气密盖密封.小瓶在黑暗中预培养24 h去除NO3-和NO2-, 随后分为3组分别添加: ① 15NH4+; ② 15NH4+和14NO3-; ③ 15NO3-, 最终15N含量为100 μmol·L-1.培养8 h后, 向小瓶中注入200 μL的饱和ZnCl2溶液停止微生物活动[19, 20].用MAT253稳定同位素比质谱仪(Thermo Fisher Scientific Inc., MA, U.S.)测定28N2、29N2和30N2的产生量.根据公式(1)计算反硝化速率, 根据公式(2)计算厌氧氨氧化速率, 公式(3)为反硝化速率产生的N2对总N2的贡献[9, 20]:
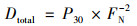
|
(1) |
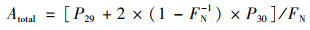
|
(2) |
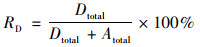
|
(3) |
式中, Dtotal和Atotal分别为反硝化速率和厌氧氨氧化速率[nmol·(g·h)-1], P29和P30分别为29N2和30N2的释放速率[nmol·(g·h)-1], FN为添加15NO3-组中15NO3-占总NO3-的百分比.
1.4 土壤微生物DNA提取和高通量测序准确称量0.5 g新鲜土壤, 根据OMEGA Soil DNA Kit试剂盒提取总DNA, 使用NanoDrop2000分光光度计(Thermo Scientific, USA)检测DNA的浓度和纯度, 保存在-20℃待用.采用nirS型反硝化功能基因引物对: cd3aF(5′-GTSAACGTSAAGGA RACSGG-3′)和R3cd(5′-GASTTCGGRTGSGTC TTGA-3′)进行nirS反硝化功能基因的扩增.扩增体系为20 μL: DNA模板10 ng、5×FastPfu缓冲液4 μL、dNTPs 2 μL(2.5 mmol·L-1)、FastPfu聚合酶0.4 μL、0.8 μL引物(5 μmol·L-1)、剩余体积用双蒸水补足.扩增程序为: 95℃预变性3 min, 27个循环(95℃变性30 s, 55℃退火30 s, 72℃延伸30 s), 最后72℃延伸10 min.纯化后的PCR样本在Illumina MiSeq测序平台测序.测序得到的双端序列数据, 首先根据PE reads之间的overlap关系, 将成对的reads拼接成一条序列, 同时对reads的质量和拼接效果进行质控, 根据序列首尾两段的barcode和引物序列区分样品得到有效序列, 并校正序列方向, 即为优化数据.利用UPARSE按照97%的序列相似度聚类成OTU, 对比GenBank数据库, 得到OTU的物种分类信息.序列及物种分类信息已经提交至国家微生物科学数据中心, accession编号为NMDCN0000JBH~NMDCN0000LT7.
1.5 数据分析采用Excel 2016进行数据整理, OriginPro 2018进行图形绘制与分析.使用R语言的“vegan”包进行基于距离的冗余分析(Distance-based redundancy analysis, db-RDA), 分析环境因子对反硝化微生物群落的影响.利用SPSS 25.0进行Pearson相关分析, 分析反硝化速率和贡献与环境因子的关系.
2 结果与讨论 2.1 土壤理化性质不同季节和海拔青藏高原黄河源区高寒湿地根际与非根际土壤理化性质如表 2所示.高寒湿地土壤pH值在7.30~8.19之间, 呈弱碱性.整体来看, 高寒湿地土壤pH值随海拔升高而升高(春季相关系数0.74, P < 0.05; 夏季相关系数0.93, P < 0.05), 春季土壤样品pH值显著高于同一地点对应的夏季土壤样品pH(P < 0.05), pH随海拔和季节的变化可能与温度有关.Ji等[21]曾报道青藏高原土壤pH与温度呈负相关关系, 温度升高, 土壤有机质将释放更多的H+导致pH降低, 因此, 夏季低海拔地区土壤pH较低.同一地点的非根际土壤pH值和SOC略高于根际土壤, 原因可能是植物的根系分泌物如糖类、氨基酸和其他有机酸的释放导致根际区域SOC含量较高且pH值较低[22, 23].
|
|
表 2 不同季节和海拔青藏高原高寒湿地根际与非根际土壤理化性质1) Table 2 Rhizosphere and bulk soil physicochemical properties of alpine wetlands along an altitudinal and gradient and among seasons on the Qinghai-Tibet Plateau |
整体上看, 土壤中铵态氮、硝态氮和亚硝态氮的含量与海拔没有显著相关关系, 而与采样月温度显著正相关(相关系数0.44, P < 0.05), 这可能是温度更高导致的有机质矿化速率增加造成的.根际土壤的NH3-N含量低于非根际土壤(达日夏季除外), 根际NO3--N含量高于非根际土壤.这与吴讷等[24]的研究结果类似.原因可能是植物对不同氮素形态的需求的差异性造成的[25].且先前的研究表明, 藏区的牧草在生长初期更易吸收NH4+, 后期更易吸收NO3-和NO2-[26], 因此, 整体上春季的NH3-N含量可能低于夏季, 而NO3--N和NO2--N含量高于夏季.例如, 植物根系需要分泌质子交换NH4+, NH4+在生物膜上释放质子以氨的形态通过生物膜; 而NO3-通过生物膜一般采用H+/NO3-共转运的方式[26].
2.2 反硝化速率及反硝化对总N2释放量的贡献高寒湿地土壤反硝化作用产生N2的速率及其对总N2释放量的贡献如图 2所示.春季, 高寒湿地土壤反硝化速率范围为1.44~7.13 nmol·(g·h)-1; 夏季, 高寒湿地土壤反硝化速率范围为0.80~14.98 nmol·(g·h)-1.所有采样点中, 春季和夏季JZ的土壤反硝化速率最高.反硝化同时受到温度以及底物含量的影响[27], 土壤的反硝化速率与采样月温度呈显著正相关(春季相关系数0.67, P < 0.05; 夏季相关系数0.68, P < 0.05).不同季节, TK、JZ、DR地区根际反硝化速率显著高于非根际土壤(P < 0.05), 而MD高寒湿地非根际土壤的反硝化速率比根际更高(P < 0.05), 表明整体看来植物根际微环境可能比非根际更适合反硝化作用.原因可能是植物根系的泌氧造成了根系存在氧气梯度, 氧气的存在不利于反硝化的进行, 但对氨氧化和硝化作用有促进作用[28], 为反硝化微生物提供了更多的底物来源[29]; 同时, 根系分泌物以及有机物残屑也会为反硝化菌提供营养来源[24], 因此整体表现出根际土壤的反硝化速率较高.

|
柱状图上不同小写字母表示反硝化速率在不同采样点之间统计上具有显著性差异(P < 0.05) 图 2 高寒湿地根际和非根际土壤反硝化速率及反硝化对总N2产量的贡献 Fig. 2 N2 production rates through denitrification in the rhizosphere and bulk soil samples of alpine wetlands, and denitrification contributions to N2 production in spring and summer |
春季, 反硝化对总氮释放的贡献率为11.23%~25.39%; 夏季, JZ和MD的非根际土壤反硝化对总氮释放的贡献率分别为69.48%和71.16%, 其余土壤样品反硝化的贡献率为13.95%~37.34%.本研究的反硝化贡献率低于之前的研究, 如印度河河口沉积物反硝化脱氮贡献率为21.9%[30]、水稻土(87.8%~99.8%)[31]和高海拔河流沉积物(27%)[10]等.而姜博等[32]研究了天山天池沉积物中反硝化对脱氮贡献率, 约为17.22%.天山天池和青藏高原地区反硝化脱氮贡献较低的原因可能是这些地区几乎没有大规模的人工氮素输入, 且高海拔、强光照和低温环境可能限制了硝化细菌的生长和活性, 极大地限制了NO3-的生成, 底物不足导致反硝化过程受限.而与此同时, 若该环境下NO2-底物并不受限, 则自养的厌氧氨氧化细菌不会受到明显抑制, 导致反硝化的脱氮效率较低.
2.3 反硝化微生物群落结构本研究每个样品的有效序列范围为13 427~24 843条, 平均长度为371 bp.按照最低序列数抽平后按照97%的相似度进行OTU的聚类.不同季节和海拔高寒湿地根际与非根际土壤nirS型反硝化微生物α多样性指数如表 3所示. Coverage指各样品文库的覆盖率, 本研究中各样本的Coverage范围为0.97~1.00, 表明测序结果接近样本中微生物群落结构的实际情况.ACE和Chao指数反映微生物群落的丰度, 数值越大表明菌群丰度越高.除了春季和夏季的TK和夏季的DR外, 其余采样点的ACE指数、Chao指数和OTU数量均呈现出根际高于非根际的趋势. Shannon和Simpson指数反映微生物群落的多样性, Shannon指数越大, Simpson指数越小表明多样性越高.TK的根际土壤多样性低于非根际, JZ、MD和DR的根际土壤多样性高于非根际土壤.这与已有研究类似: 陈悦等[33]研究了独子山区优势草本植物的根际与非根际土壤微生物功能多样性, 发现植物根际土壤微生物多样性指数大于自身非根际土; 潘福霞等[34]的研究发现人工湿地植物根际的土壤微生物多样性也高于非根际.原因可能是植物根系和植物残体为根际微生物提供了适宜生长的环境和物质来源, 丰富了微生物多样性和丰度.
|
|
表 3 不同季节和海拔青藏高原高寒湿地根际与非根际nirS型反硝化细菌多样性指数1) Table 3 The nirS-type denitrifying bacteria community diversity index for rhizosphere and bulk soil of alpine wetlands along an altitudinal gradient and among seasons on the Qinghai-Tibet Plateau |
本研究发现Proteobacteria是青藏高原高寒湿地土壤中主要的反硝化细菌门, 这与大多数的研究结果一致[35, 36].不同季节和海拔青藏高原高寒湿地根际与非根际nirS型反硝化细菌群落在属水平上的相对丰度如图 3所示.除未分类的反硝化细菌属(uncultured denitrifying bacteria)和相对丰度在所有样本中不足1%的细菌属外, 各土壤样本中含量最高的为一种未分类的变形杆菌(unclassified Proteobacteria), 相对丰度为2.86%~29.41%, 且春季明显高于夏季, 尤其是春季高海拔的DR土壤, 根际与非根际土壤的相对丰度均达到了29%以上, 这说明主导青藏高原高寒湿地反硝化作用的可能有一些独特的未被鉴定的反硝化菌属.相对丰度其次为假单胞菌属(Pseudomonas, 2.45%~26.52%), 从春季到夏季, 假单胞菌属(Pseudomonas)在TK和JZ相对丰度增加, 而在DR和MD降低.杨文焕等[37]研究了包头南海湖高原湖泊沉积物和Wu等[3]研究了洪湖植物根际沉积物, 均发现假单胞菌属(Pseudomonas)是最优势的反硝化菌属.假单胞菌属(Pseudomonas)是目前已知的反硝化细菌中最大的类群之一, 可以去除较高的硝酸盐氮, 因此可能对脱氮有较大作用[38].春季MD的假单胞菌属(Pseudomonas)相对丰度最低, 这可能一定程度上解释春季MD的反硝化速率低于其他地点.贪铜菌属(Cupriavidus, 0%~34.14%)的相对丰度也整体表现为夏季>春季, 根际>非根际, 尤其是夏季DR和TK根际土壤贪铜菌属相对丰度分别达到了34.14%和23.12%.两种硫氧化菌Sulfuritalea(0.04%~27.46%)和Sulfuritortus(0%~23.18%)显示不同的趋势, Sulfuritalea在夏季样本中含量较高; 而Sulfuritortus在春季样本中明显高于夏季, 且非根际土壤中的相对丰度高于根际.且unclassified Nitrosomonadales也呈现夏季(0.01%~10.81%)相对丰度高于春季(0%~6.45%); 副球菌属(Paracoccus)相对丰度春季(0.04%~12.66%)高于夏季(0~2.49%).较为特殊的铁氧化细菌属(Sideroxydans), 在夏季MD非根际土壤中的相对丰度极高, 为51.64%, 而其余土壤样本中相对丰度范围为0~2.45%.

|
图 3 不同季节和海拔青藏高原高寒湿地根际与非根际nirS型反硝化细菌群落在属水平上的相对丰度 Fig. 3 Relative abundance of nirS-type denitrifying bacteria at the genus level for rhizosphere and bulk soil of alpine wetlands along an altitudinal gradient and among seasons on the Qinghai-Tibet Plateau |
采用基于距离的冗余分析(distance-based redundancy analysis, db-RDA)探究环境因子(海拔、温度)和土壤理化性质(pH、SOC、铵态氮、硝态氮和亚硝态氮)对春季和夏季nirS型反硝化微生物群落组成的影响(图 4).结果发现, 春季第一和第二排序轴占总特征值的43.16%, 夏季的第一和第二排序轴占总特征值的42.26%.其中, 春季土壤样本, 海拔显著影响nirS型反硝化微生物群落(P < 0.05); 夏季海拔、pH和NO2-含量对nirS型反硝化微生物群落的影响均显著(P < 0.05); 其余环境因子和土壤理化性质对nirS型反硝化微生物群落的影响不显著.Song等[39]的研究也发现pH与nirS型反硝化微生物群落组成密切相关.NO2-是nirS基因控制过程的底物, nirS型反硝化微生物参与将NO2-还原的过程, 因此其含量通常与nirS反硝化微生物群落相关[40]; 文献[41]也发现NO2-是对nirS型反硝化微生物影响最大的因素.而海拔通过对植被、温度、辐射强度以及大气含氧量等多种条件的影响来影响微生物群落结构[42].

|
图 4 青藏高原高寒湿地土壤nirS型反硝化细菌群落与环境因子相关性的db-RDA分析 Fig. 4 The db-RDA plots of correlation between the nirS-type denitrifying bacterial community and environmental variables of alpine wetland soils on the Qinghai-Tibet Plateau |
反硝化速率和贡献、菌群的多样性和丰富度指数、以及主要菌属与环境因子的Pearson相关性如表 4所示.unclassified Nitrosomonadales的相对丰度与采样月温度呈显著正相关(P < 0.05), 与土壤pH显著负相关(P < 0.05).Paracoccus的相对丰度与采样月温度呈显著负相关(P < 0.05).反硝化速率和Shannon指数与土壤pH显著负相关(P < 0.05), Simpson指数与土壤pH显著正相关(P < 0.05). Šimek等[43]的研究也发现反硝化速率与pH呈现负相关趋势.这说明, 适当提高土壤的pH有利于降低反硝化细菌的多样性和丰度, 以及减缓反硝化的气体释放, 青藏高原高寒湿地的土壤pH偏碱性, 反硝化速率较低, 对总N2释放的贡献较小.而SOC、铵态氮、硝态氮和亚硝态氮含量对反硝化速率和微生物多样性和丰富度以及主要菌属的影响均不显著, 这与Jiao等[44]的研究结果类似.汪银龙等[8]的研究表明汾河下游水体的nirS反硝化细菌优势属主要受到温度、pH和溶解氧这3个因素影响, 而无机氮几乎没有影响.
|
|
表 4 青藏高原高寒湿地土壤反硝化速率、nirS型反硝化细菌群落多样性及相对丰度前十的菌属与环境因子的Pearson相关分析1) Table 4 Pearson correlation coefficients between environmental variables and denitrification rates and nirS-type denitrifying bacterial community diversity indexes of alpine wetland soils on the Qinghai-Tibet Plateau |
3 结论
(1) 青藏高原高寒湿地土壤反硝化速率范围为0.80~14.98 nmol·(g·h)-1, 整体上呈现出根际土壤的反硝化速率高于非根际的趋势(P < 0.05); 反硝化对总N2释放的贡献率为11.23%~71.16%, 贡献率低于其他生态系统.
(2) 高通量测序结果显示, 各土壤样本中相对丰度较高的为一种未分类的变形杆菌(unclassified Proteobacteria, 2.86%~29.41%), 其次为假单胞菌属(Pseudomonas, 2.45%~26.52%)和贪铜菌属(Cupriavidus, 0~34.14%).说明主导青藏高原高寒湿地这一特殊生境反硝化作用的可能包括独特的未被鉴定的反硝化菌属.
(3) 高寒湿地的反硝化细菌群落结构主要受到海拔、pH和亚硝态氮的影响(P < 0.05); 主要反硝化菌群的相对丰度受到温度和pH的影响; SOC和无机氮对nirS反硝化细菌的影响均不显著.
| [1] |
何芳, 张丽梅, 申聪聪, 等. 青藏高原林地土壤的氮转化特征及其影响因素分析: 以祁连山和藏东南地区为例[J]. 环境科学, 2021, 42(5): 2449-2456. He F, Zhang L M, Shen C C, et al. Analysis of nitrogen transformation characteristics and influencing factors of forestland soil in the Qinghai-Tibet Plateau: a case study of the Qilian Mountains and Southeast Tibet[J]. Environmental Science, 2021, 42(5): 2449-2456. DOI:10.3969/j.issn.1000-6923.2021.05.053 |
| [2] | Vilas M P, Marti C L, Adams M P, et al. Invasive macrophytes control the spatial and temporal patterns of temperature and dissolved oxygen in a shallow lake: a proposed feedback mechanism of macrophyte loss[J]. Frontiers in Plant Science, 2017, 8. DOI:10.3389/fpls.2017.02097 |
| [3] | Wu H P, Hao B B, Cai Y P, et al. Effects of submerged vegetation on sediment nitrogen-cycling bacterial communities in Honghu Lake(China)[J]. Science of the Total Environment, 2021, 755. DOI:10.1016/j.scitotenv.2020.142541 |
| [4] | Sun Y L, Li A, Zhang X N, et al. Regulation of dissolved oxygen from accumulated nitrite during the heterotrophic nitrification and aerobic denitrification of Pseudomonas stutzeri T13[J]. Applied Microbiology and Biotechnology, 2015, 99(7): 3243-3248. DOI:10.1007/s00253-014-6221-6 |
| [5] | Pajares S, Bohannan B J M. Ecology of nitrogen fixing, nitrifying, and denitrifying microorganisms in tropical forest soils[J]. Frontiers in Microbiology, 2016, 7. DOI:10.3389/fmicb.2016.01045 |
| [6] | Mosier A C, Francis C A. Denitrifier abundance and activity across the San Francisco Bay estuary[J]. Environmental Microbiology Reports, 2010, 2(5): 667-676. DOI:10.1111/j.1758-2229.2010.00156.x |
| [7] | Zhao S Y, Zhou J M, Yuan D D, et al. NirS-type N2O-producers and nosZ Ⅱ-type N2O-reducers determine the N2O emission potential in farmland rhizosphere soils[J]. Journal of Soils and Sediments, 2020, 20(1): 461-471. DOI:10.1007/s11368-019-02395-3 |
| [8] |
汪银龙, 冯民权, 董向前. 汾河下游水体nirS型反硝化细菌群落组成与无机氮关系[J]. 环境科学, 2019, 40(8): 3596-3603. Wang Y L, Feng M Q, Dong X Q. Community composition of nirS-type denitrifying bacteria in the waters of the lower reaches of the Fenhe River and its relationship with inorganic nitrogen[J]. Environmental Science, 2019, 40(8): 3596-3603. |
| [9] |
陈志浩, 覃云斌, 丁帮璟, 等. 宛山荡农田土壤氮迁移过程反硝化与厌氧氨氧化[J]. 环境科学, 2020, 41(1): 412-419. Chen Z H, Qin Y B, Ding B J, et al. Denitrification and anaerobic ammonium oxidation in soil nitrogen migration process in a farmland of Wanshandang Lake[J]. Environmental Science, 2020, 41(1): 412-419. |
| [10] | Zhang S B, Qin W, Bai Y B, et al. Linkages between anammox and denitrifying bacterial communities and nitrogen loss rates in high-elevation rivers[J]. Limnology and Oceanography, 2021, 66(3): 765-778. DOI:10.1002/lno.11641 |
| [11] |
曹乾斌, 王邵军, 陈闽昆, 等. 不同恢复阶段热带森林土壤nirS型反硝化微生物群落结构及多样性特征[J]. 生态学报, 2021, 41(2): 626-636. Cao Q B, Wang S J, Chen M K, et al. The structure and diversity of nirS-denitrifying microbial community across three restoration stages of Xishuangbanna tropical forests[J]. Acta Ecologica Sinica, 2021, 41(2): 626-636. |
| [12] |
张艺冉, 李再兴, 孙悦, 等. 石家庄市春季景观水体nirS型反硝化细菌群落特征分析[J]. 环境科学, 2019, 40(7): 3295-3303. Zhang Y R, Li Z X, Sun Y, et al. Community structure characteristics of nirS denitrifying bacteria of spring typical parkland waterbodies in Shijiazhuang City[J]. Environmental Science, 2019, 40(7): 3295-3303. |
| [13] | Bai J H, Ouyang H, Xiao R, et al. Spatial variability of soil carbon, nitrogen, and phosphorus content and storage in an alpine wetland in the Qinghai-Tibet Plateau, China[J]. Australian Journal of Soil Research, 2010, 48(8): 730-736. DOI:10.1071/SR09171 |
| [14] |
赵志龙, 张镱锂, 刘林山, 等. 青藏高原湿地研究进展[J]. 地理科学进展, 2014, 33(9): 1218-1230. Zhao Z L, Zhang Y L, Liu L S, et al. Advances in research on wetlands of the Tibetan Plateau[J]. Progress in Geography, 2014, 33(9): 1218-1230. |
| [15] | Xia X H, Li S L, Wang F, et al. Triple oxygen isotopic evidence for atmospheric nitrate and its application in source identification for river systems in the Qinghai-Tibetan Plateau[J]. Science of the Total Environment, 2019, 688: 270-280. DOI:10.1016/j.scitotenv.2019.06.204 |
| [16] | Wang S Y, Liu W Y, Zhao S Y, et al. Denitrification is the main microbial N loss pathway on the Qinghai-Tibet Plateau above an elevation of 5000 m[J]. Science of the Total Environment, 2019, 696. DOI:10.1016/j.scitotenv.2019.133852 |
| [17] | Zhang S B, Qin W, Xia X H, et al. Ammonia oxidizers in river sediments of the Qinghai-Tibet Plateau and their adaptations to high-elevation conditions[J]. Water Research, 2020, 173. DOI:10.1016/j.watres.2020.115589 |
| [18] | Zhao S Y, Zhuang L J, Wang C, et al. High-throughput analysis of anammox bacteria in wetland and dryland soils along the altitudinal gradient in Qinghai-Tibet Plateau[J]. Microbiologyopen, 2018, 7(2). DOI:10.1002/mbo3.556 |
| [19] | Hou L J, Zheng Y L, Liu M, et al. Anaerobic ammonium oxidation(anammox)bacterial diversity, abundance, and activity in marsh sediments of the Yangtze Estuary[J]. Journal of Geophysical Research: Biogeosciences, 2013, 118(3): 1237-1246. DOI:10.1002/jgrg.20108 |
| [20] | Xue D M, Yu H, Fang Y T, et al. 15 N-tracer approach to assess nitrogen cycling processes: nitrate reduction, anammox and denitrification in different pH cropland soils[J]. CATENA, 2020, 193. DOI:10.1016/j.catena.2020.104611 |
| [21] | Ji C J, Yang Y H, Han W X, et al. Climatic and edaphic controls on soil pH in alpine grasslands on the Tibetan Plateau, China: a quantitative analysis[J]. Pedosphere, 2014, 24(1): 39-44. DOI:10.1016/S1002-0160(13)60078-8 |
| [22] | Finn D, Page K, Catton K, et al. Ecological stoichiometry controls the transformation and retention of plant-derived organic matter to humus in response to nitrogen fertilisation[J]. Soil Biology and Biochemistry, 2016, 99: 117-127. DOI:10.1016/j.soilbio.2016.05.006 |
| [23] | Du L S, Zhu Z K, Qi Y T, et al. Effects of different stoichiometric ratios on mineralisation of root exudates and its priming effect in paddy soil[J]. Science of the Total Environment, 2020, 743. DOI:10.1016/j.scitotenv.2020.140808 |
| [24] |
吴讷, 邵嘉薇, 盛荣, 等. 水稻分蘖期和孕穗期根际反硝化菌群落结构及功能变化[J]. 应用生态学报, 2019, 30(4): 1344-1350. Wu N, Shao J W, Sheng R, et al. Variation of community structure and function of rhizosnheric denitrifiers at tillering and booting stages of rice[J]. Chinese Journal of Applied Ecology, 2019, 30(4): 1344-1350. |
| [25] | Jampeetong A, Konnerup D, Piwpuan N, et al. Interactive effects of nitrogen form and pH on growth, morphology, N uptake and mineral contents of Coix lacryma-jobi L.[J]. Aquatic Botany, 2013, 111: 144-149. DOI:10.1016/j.aquabot.2013.06.002 |
| [26] | Cui J H, Yu C Q, Qiao N, et al. Plant preference for NH4+ versus NO3- at different growth stages in an alpine agroecosystem[J]. Field Crops Research, 2017, 201: 192-199. DOI:10.1016/j.fcr.2016.11.009 |
| [27] |
赵锋, 许海, 詹旭, 等. 太湖春夏两季反硝化与厌氧氨氧化速率的空间差异及其影响因素[J]. 环境科学, 2021, 42(5): 2296-2302. Zhao F, Xu H, Zhan X, et al. Spatial differences and influencing factors of denitrification and anammox rates in spring and summer in Lake Taihu[J]. Environmental Science, 2021, 42(5): 2296-2302. |
| [28] | Zheng Y L, Hou L J, Liu M, et al. Community composition and activity of anaerobic ammonium oxidation bacteria in the rhizosphere of salt-marsh grass Spartina alterniflora[J]. Applied Microbiology and Biotechnology, 2016, 100(18): 8203-8212. DOI:10.1007/s00253-016-7625-2 |
| [29] | Zhang Z X, Zhang W Z, Yang H C, et al. Elevated N2O emission by the rice roots: based on the abundances of narG and bacterial amoA genes[J]. Environmental Science and Pollution Research, 2017, 24(2): 2116-2125. DOI:10.1007/s11356-016-7993-2 |
| [30] | Fozia, Zheng Y L, Hou L J, et al. Anaerobic ammonium oxidation(anammox)bacterial diversity, abundance, and activity in sediments of the Indus Estuary[J]. Estuarine, Coastal and Shelf Science, 2020, 243. DOI:10.1016/j.ecss.2020.106925 |
| [31] | Bai R, Xi D, He J Z, et al. Activity, abundance and community structure of anammox bacteria along depth profiles in three different paddy soils[J]. Soil Biology and Biochemistry, 2015, 91: 212-221. DOI:10.1016/j.soilbio.2015.08.040 |
| [32] |
姜博, 祝贵兵, 周磊榴, 等. 低温高海拔湖泊岸边带厌氧氨氧化菌的存在、生物多样性及活性——以天山天池为例[J]. 环境科学学报, 2015, 35(7): 2045-2051. Jiang B, Zhu G B, Zhou L L, et al. The activity and diversity analysis of anaerobic ammonium oxidation bacteria in the littoral sediments of the Heavenly Lake[J]. Acta Scientiae Circumstantiae, 2015, 35(7): 2045-2051. |
| [33] |
陈悦, 吕光辉, 李岩. 独山子区优势草本植物根际与非根际土壤微生物功能多样性[J]. 生态学报, 2018, 38(9): 3110-3117. Chen Y, Lv G H, Li Y. Soil microbial functional diversity of rhizosphere and non-rhizosphere of three dominant herbaceous plants in the Dushanzi District[J]. Acta Ecologica Sinica, 2018, 38(9): 3110-3117. |
| [34] |
潘福霞, 来晓双, 李欣, 等. 不同湿地植物脱氮效果与根际土壤微生物群落功能多样性特征分析[J]. 环境科学研究, 2020, 33(6): 1497-1503. Pan F X, Lai X S, Li X, et al. Nitrogen removal efficiencies and rhizosphere soil microbial community functional diversities of different plants in constructed wetlands[J]. Research of Environmental Sciences, 2020, 33(6): 1497-1503. |
| [35] |
胡晓婧, 刘俊杰, 于镇华, 等. 东北黑土nirS型反硝化细菌群落和网络结构对长期施用化肥的响应[J]. 植物营养与肥料学报, 2020, 26(1): 1-9. Hu X J, Liu J J, Yu Z H, et al. Response of nirS-type denitrifier community and network structures to long-term application of chemical fertilizers in a black soil of Northeast China[J]. Journal of Plant Nutrition and Fertilizers, 2020, 26(1): 1-9. |
| [36] | Hou S P, Ai C, Zhou W, et al. Structure and assembly cues for rhizospheric nirK-and nirS-type denitrifier communities in long-term fertilized soils[J]. Soil Biology and Biochemistry, 2018, 119: 32-40. DOI:10.1016/j.soilbio.2018.01.007 |
| [37] |
杨文焕, 石大钧, 张元, 等. 高原湖泊沉积物中反硝化微生物的群落特征——以包头南海湖为例[J]. 中国环境科学, 2020, 40(1): 431-438. Yang W H, Shi D J, Zhang Y, et al. Community characteristics of denitrifying microorganisms in plateau lake sediments-taking Nanhaihu lake as example[J]. China Environmental Science, 2020, 40(1): 431-438. DOI:10.3969/j.issn.1000-6923.2020.01.048 |
| [38] |
王弘宇, 马放, 杨开, 等. 两株异养硝化细菌的氨氮去除特性[J]. 中国环境科学, 2009, 29(1): 47-52. Wang H Y, Ma F, Yang K, et al. Ammonia removal by two strains of heterotrophic nitrifying bacteria[J]. China Environmental Science, 2009, 29(1): 47-52. DOI:10.3321/j.issn:1000-6923.2009.01.010 |
| [39] | Song H, Che Z, Jin W J, et al. Changes in denitrifier communities and denitrification rates in an acidifying soil induced by excessive N fertilization[J]. Archives of Agronomy and Soil Science, 2020, 66(9): 1203-1217. DOI:10.1080/03650340.2019.1661382 |
| [40] | Yin X J, Lu J, Wang Y C, et al. The abundance of nirS-type denitrifiers and anammox bacteria in rhizospheres was affected by the organic acids secreted from roots of submerged macrophytes[J]. Chemosphere, 2020, 240. DOI:10.1016/j.chemosphere.2019.124903 |
| [41] |
毛铁墙, 董宏坡, 陈法锦, 等. 湛江湾沉积物中反硝化和厌氧氨氧化细菌的丰度、多样性及分布特征[J]. 海洋与湖沼, 2020, 51(1): 59-74. Mao T Q, Dong H P, Chen F J, et al. Abundance, diversity, and distribution of denitrifier and anammox bacteria in Zhanjiang Bay sediments[J]. Oceanologia et Limnologia Sinica, 2020, 51(1): 59-74. |
| [42] |
王晓彤, 靳振江, 周军波, 等. 龙脊稻作梯田土壤细菌群落结构和功能类群及影响因子分析[J]. 农业资源与环境学报, 2021, 38(3): 365-376. Wang X T, Jin Z J, Zhou J B, et al. Community structure and functional groups of soil bacteria and their influencing factors in Longji rice terraces[J]. Journal of Agricultural Resources and Environment, 2021, 38(3): 365-376. |
| [43] | Šimek M, Jíšová L, Hopkins D W. What is the so-called optimum pH for denitrification in soil?[J]. Soil Biology and Biochemistry, 2002, 34(9): 1227-1234. DOI:10.1016/S0038-0717(02)00059-7 |
| [44] | Jiao L J, Wu J P, He X, et al. Significant microbial nitrogen loss from denitrification and anammox in the land-sea interface of low permeable sediments[J]. International Biodeterioration & Biodegradation, 2018, 135: 80-89. |